


Fitness Cost of the Field-Evolved Resistance to Sulfoxaflor and Multi-Insecticide Resistance of the Wheat Aphid Sitobion miscanthi (Takahashi)
 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Insecticides
2.2. Insecticide Bioassays
2.3. Fitness Comparison
2.4. Data Analysis
3. Results
3.1. Field-Evolved Resistance of SulR and SS S. miscanthi Populations to Insecticides
3.2. Developmental Duration and Fecundity of SulR and SS S. miscanthi Populations
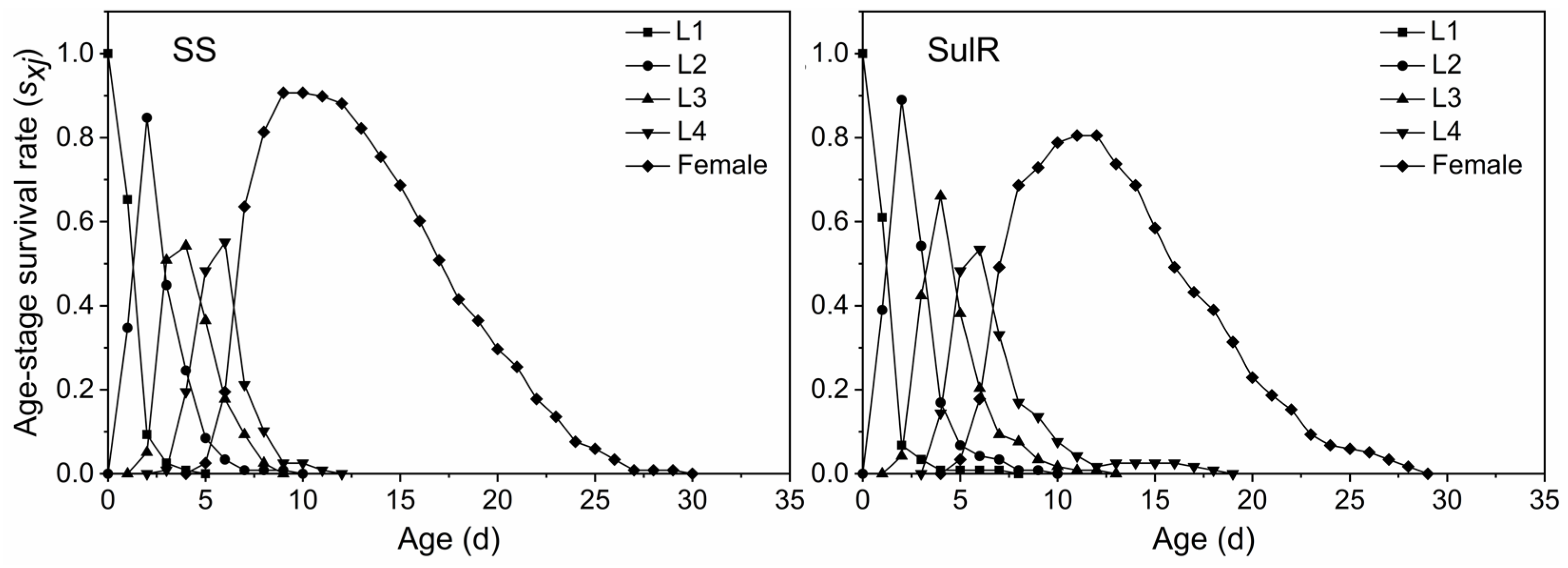
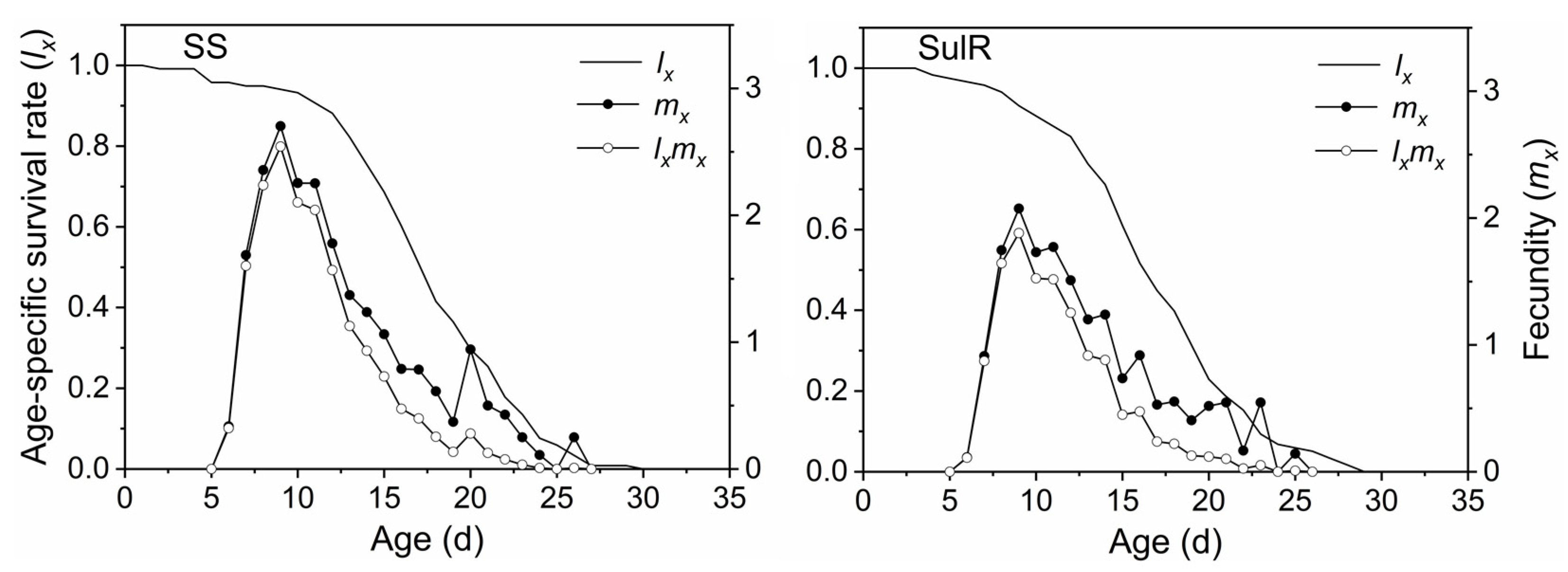
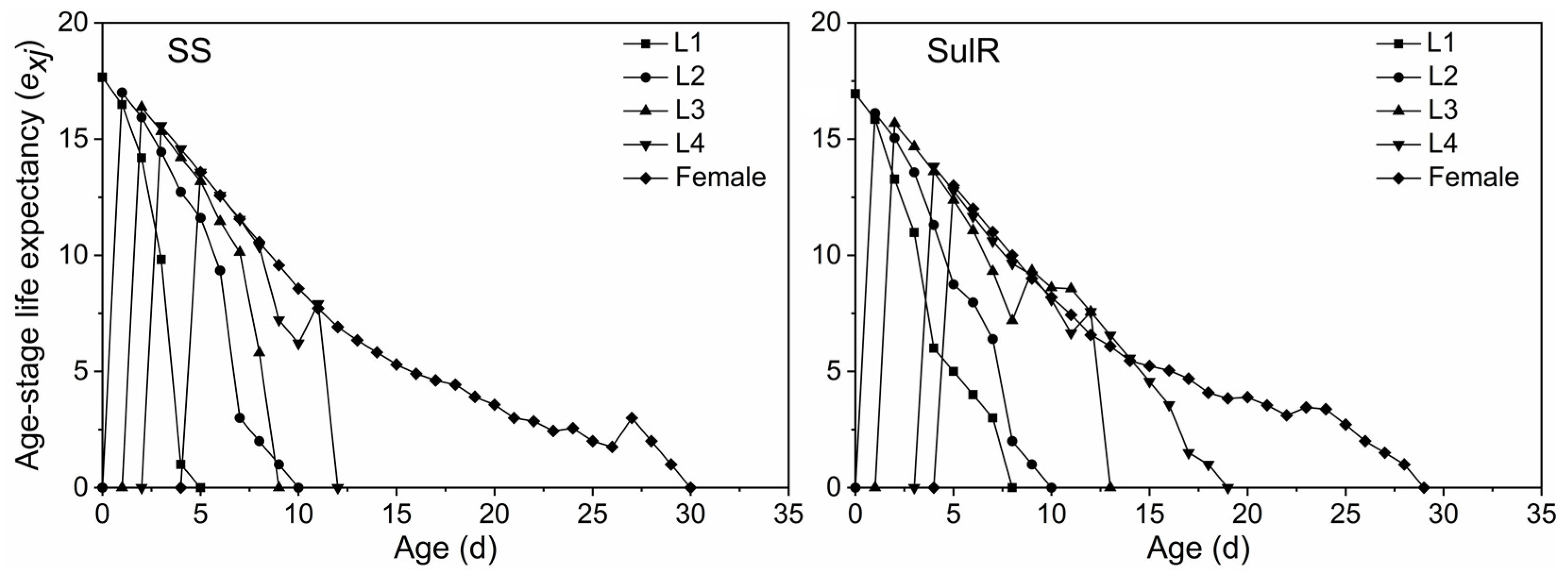
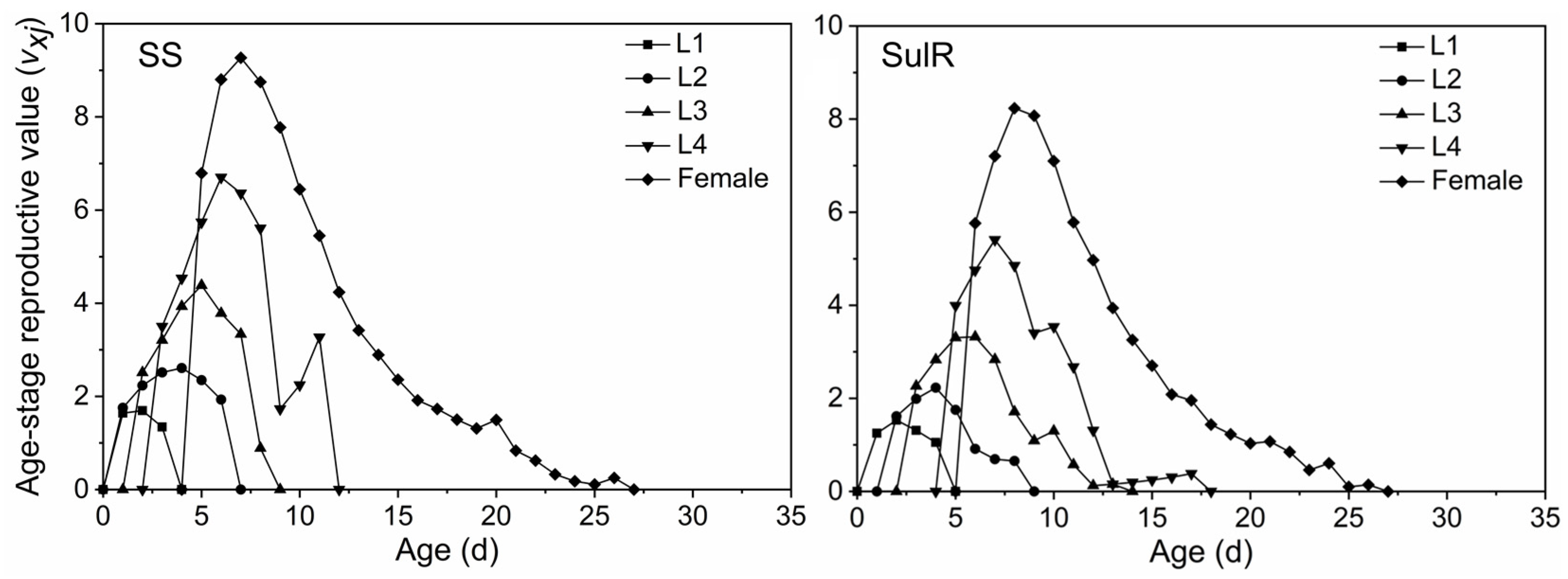
3.3. Comparison of sxj, lxmx, exj, and vxj between the SulR and SS S. miscanthi Populations
3.4. Population Life Table Parameters of the SulR and SS S. miscanthi Populations
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Babcock, J.M.; Gerwick, C.B.; Huang, J.X.; Loso, M.R.; Nakamura, G.; Nolting, S.P.; Rogers, R.B.; Sparks, T.C.; Thomas, J.; Watson, G.B.; et al. Biological characterization of sulfoxaflor, a novel insecticide. Pest Manag. Sci. 2011, 67, 328–334. [Google Scholar] [CrossRef]
- Sparks, T.C.; Watson, G.B.; Loso, M.R.; Geng, C.; Babcock, J.M.; Thomas, J.D. Sulfoxaflor and the sulfoximine insecticides: Chemistry, mode of action and basis for efficacy on resistant insects. Pestic. Biochem. Physiol. 2013, 107, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Watson, G.B.; Loso, M.R.; Babcock, J.M.; Hasler, J.M.; Letherer, T.J.; Young, C.D.; Zhu, Y.M.; Casida, J.E.; Sparks, T.C. Novel nicotinic action of the sulfoximine insecticide sulfoxaflor. Insect Biochem. Mol. Biol. 2011, 41, 432–439. [Google Scholar] [CrossRef]
- Zhu, Y.; Loso, M.R.; Watson, G.B.; Sparks, T.C.; Rogers, R.B.; Huang, J.X.; Gerwick, B.C.; Babcock, J.M.; Kelley, D.; Hegde, V.B.; et al. Discovery and characterization of sulfoxaflor, a novel insecticide targeting sap-feeding pests. J. Agric. Food Chem. 2011, 59, 2950–2957. [Google Scholar] [CrossRef]
