

Chinese Species of the Genus Pseudanaphes Noyes & Valentine (Hymenoptera: Mymaridae) with Description of a New Species †

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens Collection
2.2. Taxonomic Studies
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Sequence Analysis
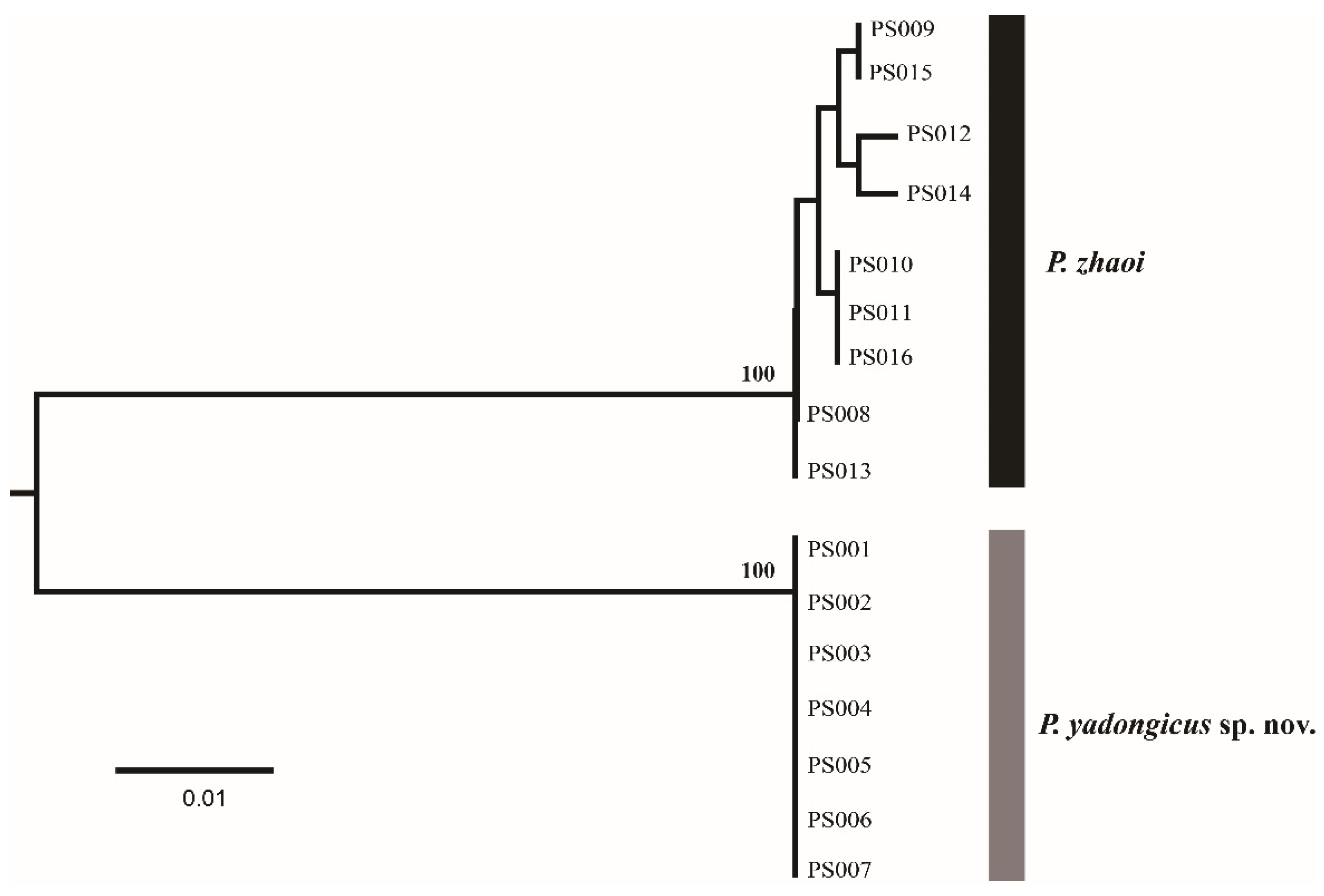
