
Orthopteran Diversity in Steep Slope Vineyards: The Role of Vineyard Type and Vegetation Management


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
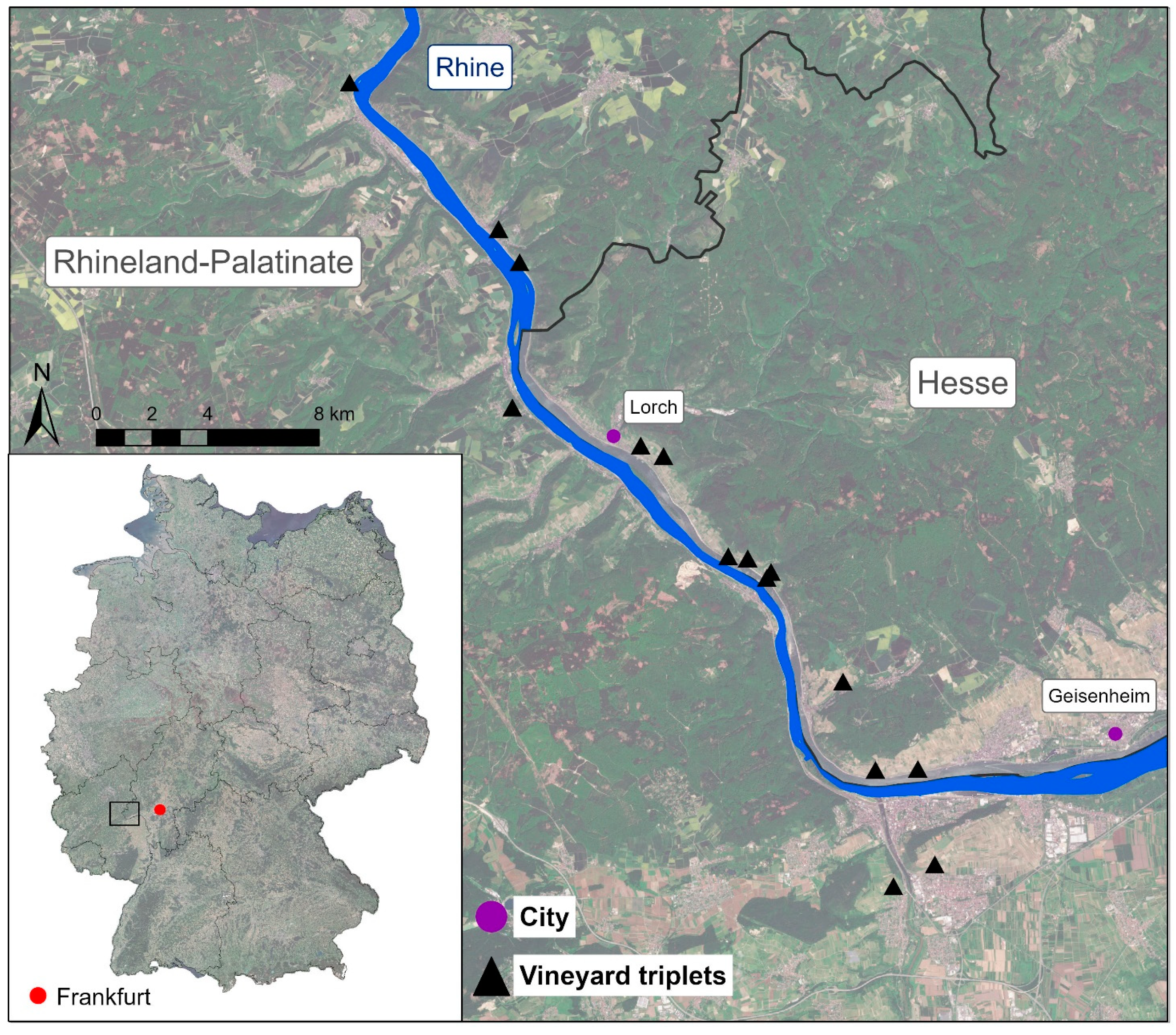
2.1. Study Area and Sampling Design
- (i)
- vertically oriented vineyards received an alternating tillage treatment, i.e., every second inter-row was kept open by regular tillage while the other inter-row was permanently covered with grassy vegetation dominated by Lolium perenne. In order to prevent competition for water and nutrients with vines, inter-row vegetation was kept short by regular mulching.
- (ii)
- terraced vineyards received a regular tillage treatment in inter-rows, while terrace embankments were permanently covered with grassy and herbaceous vegetation such as Arrhenaterum elatius, Bromus erectus, Galium album, and Isatis tinctoria. Embankment vegetation was extensively managed and mulched/mown once in summer.
- (iii)
- vineyard fallows were abandoned for at least 10 years and were mostly overgrown with woody vegetation dominated by Rubus fruticosus agg., Rosa canina agg., Prunus avium, and Crataegus laevigata.