- Liao, X.; Mao, K.; Ali, E.; Zhang, X.; Wan, H.; Li, J. Temporal variability and resistance correlation of sulfoxaflor susceptibility among Chinese populations of the brown planthopper Nilaparvata lugens (Stål). Crop Prot. 2017, 102, 141–146. [Google Scholar] [CrossRef]
- Ma, K.; Tang, Q.; Xia, J.; Lv, N.; Gao, X. Fitness costs of sulfoxaflor resistance in the cotton aphid, Aphis gossypii Glover. Pestic. Biochem. Physiol. 2019, 158, 40–46. [Google Scholar] [CrossRef]
- Longhurst, C.; Babcock, J.M.; Denholm, I.; Gorman, K.; Thomas, J.D.; Sparks, T.C. Cross-resistance relationships of the sulfoximine insecticide sulfoxaflor with neonicotinoids and other insecticides in the whiteflies Bemisia tabaci and Trialeurodes vaporariorum. Pest Manag. Sci. 2013, 69, 809–813. [Google Scholar] [CrossRef]
- Liao, X.; Jin, R.; Zhang, X.; Ali, E.; Mao, K.; Xu, P.; Li, J.; Wan, H. Characterization of sulfoxaflor resistance in the brown planthopper, Nilaparvata lugens (Stål). Pest Manag. Sci. 2018, 75, 1646–1654. [Google Scholar] [CrossRef]
- Zhang, X.; Mao, K.; Liao, X.; He, B.; Jin, R.; Tang, T.; Wan, H.; Li, J. Fitness cost of nitenpyram resistance in the brown planthopper Nilaparvata lugens. J. Pest Sci. 2018, 91, 1145–1151. [Google Scholar] [CrossRef]
- Feng, Y.T.; Wu, Q.J.; Xu, B.Y.; Wang, S.L.; Chang, X.L.; Xie, W.; Zhang, Y.J. Fitness costs and morphological change of laboratory-selected thiamethoxam resistance in the B-type Bemisia tabaci (Hemiptera: Aleyrodidae). J. Appl. Entomol. 2009, 133, 466–472. [Google Scholar] [CrossRef]
- Tieu, S.; Chen, Y.; Woolley, L.K.; Collins, D.; Barchia, I.; Lo, N.; Herron, G.A. A significant fitness cost associated with ACE1 target site pirimicarb resistance in a field isolate of Aphis gossypii Glover from Australian cotton. J. Pest Sci. 2016, 90, 773–779. [Google Scholar] [CrossRef]
