
Mechanical Egg Activation and Rearing of First Instar Larvae of Sirex noctilio (Hymenoptera: Siricidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
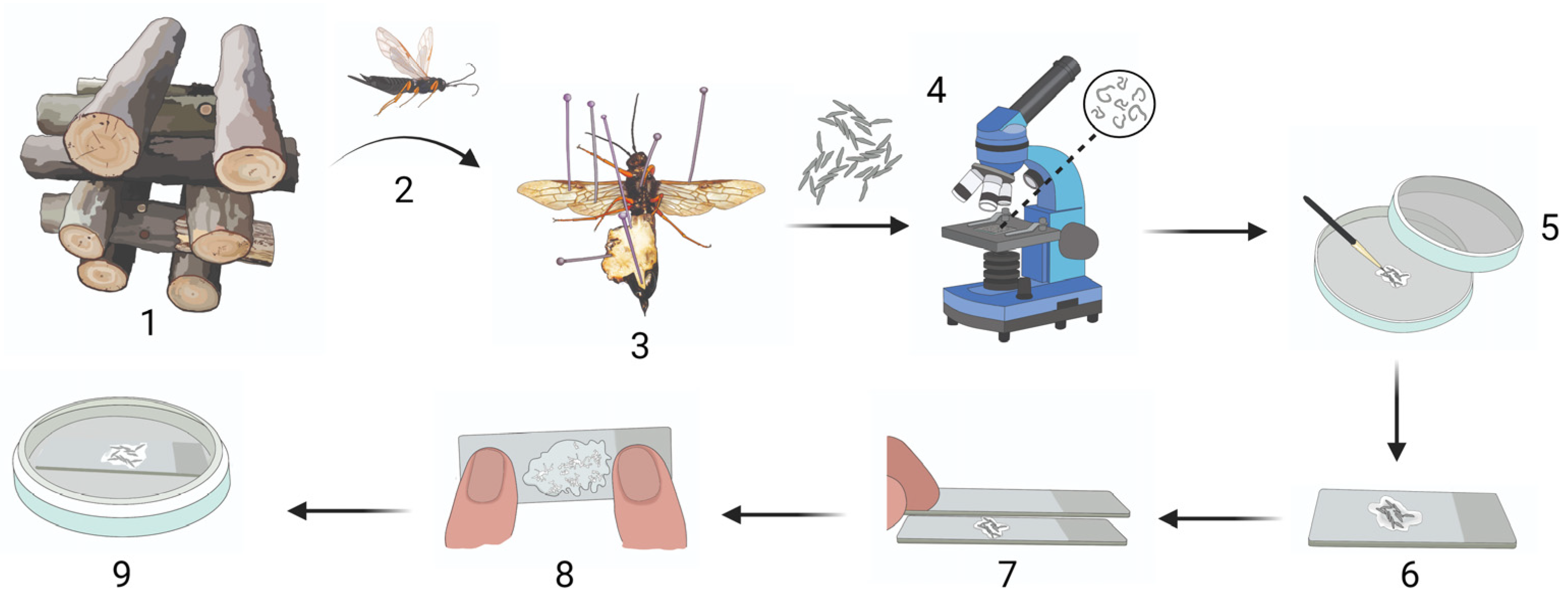
2.1. Insect Collection
2.2. Dissection and Extraction of Eggs
2.3. Egg Activation
2.4. Egg Activation and Subsequent Development with Treatments
2.5. Rearing of Sirex Noctilio Larvae, Emerging from Activated Eggs, on a Modified Artificial Diet
2.6. Statistical Analysis
3. Results
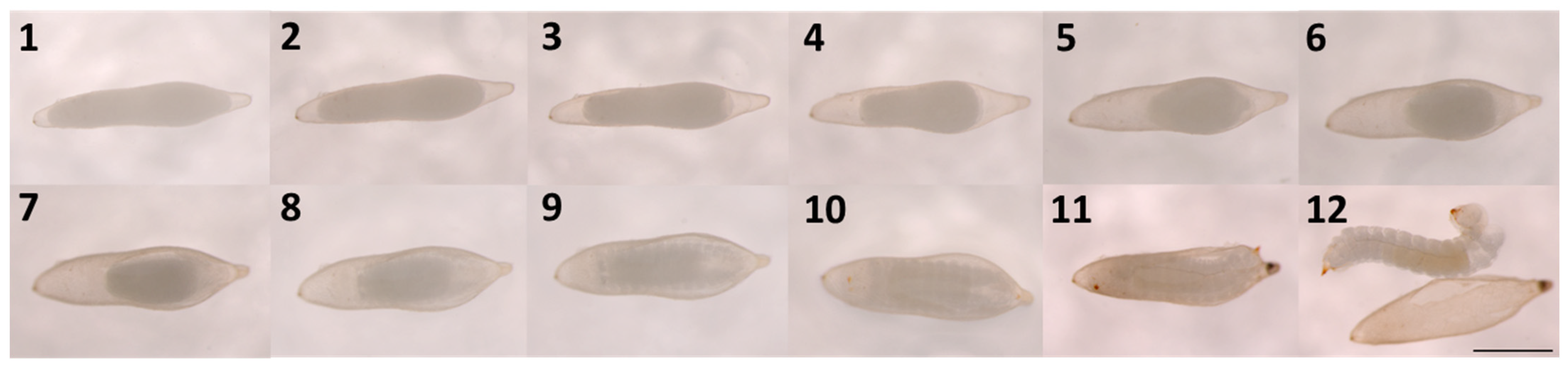
3.1. Egg Activation
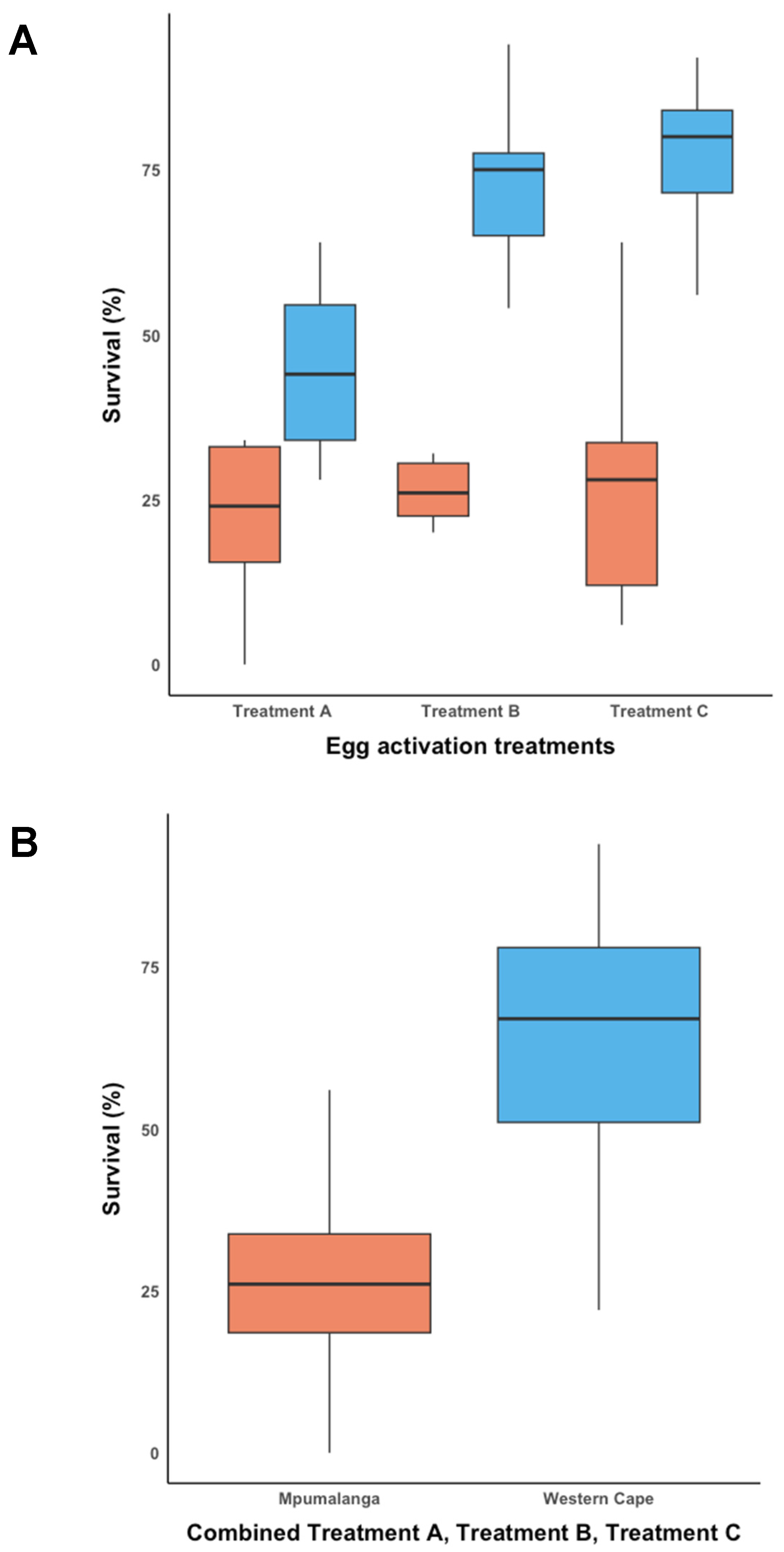
3.2. Egg Activation and Subsequent Development with Treatments
3.3. Rearing of Sirex Noctilio Larvae, Emerging from Activated Eggs, on a Modified Artificial Diet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sartain, C.V.; Wolfner, M.F. Calcium and egg activation in Drosophila. Cell Calcium 2012, 53, 10–15. [Google Scholar] [CrossRef]
- Yamamoto, D.S.; Hatakeyama, M.; Matsuoka, H. Artificial activation of mature unfertilized eggs in the malaria vector mosquito, Anopheles stephensi (Diptera, Culicidae). J. Exp. Biol. 2013, 216, 2960–2966. [Google Scholar] [CrossRef]
- Horner, V.L.; Wolfner, M.F. Transitioning from egg to embryo: Triggers and mechanisms of egg activation. Dev. Dyn. 2008, 237, 527–544. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Sobajima, H.; Satoh, T.; Obara, Y. Activation in vitro of unfertilized egg development in honeybee queens. Naturwissenschaften 1997, 84, 74–76. [Google Scholar] [CrossRef]
- Sperling, A.L.; Glover, D.M. Parthenogenesis in dipterans: A genetic perspective. Proc. R. Soc. B Biol. Sci. 2023, 290, 20230261. [Google Scholar] [CrossRef] [PubMed]
- Suomalainen, E. Significance of parthenogenesis in the evolution of insects. Annu. Rev. Entomol. 1962, 7, 349–366. [Google Scholar] [CrossRef]
- Rabeling, C.; Kronauer, D.J. Thelytokous parthenogenesis in eusocial Hymenoptera. Annu. Rev. Entomol. 2013, 58, 273–292. [Google Scholar] [CrossRef]