2.2. Orthoptera Sampling
2.3. Environmental Variables and Landscape Analysis
2.4. Data Analysis
3. Results
3.1. Local Vegetation Parameters
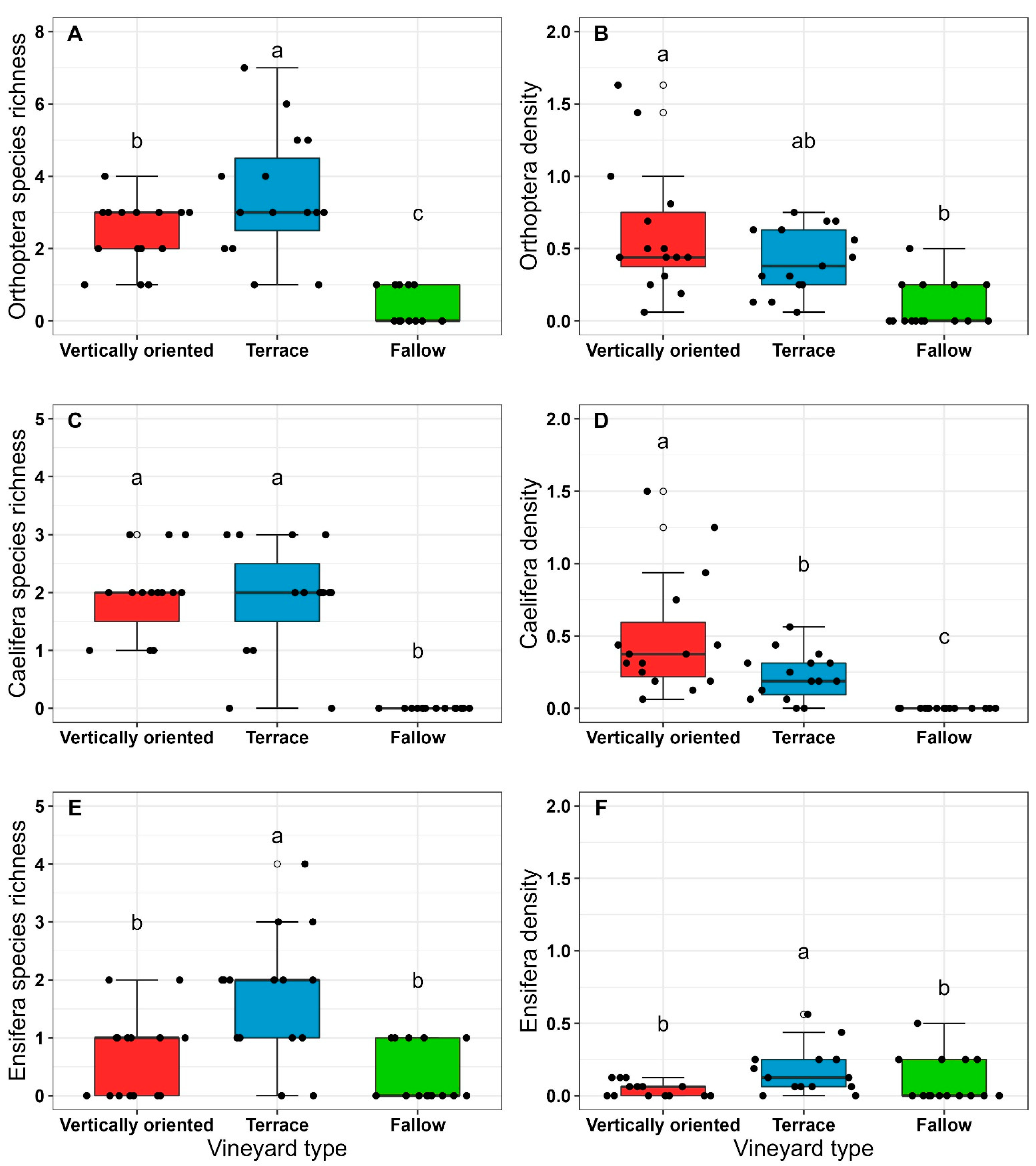
3.2. Orthoptera Diversity in Vineyard Types
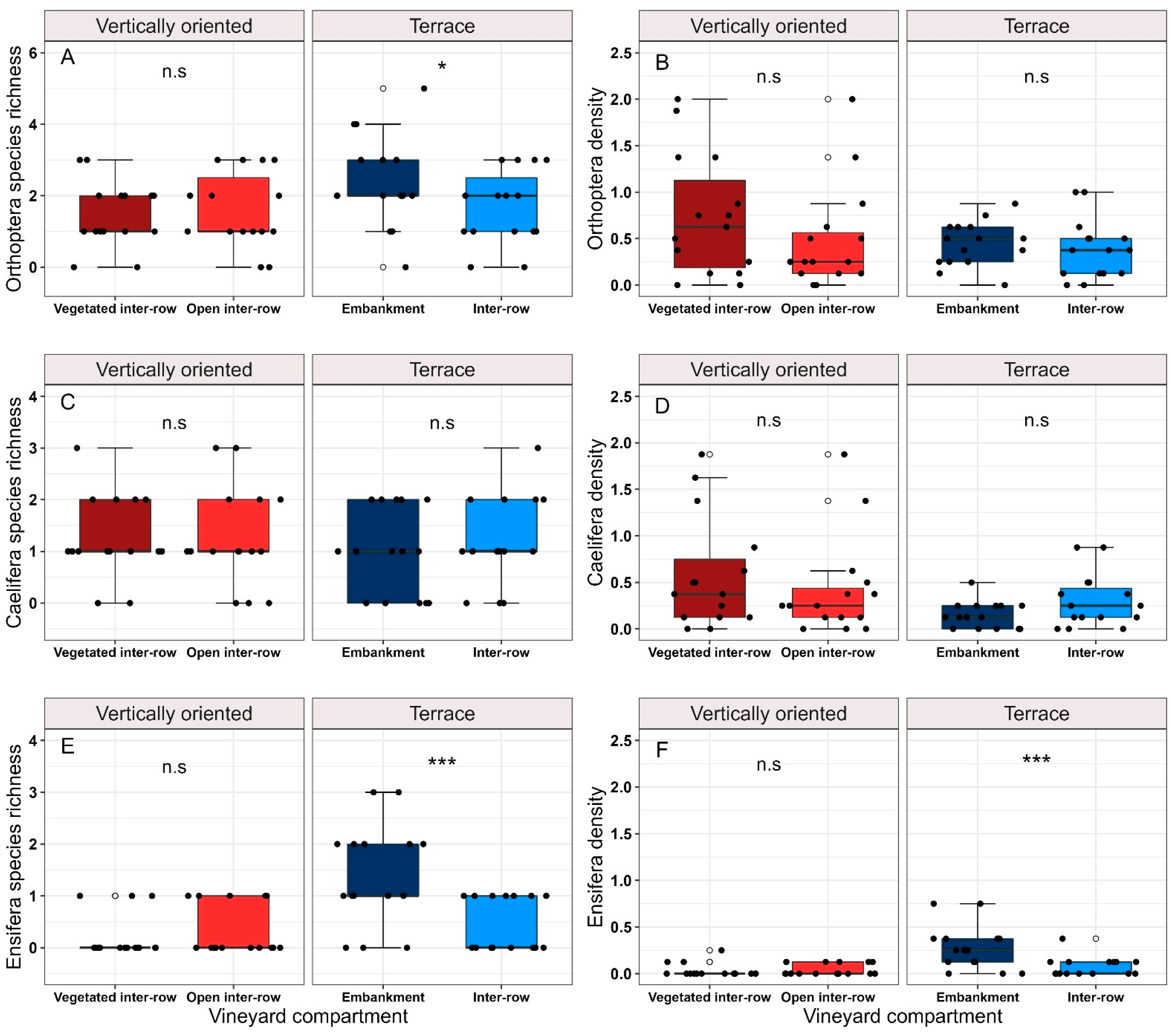
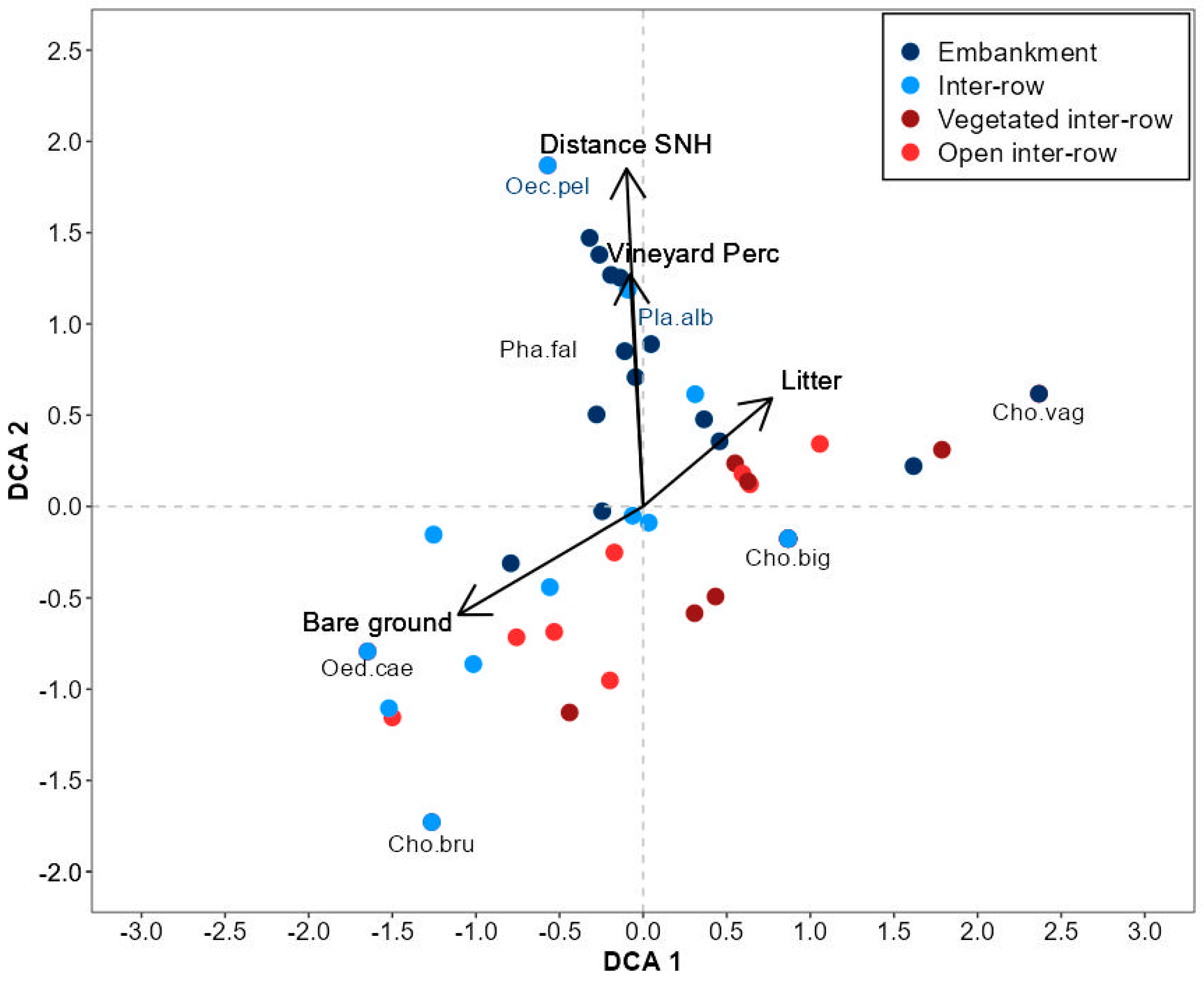
3.3. Orthoptera Diversity in Vineyard Compartments
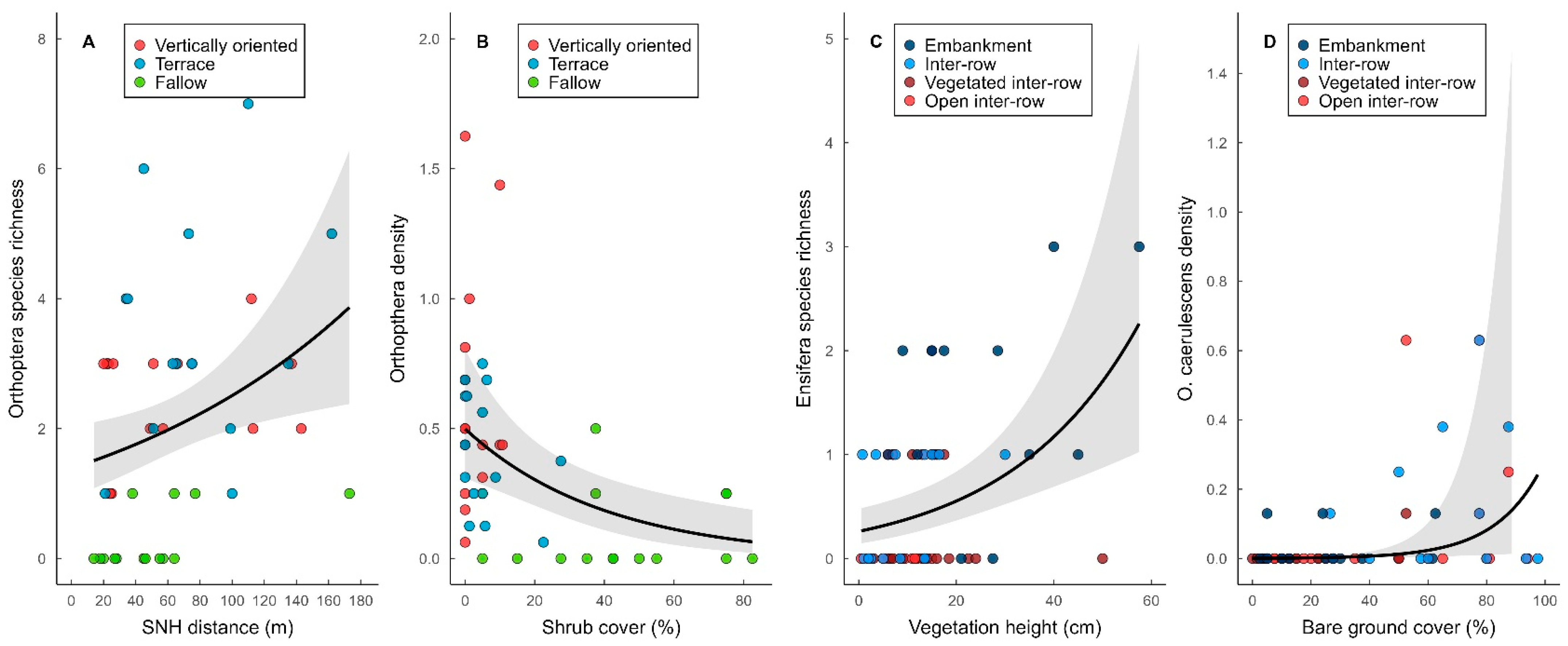
3.4. Effect of Local and Landscape Parameters on Orthoptera
4. Discussion
4.1. Effects of Vineyard Abandonment on Orthoptera Diversity
4.2. Effects of Local Vineyard Management on Orthoptera Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Harvey, J.A.; Heinen, R.; Armbrecht, I.; Basset, Y.; Baxter-Gilbert, J.H.; Bezemer, T.M.; Böhm, M.; Bommarco, R.; Borges, P.A.V.; Cardoso, P.; et al. International scientists formulate a roadmap for insect conservation and recovery. Nat. Ecol. Evol. 2020, 4, 174–176. [Google Scholar] [CrossRef]
- Thomas, C.D.; Jones, T.H.; Hartley, S.E. “Insectageddon”: A call for more robust data and rigorous analyses. Glob. Chang. Biol. 2019, 25, 1891–1892. [Google Scholar] [CrossRef] [Green Version]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Sutcliffe, L.M.E.; Batáry, P.; Kormann, U.; Báldi, A.; Dicks, L.V.; Herzon, I.; Kleijn, D.; Tryjanowski, P.; Apostolova, I.; Arlettaz, R.; et al. Harnessing the biodiversity value of Central and Eastern European farmland. Divers. Distrib. 