- Shen, J.; Li, D.; Zhang, S.; Zhu, X.; Wan, H.; Li, J. Fitness and inheritance of metaflumizone resistance in Plutella xylostella. Pestic. Biochem. Physiol. 2017, 139, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liang, P.; Gao, X. Cross-resistance patterns and fitness in fufenozide-resistant diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2012, 68, 285–289. [Google Scholar] [CrossRef]
- Liu, Z.; Han, Z. Fitness costs of laboratory-selected imidacloprid resistance in the brown planthopper, Nilaparvata lugens Stål. Pest Manag. Sci. 2006, 62, 279–282. [Google Scholar] [CrossRef]
- Yu, J.Z.; Chi, H.; Chen, B.H. Comparison of the life tables and predation rates of Harmonia dimidiata (F.) (Coleoptera: Coccinellidae) fed on Aphis gossypii glover (Hemiptera: Aphididae) at different temperatures. Biol. Control 2013, 64, 1–9. [Google Scholar] [CrossRef]
- Alami, S.; Naseri, B.; Golizadeh, A.; Razmjou, J. Age-stage, two-sex life table of the tomato looper, Chrysodeixis chalcites (Lepidoptera: Noctuidae), on different bean cultivars. Arthropod-Plant Interact. 2014, 8, 475–484. [Google Scholar] [CrossRef]
- Wang, Z.H.; Gong, Y.J.; Chen, J.C.; Su, X.C.; Cao, L.J.; Hoffmann, A.A.; Wei, S.J. Laboratory selection for resistance to sulfoxaflor and fitness costs in the green peach aphid Myzus persicae. J. Asia-Pacif. Entomol. 2018, 21, 408–412. [Google Scholar] [CrossRef]
- Lu, Y.H.; He, Y.P.; Gao, X.W. Comparative studies on acetylcholinesterase characteristics between the aphids, Sitobion avenae and Rhopalosiphum padi. J. Insect Sci. 2013, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, J.; Fu, Y.; Francis, F.; Chen, J.L. Plant-mediated interactions between two cereal aphid species: Promotion ofaphid performance and attraction of more parasitoids by infestation of wheat with phytotoxic aphid Schizaphis graminum. J. Agric. Food Chem. 2019, 67, 2763–2773. [Google Scholar] [CrossRef]