- Miura, K.; Tagami, Y. Comparison of life history characters of arrhenotokous and Wolbachia-associated thelytokous Trichogramma kaykai Pinto and Stouthamer (Hymenoptera: Trichogrammatidae). Ann. Entomol. Soc. Am. 2004, 97, 765–769. [Google Scholar] [CrossRef]
- Pearcy, M.; Hardy, O.; Aron, S. Thelytokous parthenogenesis and its consequences on inbreeding in an ant. Heredity 2006, 96, 377–382. [Google Scholar] [CrossRef]
- Went, D.F. Egg activation and parthenogenetic reproduction in insects. Biol. Rev. 1982, 57, 319–344. [Google Scholar] [CrossRef]
- Pijnacker, L.P.; Ferwerda, M.A. Experiments on blocking and unblocking of first meiotic metaphase in eggs of the parthenogenetic stick insect Carausius morosus Br. (Phasmida, Insecta). J. Embryol. Exp. Morphol. 1976, 36, 383–394. [Google Scholar] [CrossRef]
- Mahowald, A.P.; Goralski, T.J.; Caulton, J.H. In vitro activation of Drosophila eggs. Dev. Biol. 1983, 98, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Sawa, M.; Oishi, K. Studies on the sawfly, Athalia rosae (Insecta, Hymenoptera, Teethredinidae). II. Experimental activation of mature unfertilized eggs. Zool. Sci. 1989, 6, 549–556. [Google Scholar]
- York-Andersen, A.H.; Wood, B.W.; Wilby, E.L.; Berry, A.S.; Weil, T.T. Osmolarity-regulated swelling initiates egg activation in Drosophila. Open Biol. 2021, 11, 210067. [Google Scholar] [CrossRef]
- Went, D.F.; Krause, G. Alteration of egg architecture and egg activation in an endoparasitic hymenopteran as a result of natural or imitated oviposition. Wilhelm Roux Arch. Für Entwicklungsmechanik Der Org. 1974, 175, 173–184. [Google Scholar] [CrossRef]
- Went, D.F.; Krause, G. Normal development of mechanically activated, unlaid eggs of an endoparasitic hymenopteran. Nature 1973, 244, 454–455. [Google Scholar] [CrossRef]
- King, P.E.; Rafai, J. A possible mechanism for initiating the parthenogenetic development of eggs in a parasitoid hymenopteran, Nasonia vitripennis (Walker) (Pteromalidae). Entomologist 1973, 106, 118–120. [Google Scholar]