2015, 21, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Kleijn, D.; Kohler, F.; Baldi, A.; Batary, P.; Concepción, E.; Clough, Y.; Díaz, M.; Gabriel, D.; Holzschuh, A.; Knop, E.; et al. On the relationship between farmland biodiversity and land-use intensity in Europe. Proc. Biol. Sci. 2009, 276, 903–909. [Google Scholar] [CrossRef]
- Marini, L.; Klimek, S.; Battisti, A. Mitigating the impacts of the decline of traditional farming on mountain landscapes and biodiversity: A case study in the European Alps. Environ. Sci. Policy 2011, 14, 258–267. [Google Scholar] [CrossRef]
- Plieninger, T.; Höchtl, F.; Spek, T. Traditional land-use and nature conservation in European rural landscapes. Environ. Sci. Policy 2006, 9, 317–321. [Google Scholar] [CrossRef]
- Treiling, T. Kulturlandschaftswandel im Oberen Mittelrheintal: Historische Landschaftsanalyse mit GIS; VDM Verlag Dr. Müller: Saarbrücken, Germany, 2008; ISBN 9783639020793. [Google Scholar]
- Veith, M.; Bonn, S.; Sander, U.; Albrech, J.; Poschlod, P. Nachhaltige Entwicklung Xerothermer Hanglagen am Beispiel des Mittelrheintals: Eine Naturschutzfachliche, Ökonomische und Sozio-Kulturelle Bewertung; Ergebnisse des gleichnamigen E+E-Vorhabens; Bundesamt für Naturschutz: Bonn-Bad Godesberg, Germany, 2012; ISBN 9783784340210. [Google Scholar]
- Strub, L.; Kurth, A.; Loose, S.M. Effects of viticultural mechanization on working time requirements and production costs. Am. J. Enol. Vitic. 2021, 72, 46–55. [Google Scholar] [CrossRef]