- Hu, X.S.; Liu, X.F.; Thieme, T.; Zhang, G.S.; Liu, T.X.; Zhao, H.Y. Testing the fecundity advantage hypothesis with Sitobion avenae, Rhopalosiphum padi, and Schizaphis graminum (Hemiptera: Aphididae) feeding on ten wheat accessions. Sci. Rep. 2015, 5, 18549. [Google Scholar] [CrossRef]
- Gong, P.; Li, X.; Wang, C.; Zhu, S.; Li, Q.; Zhang, Y.; Li, X.; Li, G.; Liu, E.; Gao, H.; et al. The Sensitivity of field populations of Metopolophium dirhodum (Walker) (Hemiptera: Aphididae) to seven insecticides in Northern China. Agronomy 2021, 11, 1556. [Google Scholar] [CrossRef]
- Li, X.; Wang, C.; Li, Q.; Zhu, S.; Tian, X.; Zhang, Y.; Li, X.; Gao, H.; Liu, E.; Wang, L.; et al. Field-evolved sulfoxaflor resistance of three wheat aphid species in China. Agronomy 2021, 11, 2325. [Google Scholar] [CrossRef]
- Zuo, Y.; Wang, K.; Zhang, M.; Peng, X.; Piñero, J.C.; Chen, M. Regional susceptibilities of Rhopalosiphum padi (Hemiptera: Aphididae) to ten insecticides. Fla. Entomol. 2016, 99, 269–275. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlıhan, R.; Smith, C.; Kavousi, A.; Özgökçe, M.; Güncan, A.; Tuan, S.; Fu, J.; Xu, Y.; et al. Age-Stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Gong, P.; Li, X.; Gao, H.; Wang, C.; Li, M.; Zhang, Y.; Li, X.; Liu, E.; Zhu, X. Field evolved resistance to pyrethroids, neonicotinoids, organophosphates and macrolides in Rhopalosiphum padi (Linnaeus) and Sitobion avenae (Fabricius) from China. Chemosphere 2021, 269, 128747. [Google Scholar] [CrossRef]
- Zhang, X.L.; Liao, X.; Mao, K.K.; Zhang, K.X.; Wan, H.; Li, J.H. Insecticide resistance monitoring and correlation analysis of insecticides in field populations of the brown planthopper Nilaparvata lugens (stål) in China 2012–2014. Pestic. Biochem. Physiol. 2016, 132, 13–20. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. Available online: http://140.120.197.173/ecology/Papers/04-1985-Chi-Liu-2011%20ver.pdf (accessed on 1 November 2022).