3. Results
3.1. Pseudanaphes Noyes & Valentine, 1989
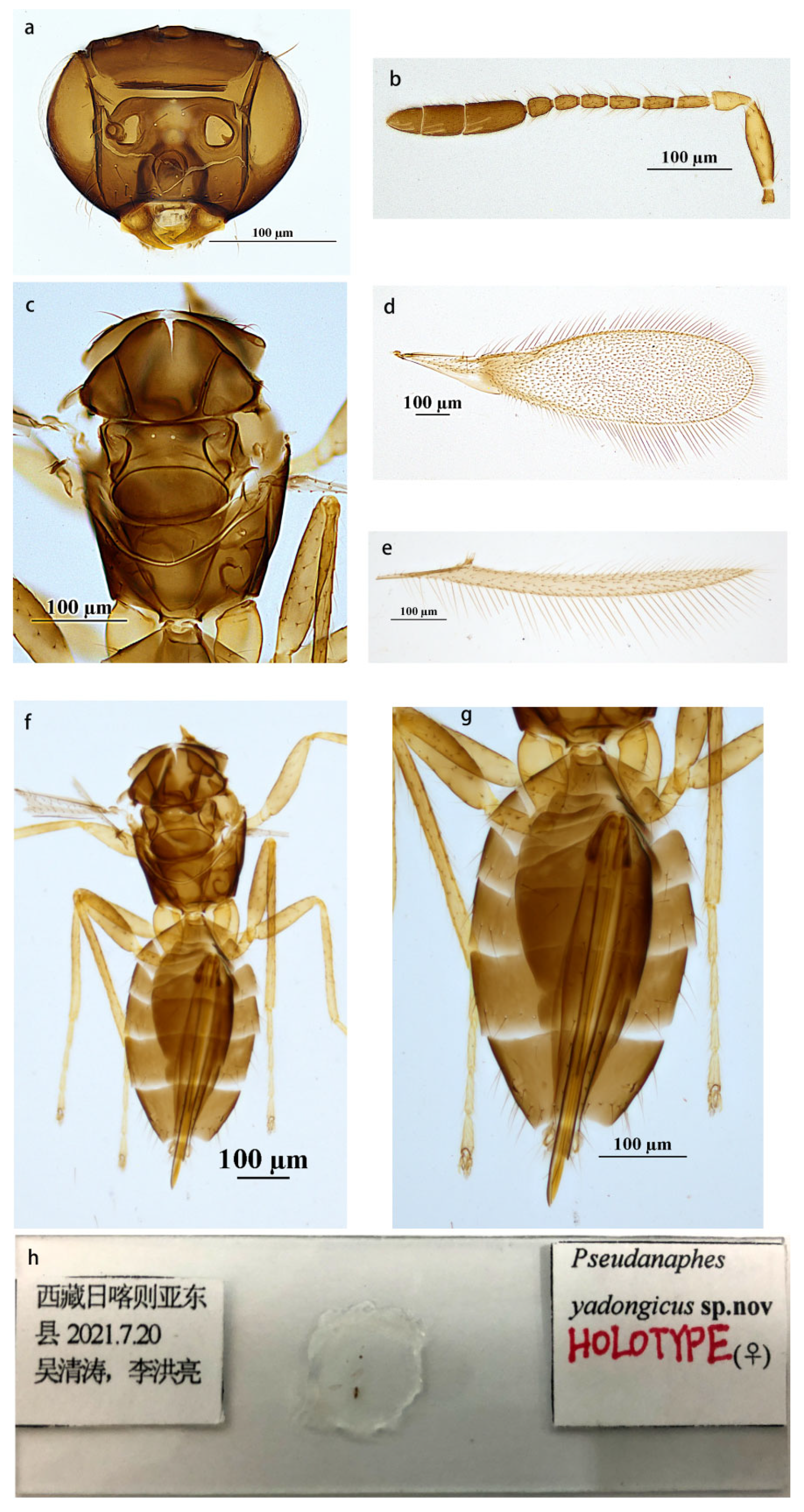
3.1.1. Pseudanaphes yadongicus Aishan & Cao, sp. nov.
3.1.2. Pseudanaphes zhaoi Lin, 1997
3.2. Key to Species of Pseudanaphes (Female)
- urn:lsid:zoobank.org:act:7B041410-9F8A-4AB2-A693-BB83DA19F0FF
- 1.