- Strub, L.; Loose, S.M. The cost disadvantage of steep slope viticulture and strategies for its preservation. OENO One 2021, 55, 49–68. [Google Scholar] [CrossRef]
- Stoll, M.; Schultz, H.-R. Deutsches Weinbaujahrbuch 2021; Verlag Eugen Ulmer: Stuttgart, Germany, 2020; ISBN 978-3-8186-1181-1. [Google Scholar]
- Statistisches Landesamt Rheinland-Pfalz. Landwirtschaft: Zeitreihen regional: Bestockte Rebfläche der Keltertrauben 1989–2020 nach Ausgewählten Rebsorten und Anbaugebieten. Available online: https://www.statistik.rlp.de/de/wirtschaftsbereiche/landwirtschaft/zeitreihen-regional/tabelle-4/ (accessed on 8 May 2021).
- Strub, L.; Loose, S. Quer arbeiten, Kosten sparen. Dtsch. Weinbau 2021, 14–19. [Google Scholar]
- Wersebeckmann, V.; Kolb, S.; Entling, M.H.; Leyer, I. Maintaining steep slope viticulture for spider diversity. Glob. Ecol. Conserv. 2021, 29, e01727. [Google Scholar] [CrossRef]
- Báldi, A.; Kisbenedek, T. Orthopteran assemblages as indicators of grassland naturalness in Hungary. Agric. Ecosyst. Environ. 1997, 66, 121–129. [Google Scholar] [CrossRef]
- Gardiner, T.; Dover, J. Is microclimate important for Orthoptera in open landscapes? J. Insect Conserv. 2008, 12, 705–709. [Google Scholar] [CrossRef]
- Badenhausser, I.; Cordeau, S. Sown grass strip—A stable habitat for grasshoppers (Orthoptera: Acrididae) in dynamic agricultural landscapes. Arthropod Plant Interact 2015, 9, 333–346. [Google Scholar] [CrossRef]