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. Version 2022.10.25. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 1 November 2022).
- Akköprü, P.E.; Atlihan, R.; Okut, H.; Chi, H. Demographic assessment of plant cultivar resistance to insect pests: A case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol. 2015, 108, 378–387. [Google Scholar] [CrossRef]
- Huang, H.W.; Chi, H.; Smith, C.L. Linking demography and consumption of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae) fed on Solanum photeinocarpum (Solanales: Solanaceae): With a new method to project the uncertainty of population growth and consumption. J. Econ. Entomol. 2018, 111, 1–9. [Google Scholar] [CrossRef]
- Huang, Y.B.; Chi, H. Life tables of Bactrocera cucurbitae (Diptera: Tephritidae): With an invalidation of the jackknife technique. J. Appl. Entomol. 2013, 137, 327–339. [Google Scholar] [CrossRef]
- Wei, M.; Chi, H.; Guo, Y.; Li, X.; Zhao, L.; Ma, R. Demography of cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of pyrus bretschneideri (rosales: Rosaceae) and P. communis pears with estimations of confidence Intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Groters, F.R.; Tabashnik, B.E.; Finson, N.; Johnson, M.W. Fitness costs of resistance to Bacillus thuringiensis in the diamondback moth (Plutella xylostella). Evolution 1994, 48, 197–201. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Taylor, C.E. Genetic and biological influences in the evolution of insecticide resistance. J. Econ. Entomol. 1977, 70, 319–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferré, J.; Van Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef] [PubMed]
- Crespo, A.L.; Rodrigo-Simon, A.; Siqueira, H.A.; Pereira, E.J.; Ferre, J.; Siegfried, B.D. Cross-resistance and mechanism of resistance to Cry1Ab toxin from Bacillus thuringiensis in a field-derived strain of European corn borer, Ostrinia nubilalis. J. Invertebr. Pathol. 2011, 107, 185–192. [Google Scholar] [CrossRef]