- Sasaki, K.; Obara, Y. Egg activation and timing of sperm acceptance by an egg in honeybees (Apis mellifera L.). Insectes Sociaux 2002, 49, 234–240. [Google Scholar] [CrossRef]
- Vinson, S.B.; Jang, H.-S. Activation of Campoletis sonorensis (Hymenoptera: Ichneumonidae) eggs by artificial means. Ann. Entomol. Soc. Am. 1987, 80, 486–489. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Addison, M.F.; Terblanche, J.S. Mass-rearing of insects for pest management: Challenges, synergies and advances from evolutionary physiology. Crop Prot. 2012, 38, 87–94. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Van Eynde, B.; Yu, N.; Ma, S.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef]
- Pacheco, I.D.; Walling, L.L.; Atkinson, P.W. Gene editing and genetic control of hemipteran pests: Progress, challenges and perspectives. Front. Bioeng. Biotechnol. 2022, 10, 900785. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Aumann, R.A.; Häcker, I.; Schetelig, M.F. CRISPR-based genetic control strategies for insect pests. J. Integr. Agric. 2023, 22, 651–668. [Google Scholar] [CrossRef]
- Espinosa del Alba, L.; Petschenka, G. A simple artificial diet for feeding and sequestration assays for the milkweed bugs Oncopeltus fasciatus and Spilostethus saxatilis. Entomol. Exp. Appl. 2023, 171, 658–667. [Google Scholar] [CrossRef]
- Ajó Fernández, A.A.F.; Martínez, A.S.; Villacide, J.M.; Corley, J.C. Behavioural response of the woodwasp Sirex noctilio to volatile emissions of its fungal symbiont. J. Appl. Entomol. 2015, 139, 654–659. [Google Scholar] [CrossRef]
- Ciesla, W.M. European woodwasp: A potential threat to North America’s conifer forests. J. For. 2003, 101, 18–23. [Google Scholar]
- Bedding, R.A. Controlling the pine-killing woodwasp, Sirex noctilio, with nematodes. In Use of Microbes for Control and Eradication of Invasive Arthropods; Hajek, A.E., Glare, T.R., O’Callaghan, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 213–235. [Google Scholar]