- Vickery, J.A.; Tallowin, J.R.; Feber, R.E.; Asteraki, E.J.; Atkinson, P.W.; Fuller, R.J.; Brown, V.K. The management of lowland neutral grasslands in Britain: Effects of agricultural practices on birds and their food resources. J. Appl. Ecol. 2001, 38, 647–664. [Google Scholar] [CrossRef]
- Fartmann, T.; Krämer, B.; Stelzner, F.; Poniatowski, D. Orthoptera as ecological indicators for succession in steppe grassland. Ecol. Indic. 2012, 20, 337–344. [Google Scholar] [CrossRef]
- Marini, L.; Fontana, P.; Scotton, M.; Klimek, S. Vascular plant and Orthoptera diversity in relation to grassland management and landscape composition in the European Alps. J. Appl. Ecol. 2008, 45, 361–370. [Google Scholar] [CrossRef]
- Weiss, N.; Zucchi, H.; Hochkirch, A. The effects of grassland management and aspect on Orthoptera diversity and abundance: Site conditions are as important as management. Biodivers. Conserv. 2013, 22, 2167–2178. [Google Scholar] [CrossRef]
- Schirmel, J.; Mantilla-Contreras, J.; Blindow, I.; Fartmann, T. Impacts of succession and grass encroachment on heathland Orthoptera. J. Insect Conserv. 2011, 15, 633–642. [Google Scholar] [CrossRef]
- Hochschule Geisenheim University. HGU|Monatsauswertung der Wetterstationen. Available online: https://rebschutz.hs-geisenheim.de/wetterstationen/monatsauswertung.php (accessed on 28 April 2021).