- Zhang, H.; Tian, W.; Zhao, J.; Jin, L.; Yang, J.; Liu, C.; Yang, Y.; Wu, S.; Wu, K.; Cui, J.; et al. Diverse genetic basis of field-evolved resistance to Bt cotton in cotton bollworm from China. Proc. Natl. Acad. Sci. USA 2012, 109, 10275–10280. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, Y. High levels of resistance to chlorantraniliprole evolved in field populations of Plutella xylostella. J. Econ. Entomol. 2012, 105, 1019–1023. [Google Scholar] [CrossRef]
- Pu, X.; Yang, Y.; Wu, S.; Wu, Y. Characterisation of abamectin resistance in a field-evolved multiresistant population of Plutella xylostella. Pest Manag. Sci. 2010, 66, 371–378. [Google Scholar] [CrossRef]
- Tang, J.D.; Smadar, G.; Roush, R.T.; Shelton, M.N. Inheritance, stability, and lack-of-fitness costs of field-selected resistance to Bacillus thuringiensis in Diamondback Moth (Lepidoptera: Plutellidae) from Florida. J. Econ. Entomol. 1997, 90, 732–741. [Google Scholar] [CrossRef] [Green Version]
- Sayyed, A.H.; Omar, D.; Wright, D.J. Genetics of spinosad resistance in a multi-resistant field-selected population of Plutella xylostella. Pest Manag. Sci. 2004, 60, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Garrood, W.T.; Zimmer, C.T.; Gorman, K.J.; Nauen, R.; Bass, C.; Davies, T.G. Field-evolved resistance to imidacloprid and ethiprole in populations of brown planthopper Nilaparvata lugens collected from across South and East Asia. Pest Manag. Sci. 2016, 72, 140–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Mao, K.; Ali, E.; Jin, R.; Li, Z.; Li, W.; Li, J.; Wan, H. Inheritance and fitness costs of sulfoxaflor resistance in Nilaparvata lugens (Stal). Pest Manag. Sci. 2019, 75, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insecticide | Population | N a | Slope ± SE b | LC50 c (95% CI d; mg/L) | χ2 | p-Value | RR e |
|---|---|---|---|---|---|---|---|