- The first claval segment longer than the other two claval segments…2
- -
- The first claval segment shorter than at least one of the other two claval segments…3
- 2.
- Ovipositor notably exserted beyond apex of metasoma; clava at most 4.85× as long as wide, apical claval segment the shortest…P. yadongicus Aishan & Cao, sp. nov.
- -
- Ovipositor not exserted beyond apex of metasoma; clava 5.91× as long as wide, second claval segment the shortest…P. lincolni (Girault)
- 3.
- Metasoma with anterior scutellum with reticulate sculpture near posterior margin…P. hirtus Noyes & Valentine
- -
- Metasoma smooth, without reticulate sculpture…4
- 4.
- Mesothorax golden; first segment of clava the shortest, clava 3.42× as long as wide…P. particoxae (Girault)
- -
- Mesothorax brown to dark brown; first segment of clava not the shortest, clava at least 3.9× as long as wide…5
- 5.
- F5 and F4 subequal in length, F4 without longitudinal sensilla; anterior scutellum without setae; fore wing 3.33× as long as wide…P. sikkimianus Rehmat & Anis
- -
- F5 distinctly shorter than F4, F4 with one longitudinal sensillum; anterior scutellum with one pair of setae; fore wing 2.85× as long as wide…P. zhaoi Lin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noyes, J.S.; Valentine, E.W. Mymaridae (Insecta: Hymenoptera)-introduction and review of genera. Fauna N. Z. 1989, 17, 47–48. [Google Scholar]
- Lin, N.Q. A new species of Pseudanaphes (Hymenoptera: Mymaridae) from China. Wuyi Sci. J. 1997, 13, 98–100. [Google Scholar]
- Girault, A.A. Australian Hymenoptera Chalcidoidea—II. Supplement. Mem. Qld. Mus. 1913, 13, 107–129. [Google Scholar]
- Girault, A.A. New Trichogrammatidae and Mymaridae from Australia (Hym). Rev. Entomol. 1938 9, 382–396.
- New, T.R. The relationships of the genus Australomymar (Hymenoptera: Mymaridea). Entomophaga 1974, 19, 229–235. [Google Scholar] [CrossRef]
- Lin, N.Q.; Huber, J.T.; LaSalle, J. The Australian genera of Mymaridae (Hymenoptera: Chalcidoidea). Zootaxa 2007, 1596, 1–111. [Google Scholar] [CrossRef]
- Rehmat, T.; Anis, S.B. First record of Pseudanaphes (Hymenoptera: Mymaridae) from India, with description of a new species. Biosystematica 2011, 5, 14–16. [Google Scholar]
- Noyes, J. Universal Chalcidoidea Database. 2019. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 11 June 2022).
- Triapitsyn, S.V.; Rugman-Jones, P.F.; Tretiakov, P.S.; Daane, K.M.; Wilson, H. Reassessment of molecular and morphological variation within the Anagrus atomus species complex (Hymenoptera: Mymaridae): Egg parasitoids of leafhoppers (Hemiptera: Cicadellidae) in Europe and North America. J. Nat. Hist. 2020, 54, 1735–1758. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. NucleicAcids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuber, J.T. Systematics, biology, and hosts of the Mymaridae and Mymarommatidae (Insecta: Hymenoptera): 1758–1984. Entomography 1986, 4, 185–243. [Google Scholar]
- Zhou, Q.S.; Xi, Y.Q.; Fang, Y.; Zhang, X.; Li, X.J.; Liu, C.L.; Niu, Z.Q.; Zhu, C.D.; Qiao, G.X.; Zhang, Y.Z. Application of DNA barcoding to the identification of hymenoptera parasitoids from the soybean aphid (Aphis glycines) in china. Insect Sci. 2014, 21, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.T.; Read, J.D.; Triapitsyn, S.V. Illustrated key to genera and catalogue of Mymaridae (Hymenoptera) in America north of Mexico. Zootaxa 2020, 4773, 4773. [Google Scholar] [CrossRef] [PubMed]