- Mody, K.; Lerch, D.; Müller, A.-K.; Simons, N.K.; Blüthgen, N.; Harnisch, M. Flower power in the city: Replacing roadside shrubs by wildflower meadows increases insect numbers and reduces maintenance costs. PLOS ONE 2020, 15, e0234327. [Google Scholar] [CrossRef]
- Fischer, J.; Steinlechner, D.; Zehm, A.; Poniatowski, D.; Fartmann, T.; Beckmann, A.; Stettmer, C. Die Heuschrecken Deutschlands und Nordtirols: Bestimmen–Beobachten–Schützen, 2., Korrigierte Auflage; Quelle & Meyer Verlag: Wiebelsheim, Germany, 2020; ISBN 9783494017952. [Google Scholar]
- Horstkotte, J.; Lorenz, C.; Wendler, A. Heuschrecken: Bestimmung, Verbreitung, Lebensräume und Gefährdung aller in Deutschland Vorkommenden Arten, 13., unveränd. Aufl.; Dt. Jugendbund für Naturbeobachtung: Hamburg, Germany, 1999; ISBN 3923376030. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means: Version 1.4.8. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 5 September 2021).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package: Version 2.5–6. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 3 August 2021).
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; ISBN 9780511615146. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 3 August 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 3 August 2021).
- Wilke, C.O.; Cowplot: Streamlined Plot Theme and Plot Annotations for ’ggplot2’. R Package Version 1.1.1. Available online: https://CRAN.R-project.org/package=cowplot (accessed on 3 August 2021).
- ESRI. ArcMap 10.7. Available online: https://desktop.arcgis.com/de/arcmap/ (accessed on 10 May 2021).
- Inkscape, version 1.2.1; Inkscape Project, 2020. Available online: https://inkscape.org. (accessed on 15 October 2022).
- Maas, S.; Detzel, P.; Staudt, A. Rote Liste und Gesamtartenliste der Heuschrecken (Saltatoria) Deutschlands. In Rote Liste Gefährdeter Tiere, Pflanzen und Pilze Deutschlands: Band 70 (3): Wirbellose Tiere (Teil 1); Binot-Hafke, M., Balzer, S., Becker, N., Gruttke, H., Haupt, H., Hofbauer, N., Ludwig, G., Matzke-Hajek, G., Strauch, M., Eds.; Bundesamt für Naturschutz: Bonn-Bad Godesberg, Germany, 2011; pp. 577–606. ISBN 9783784352312. [Google Scholar]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Branson, D.H. Relationships between Plant Diversity and Grasshopper Diversity and Abundance in the Little Missouri National Grassland. Psyche J. Entomol. 2011, 2011, 748635. [Google Scholar] [CrossRef]
- Bundschuh, R.; Schmitz, J.; Bundschuh, M.; Brühl, C.A. Does insecticide drift adversely affect grasshoppers (Orthoptera: Saltatoria) in field margins? A case study combining laboratory acute toxicity testing with field monitoring data. Environ. Toxicol. Chem. 2012, 31, 1874–1879. [Google Scholar] [CrossRef]
- Marini, L.; Fontana, P.; Battisti, A.; Gaston, K.J. Response of orthopteran diversity to abandonment of semi-natural meadows. Agric. Ecosyst. Environ. 2009, 132, 232–236. [Google Scholar] [CrossRef]
- Willott, S.J.; Hassall, M. Life-history responses of British grasshoppers (Orthoptera: Acrididae) to temperature change. Funct. Ecol. 1998, 12, 232–241. [Google Scholar] [CrossRef]
- Poniatowski, D.; Fartmann, T. The classification of insect communities: Lessons from orthopteran assemblages of semi-dry calcareous grasslands in central Germany. Eur. J. Èntomol. 2008, 105, 659–671. [Google Scholar] [CrossRef]
- Gardiner, T.; Pye, M.; Field, R.; Hill, J. The influence of sward height and vegetation composition in determining the habitat preferences of three Chortippus species (Orthoptera: Acrididae) in Chelmsford, Essex, UK. J. Orthoptera Res. 2002, 11, 207–213. [Google Scholar] [CrossRef]
- Bieringer, G.; Zulka, K.P. Shading out species richness: Edge effect of a pine plantation on the Orthoptera (Tettigoniidae and Acrididae) assemblage of an adjacent dry grassland. Biodivers. Conserv. 2003, 12, 1481–1495. [Google Scholar] [CrossRef]
- Van Wingerden, W.K.R.E.; Musters, J.C.M.; Maaskamp, F.I.M. The influence of temperature on the duration of egg development in West European grasshoppers (Orthoptera: Acrididae). Oecologia 1991, 87, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Ortis, G.; Mazzon, L.; Cavaletto, G.; Marangoni, F.; Marini, L. Can extensively managed perennial crops serve as surrogate habitat for orthopterans typical of dry calcareous grasslands? Agric. Ecosyst. Environ. 2021, 319, 107536. [Google Scholar] [CrossRef]