| Sulfoxaflor | SS | 708 | 0.51 ± 0.11 | 1.7 (0.2–4.6) | 0.26 | 0.88 | |
| SulR | 585 | 0.53 ± 0.10 | 339.6 (138.1–1835.7) | 2.30 | 0.32 | 199.8 | |
| Imidacloprid | SS | 542 | 0.40 ± 0.06 | 37.4 (16.0–82.5) | 0.34 | 0.84 | |
| SulR | 598 | 0.42 ± 0.07 | 70.3 (33.62–184.7) | 3.10 | 0.21 | 1.9 | |
| Thiamethoxam | SS | 614 | 0.56 ± 0.06 | 23.3 (13.6–37.4) | 6.80 | 0.08 | |
| SulR | 567 | 0.67 ± 0.08 | 37.8 (23.8–62.9) | 0.79 | 0.85 | 1.6 | |
| Beta-cypermethrin | SS | 605 | 0.63 ± 0.06 | 4.8 (2.5–8.0) | 6.40 | 0.09 | |
| SulR | 541 | 0.77 ± 0.07 | 69.5 (45.9–112.9) | 1.00 | 0.80 | 14.5 | |
| Bifenthrin | SS | 587 | 0.66 ± 0.07 | 3.5 (1.7–6.0) | 5.54 | 0.14 | |
| SulR | 584 | 0.51 ± 0.07 | 147.4 (73.8–404.6) | 0.69 | 0.88 | 42.1 | |
| Abamectin | SS | 623 | 0.86 ± 0.10 | 12.2 (8.5–17.8) | 1.72 | 0.42 | |
| SulR | 645 | 1.31 ± 0.15 | 9.8 (7.0–14.0) | 0.56 | 0.76 | 0.8 | |
| Chlorpyrifos | SS | 595 | 2.28 ± 0.22 | 2.3 (1.9–2.8) | 5.29 | 0.15 | |
| SulR | 615 | 2.76 ± 0.42 | 13.2 (11.6–14.9) | 0.37 | 0.54 | 5.7 | |
| Omethoate | SS | 593 | 1.24 ± 0.11 | 97.5 (7.05–140.8) | 6.09 | 0.11 | |
| SulR | 655 | 1.46 ± 0.15 | 263.4 (192.4–386.9) | 0.30 | 0.86 | 2.7 |
| Parameter a | SS Population | SulR Population | p-Value | ||
|---|---|---|---|---|---|
| N b | Mean ± SE | N | Mean ± SE | ||
| L1 (d) | 120 | 1.75 ± 0.06 a | 120 | 1.75 ± 0.08 a | 0.9705 |
| L2 (d) | 113 | 2.01 ± 0.09 a | 112 | 2.08 ± 0.08 a | 0.5565 |
| L3 (d) | 110 | 1.84 ± 0.07 a | 109 | 2.02 ± 0.08 a | 0.0940 |
| L4 (d) | 109 | 1.72 ± 0.07 b | 103 | 2.08 ± 0.07 a | 0.0005 |
| Pre-adult (d) | 109 | 7.24 ± 0.10 b | 103 | 7.73 ± 0.18 a | 0.0173 |
| Adult longevity (d) | 109 | 11.34 ± 0.42 a | 103 | 10.34 ± 0.46 a | 0.1075 |
| Total longevity (d) | 118 | 17.66 ± 0.48 a | 118 | 16.93 ± 0.48 a | 0.2851 |
| APRP (d) | 107 | 0.45 ± 0.06 b | 97 | 0.64 ± 0.06 a | 0.0328 |
| TPRP (d) | 107 | 7.63 ± 0.11 b | 97 | 8.20 ± 0.17 a | 0.0055 |
| Reproductive period (d) | 109 | 7.50 ± 0.32 a | 103 | 6.74 ± 0.31 a | 0.0889 |