- Triapitsyn, S.V. Revised Taxonomy of the Common Northern European Fairyfly Polynema (Doriclytus) Atratum Haliday, 1833 (Hymenoptera: Mymaridae), with Fourteen New Synonymies. Ann. Zool. Fenn. 2021, 58, 87–107. [Google Scholar] [CrossRef]
- Triapitsyn, S.V.; Koponen, M.; Vikberg, V.; Várkonyi, G. Taxonomy, annotated new records and a checklist of Mymaridae (Hymenoptera) of Finland, with description of a new species of Eustochus. Acta Entomol. Musei Natl. Pragae 2020, 60, 565–589. [Google Scholar] [CrossRef]
- Triapitsyn, S.V.; Rugman-Jones, P.F.; Jeong, G.; Morse, J.G.; Stouthamer, R. Morphological and molecular differentiation of the Anagrus epos species complex (Hymenoptera: Mymaridae), egg parasitoids of leafhoppers Hemiptera: Cicadellidae) in North America. Zootaxa 2010, 2428, 1. [Google Scholar] [CrossRef]
- Triapitsyn, S.V.; Rugman-Jones, P.F.; Tretiakov, P.S.; Shih, H.T.; Huang, S.H. New synonymies in the Anagrus incarnatus haliday ‘species complex’ (Hymenoptera: Mymaridae) including a common parasitoid of economically important planthopper (Hemiptera: Delphacidae) pests of rice in Asia. J. Nat. Hist. 2018, 52, 2795–2822. [Google Scholar] [CrossRef]
- Munro, J.B.; Heraty, J.M.; Burks, R.A.; Hawks, D.; Mottern, J.; Cruaud, A.; Rasplus, J.-Y.; Jansta, P. A molecular phylogeny of the Chalcidoidea (Hymenoptera). PLoS ONE 2011, 6, e27023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noort, S.V.; Triapitsyn, S.V. Revision of the Polynema dikobraz species-group with description of a remarkable new species from South Africa (Hymenoptera, Chalcidoidea, Mymaridae). ZooKeys 2018, 783, 67–84. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species | Sex | Primer Pair | Accession Number |
|---|---|---|---|---|
| PS001 PS002 PS003 PS004 PS005 PS006 PS007 PS008 PS009 PS010 PS011 PS012 PS013 PS014 PS015 PS016 | P. yadongicus P. yadongicus P. yadongicus P. yadongicus P. yadongicus P. yadongicus P. yadongicus P. zhaoi P. zhaoi P. zhaoi P. zhaoi P. zhaoi P. zhaoi P. zhaoi P. zhaoi P. zhaoi | female female female female female female female female female female female female female female female female | Dicopus-COI-F and Gonat-COI-R Alaptus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Alaptus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Dicopus-COI-F and Gonat-COI-R Alaptus-COI-F and Gonat-COI-R Alaptus-COI-F and Gonat-COI-R Alaptus-COI-F and Gonat-COI-R | ON834699 ON834700 ON834701 ON834702 ON834703 ON834704 ON834705 ON834706 ON834707 ON834708 ON834709 ON834710 ON834711 ON834712 ON834713 ON834714 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aishan, Z.; Cao, H.-X.; Hu, H.-Y.; Zhu, C.-D. Chinese Species of the Genus Pseudanaphes Noyes & Valentine (Hymenoptera: Mymaridae) with Description of a New Species. Insects 2023, 14, 39. https://doi.org/10.3390/insects14010039
Aishan Z, Cao H-X, Hu H-Y, Zhu C-D. Chinese Species of the Genus Pseudanaphes Noyes & Valentine (Hymenoptera: Mymaridae) with Description of a New Species. Insects. 2023; 14(1):39. https://doi.org/10.3390/insects14010039
Chicago/Turabian StyleAishan, Zhulidezi, Huan-Xi Cao, Hong-Ying Hu, and Chao-Dong Zhu. 2023. "Chinese Species of the Genus Pseudanaphes Noyes & Valentine (Hymenoptera: Mymaridae) with Description of a New Species" Insects 14, no. 1: 39. https://doi.org/10.3390/insects14010039
APA StyleAishan, Z., Cao, H. -X., Hu, H. -Y., & Zhu, C. -D. (2023). Chinese Species of the Genus Pseudanaphes Noyes & Valentine (Hymenoptera: Mymaridae) with Description of a New Species. Insects, 14(1), 39. https://doi.org/10.3390/insects14010039