- Steck, C.E.; Bürgi, M.; Bolliger, J.; Kienast, F.; Lehmann, A.; Gonseth, Y. Conservation of grasshopper diversity in a changing environment. Biol. Conserv. 2007, 138, 360–370. [Google Scholar] [CrossRef]
- Gardiner, T.; Hill, J.; Chesmore, D. Review of the methods frequently used to estimate the abundance of Orthoptera in grassland ecosystems. J. Insect Conserv. 2005, 9, 151–173. [Google Scholar] [CrossRef]
- Uzman, D.; Reineke, A.; Entling, M.H.; Leyer, I. Habitat area and connectivity support cavity-nesting bees in vineyards more than organic management. Biol. Conserv. 2020, 242, 108419. [Google Scholar] [CrossRef]
- Pithon, J.A.; Beaujouan, V.; Daniel, H.; Pain, G.; Vallet, J. Are vineyards important habitats for birds at local or landscape scales? Basic Appl. Ecol. 2016, 17, 240–251. [Google Scholar] [CrossRef]
- Assandri, G.; Bogliani, G.; Pedrini, P.; Brambilla, M. Assessing common birds’ ecological requirements to address nature conservation in permanent crops: Lessons from Italian vineyards. J. Environ. Manag. 2017, 191, 145–154. [Google Scholar] [CrossRef]
- Bonari, G.; Fajmon, K.; Malenovský, I.; Zelený, D.; Holuša, J.; Jongepierová, I.; Kočárek, P.; Konvička, O.; Uřičář, J.; Chytrý, M. Management of semi-natural grasslands benefiting both plant and insect diversity: The importance of heterogeneity and tradition. Agric. Ecosyst. Environ. 2017, 246, 243–252. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Raemakers, I.P.; Sýkora, K.V.; ter Braak, C. Arthropod assemblages are best predicted by plant species composition. Ecology 2008, 89, 782–794. [Google Scholar] [CrossRef]
- Poniatowski, D.; Stuhldreher, G.; Löffler, F.; Fartmann, T. Patch occupancy of grassland specialists: Habitat quality matters more than habitat connectivity. Biol. Conserv. 2018, 225, 237–244. [Google Scholar] [CrossRef]
- Batáry, P.; Orci, K.M.; Báldi, A.; Kleijn, D.; Kisbenedek, T.; Erdős, S. Effects of local and landscape scale and cattle grazing intensity on Orthoptera assemblages of the Hungarian Great Plain. Basic Appl. Ecol. 2007, 8, 280–290. [Google Scholar] [CrossRef]
- Ingrisch, S. Labor- und Freilanduntersuchungen zur Dauer der postembryonalen Entwicklung einiger mitteleuropäischer Laubheuschrecken (Orthoptera: Tettigoniidae) und ihre Beeinflussung durch Temperatur und Feuchte. Zool. Anz 1978, 200, 309–320. [Google Scholar]
- Ingrisch, S.; Köhler, G. Die Heuschrecken Mitteleuropas; Westarp Wissenschaften: Magdeburg, Germany, 1998; ISBN 3894324619. [Google Scholar]
- Gaigher, R.; Samways, M.J. Surface-active arthropods in organic vineyards, integrated vineyards and natural habitat in the Cape Floristic Region. J. Insect Conserv. 2010, 14, 595–605. [Google Scholar] [CrossRef]
- Adu-Acheampong, S.; Bazelet, C.S.; Samways, M.J. Extent to which an agricultural mosaic supports endemic species-rich grasshopper assemblages in the Cape Floristic Region biodiversity hotspot. Agric. Ecosyst. Environ. 2016, 227, 52–60. [Google Scholar] [CrossRef]
- Uchida, K.; Ushimaru, A. Biodiversity declines due to abandonment and intensification of agricultural lands: Patterns and mechanisms. Ecol. Monogr. 2014, 84, 637–658. [Google Scholar] [CrossRef]
- Gardiner, T.; Hassall, M. Does microclimate affect grasshopper populations after cutting of hay in improved grassland? J. Insect Conserv. 2009, 13, 97–102. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Ghazoul, J.; Richner, N.; Walter, T. Uncut grass refuges mitigate the impact of mechanical meadow harvesting on orthopterans. Biol. Conserv. 2012, 152, 96–101. [Google Scholar] [CrossRef]
- Reiff, J.; Kolb, S.; Entling, M.; Herndl, T.; Möth, S.; Walzer, A.; Kropf, M.; Hoffmann, C.; Winter, S. Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards. Insects 2021, 12, 220. [Google Scholar] [CrossRef]
- Grabenweger, G.; Kehrli, P.; Schlick-Steiner, B.; Steiner, F.; Stolz, M.; Bacher, S. Predator complex of the horse chestnut leafminer Cameraria ohridella: Identification and impact assessment. J. Appl. Èntomol. 2005, 129, 353–362. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Managed Vineyards | Abandoned Vineyards | ||
|---|---|---|---|