| Fecundity (offspring/female) | 109 | 18.40 ± 0.98 a | 103 | 14.22 ± 1.00 b | 0.0031 |
| Parameter | SS Population | SulR Population | p-Value | ||
|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | ||
| Net reproductive rate (R0) | 120 | 17.00 ± 1.00 a | 120 | 12.41 ± 0.98 b | 0.0011 |
| Mean generation time (T) | 120 | 10.93 ± 0.14 a | 120 | 11.28 ± 0.14 a | 0.0790 |
| Intrinsic rate of increase (r) | 120 | 0.26 ± 0.01 a | 120 | 0.22 ± 0.01 b | 0.0003 |
| Finite rate of increase (λ) | 120 | 1.30 ± 0.01 a | 120 | 1.25 ± 0.01 b | 0.0003 |
| Relative fitness (Rf) a | 0.73 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhu, S.; Li, Q.; Sun, Y.; Wang, Y.; Tian, X.; Ran, X.; Li, X.; Zhang, Y.; Gao, H.; et al. Fitness Cost of the Field-Evolved Resistance to Sulfoxaflor and Multi-Insecticide Resistance of the Wheat Aphid Sitobion miscanthi (Takahashi). Insects 2023, 14, 75. https://doi.org/10.3390/insects14010075
Li X, Zhu S, Li Q, Sun Y, Wang Y, Tian X, Ran X, Li X, Zhang Y, Gao H, et al. Fitness Cost of the Field-Evolved Resistance to Sulfoxaflor and Multi-Insecticide Resistance of the Wheat Aphid Sitobion miscanthi (Takahashi). Insects. 2023; 14(1):75. https://doi.org/10.3390/insects14010075
Chicago/Turabian StyleLi, Xinan, Saige Zhu, Qiuchi Li, Yulin Sun, Yanbo Wang, Xujun Tian, Xiao Ran, Xiangrui Li, Yunhui Zhang, Haifeng Gao, and et al. 2023. "Fitness Cost of the Field-Evolved Resistance to Sulfoxaflor and Multi-Insecticide Resistance of the Wheat Aphid Sitobion miscanthi (Takahashi)" Insects 14, no. 1: 75. https://doi.org/10.3390/insects14010075
APA StyleLi, X., Zhu, S., Li, Q., Sun, Y., Wang, Y., Tian, X., Ran, X., Li, X., Zhang, Y., Gao, H., & Zhu, X. (2023). Fitness Cost of the Field-Evolved Resistance to Sulfoxaflor and Multi-Insecticide Resistance of the Wheat Aphid Sitobion miscanthi (Takahashi). Insects, 14(1), 75. https://doi.org/10.3390/insects14010075