| Variable | Vertically Oriented | Terrace | Fallow |
| Vegetation height xx [cm] | 12.43 b ± 1.80 | 16.43 b ± 2.17 | 69.50 a ± 9.73 |
| Bare ground cover [%] | 36.87 a ± 5.45 | 39.98 a ± 4.71 | 11.83 b ± 2.33 |
| Litter cover [%] | 37.48 a ± 4.90 | 37.99 a ± 4.83 | 31.00 b ± 4.29 |
| Vegetation cover xx [%] | 22.80 a ± 4.05 | 16.03 ab ± 3.35 | 13.17 b ± 4.82 |
| Shrub cover x [%] | 2.80 b ± 1.09 | 6.00 b ± 2.13 | 44.00 a ± 6.49 |
| Vertically Oriented | Terrace | |||
|---|---|---|---|---|
| Variable | Vegetated Inter-Row | Open Inter-Row | Embankment | Inter-Row |
| Vegetation height [cm] | 15.37 a ± 2.92 | 9.50 b ± 1.13 | 23.87 a ± 3.82 | 8.98 b ± 2.09 |
| Bare ground cover [%] | 20.90 b ± 5.93 | 52.67 a ± 7.32 | 21.80 b ± 5.05 | 58.17 a ± 7.25 |
| Litter cover [%] | 46.50 a ± 6.4 | 28.13 b ± 6.07 | 50.33 a ± 6.56 | 25.65 b ± 5.49 |
| Vegetation cover xx [%] | 27.33 a ± 5.06 | 18.10 b ± 3.64 | 16.53 ± 4.75 | 15.52 ± 3.15 |
| Shrub cover xx [%] | 4.50 a ± 1.73 | 1.10 b ± 0.81 | 11.33 a ± 4.30 | 0.67 b ± 0.52 |
| Species | Vertically Oriented | Terrace | Fallow | p |
|---|---|---|---|---|
| Platycleis albopunctata | 0.53 ± 0.17 | 1.00 ± 0.22 | 0.00 ± 0.00 | 0.0055 |
| Oecanthus pellucens | 0.13 ± 0.09 | 1.33 ± 0.53 | 0.20 ± 0.14 | 0.0056 |
| Oedipoda caerulescens | 0.93 ± 0.47 | 1.20 ± 0.44 | 0.00 ± 0.00 | 0.0451 |
| Phaneroptera falcata | 0.00 ± 0.00 | 0.33 ± 0.16 | 0.00 ± 0.00 | 0.0278 |
| Chorthippus biguttulus | 5.80 ± 1.63 | 1.67 ± 0.40 | 0.00 ± 0.00 | 0.0005 |
| Vertically Oriented | Terrace | ||||
|---|---|---|---|---|---|
| Species | Vegetated Inter-Row | Open Inter-Row | Embankment | Inter-Row | p |
| Platycleis albopunctata | 0.27 ± 0.15 | 0.27 ± 0.12 | 0.93 ± 0.23 | 0.07 ± 0.07 | 0.0023 |
| Oecanthus pellucens | 0.00 ± 0.00 | 0.07 ± 0.07 | 0.87 ± 0.35 | 0.40 ± 0.21 | 0.0103 |
| Dependent Variable | Predictor | Estimate ± SE | p-Value | R2marg | R2cond | AICc |
|---|---|---|---|---|---|---|
| Orthoptera species richness | SNH (%) | −0.04 ± 0.12 | 0.7390 | 0.03 | 0.03 | 175.0 |
| Vineyard (%) | −0.01 ± 0.13 | 0.9710 | 0.01 | 0.01 | 175.1 | |
| Forest (%) | 0.09 ± 0.13 | 0.4620 | 0.02 | 0.02 | 174.6 | |
| SNH distance (m) | 0.25± 0.09 | 0.0086 | 0.14 | 0.01 | 170.3 | |
| Forest distance (m) | −0.15 ± 0.12 | 0.1840 | 0.06 | 0.01 | 174.8 | |
| Vegetation height (cm) | −0.85± 0.23 | 0.0002 | 0.63 | 0.64 | 153.3 | |
| Bare ground cover (%) | 0.32± 0.12 | 0.0081 | 0.18 | 0.18 | 168.2 | |
| Vegetation cover (%) | 0.21 ± 0.12 | 0.0969 | 0.07 | 0.07 | 172.4 | |
| Shrub cover (%) | −0.85± 0.21 | <0.0001 | 0.64 | 0.64 | 150.6 | |
| Litter cover (%) | 0.07 ± 0.13 | 0.5820 | 0.01 | 0.01 | 174.8 | |
| Orthoptera density | SNH (%) | −0.01 ± 0.05 | 0.7650 | 0.02 | 0.02 | 43.5 |
| Vineyard (%) | 0.06 ± 0.05 | 0.2910 | 0.03 | 0.03 | 42.5 | |
| Forest (%) | −0.01 ± 0.05 | 0.9150 | 0.01 | 0.01 | 43.6 | |
| SNH distance (m) | 0.08 ± 0.05 | 0.1240 | 0.05 | 0.00 | 41.3 | |
| Forest distance (m) | −0.02 ± 0.05 | 0.6470 | 0.01 | 0.01 | 43.4 | |
| Vegetation height (cm) | −0.18± 0.05 | 0.0002 | 0.23 | 0.39 | 32.7 | |
| Bare ground cover (%) | 0.10 ± 0.05 | 0.0585 | 0.08 | 0.12 | 40.2 | |
| Vegetation cover (%) | 0.12± 0.05 | 0.0177 | 0.13 | 0.13 | 38.3 | |
| Shrub cover (%) | −0.17± 0.05 | 0.0004 | 0.21 | 0.31 | 33.2 | |
| Litter cover (%) | −0.01 ± 0.05 | 0.9560 | 0.01 | 0.01 | 43.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wersebeckmann, V.; Biegerl, C.; Leyer, I.; Mody, K. Orthopteran Diversity in Steep Slope Vineyards: The Role of Vineyard Type and Vegetation Management. Insects 2023, 14, 83. https://doi.org/10.3390/insects14010083
Wersebeckmann V, Biegerl C, Leyer I, Mody K. Orthopteran Diversity in Steep Slope Vineyards: The Role of Vineyard Type and Vegetation Management. Insects. 2023; 14(1):83. https://doi.org/10.3390/insects14010083
Chicago/Turabian StyleWersebeckmann, Vera, Carolin Biegerl, Ilona Leyer, and Karsten Mody. 2023. "Orthopteran Diversity in Steep Slope Vineyards: The Role of Vineyard Type and Vegetation Management" Insects 14, no. 1: 83. https://doi.org/10.3390/insects14010083
APA StyleWersebeckmann, V., Biegerl, C., Leyer, I., & Mody, K. (2023). Orthopteran Diversity in Steep Slope Vineyards: The Role of Vineyard Type and Vegetation Management. Insects, 14(1), 83. https://doi.org/10.3390/insects14010083