- Hoebeke, E.; Haugen, D.A.; Haack, R. Sirex noctilio: Discovery of a palearctic siricid woodwasp in New York. Newsl. Mich. Entomol. Soc. 2005, 50, 24–25. [Google Scholar]
- Coutts, M.P. The mechanism of pathogenicity of Sirex noctilio on Pinus radiata. I. Effects of the symbiotic fungus Amylostereum sp. (Thelophoraceae). Aust. J. Biol. Sci. 1969, 22, 915–924. [Google Scholar] [CrossRef]
- Tribe, G.D. A wasp counterattack to save pine trees. Village Life 2006, 18, 38–40. [Google Scholar]
- Thompson, B.M.; Bodart, J.; McEwen, C.; Gruner, D.S. Adaptations for symbiont-mediated external digestion in Sirex noctilio (Hymenoptera: Siricidae). Ann. Entomol. Soc. Am. 2014, 107, 453–460. [Google Scholar] [CrossRef]
- Coutts, M.P. The mechanism of pathogenicity of Sirex noctilio on Pinus radiata. II. Effects of S. noctilio mucus. Aust. J. Biol. Sci. 1969, 22, 1153–1161. [Google Scholar] [CrossRef]
- Madden, J.L. Egg and larval development in the woodwasp, Sirex noctilio F. Aust. J. Zool. 1981, 29, 493–506. [Google Scholar] [CrossRef]
- Madden, J.L.; Coutts, M.P. The role of fungi in the biology and ecology of woodwasps (Hymenoptera: Siricidae). In Insect-Fungus Symbiosis; Batra, L.R., Ed.; Allanheld, Osmun & Co.: Totowa, NJ, USA, 1979; pp. 165–174. [Google Scholar]
- Hurley, B.P.; Slippers, B.; Wingfield, M.J. A comparison of control results for the alien invasive woodwasp, Sirex noctilio, in the southern hemisphere. Agric. For. Entomol. 2007, 9, 159–171. [Google Scholar] [CrossRef]
- Ryan, K.; Hurley, B.P. Life history and biology of Sirex noctilio. In The Sirex Woodwasp and Its Fungal Symbiont; Slippers, B., De Groot, P., Wingfield, M.J., Eds.; Springer: Dordrecht, The Netherland, 2012; pp. 15–30. [Google Scholar]
- Coutts, M.P.; Dolezal, J.E. Emplacement of fungal spores by the woodwasp, Sirex noctilio, during oviposition. For. Sci. 1969, 15, 412–416. [Google Scholar]
- Corley, J.C.; Lantschner, M.V.; Martínez, A.S.; Fischbein, D.; Villacide, J.M. Management of Sirex noctilio populations in exotic pine plantations: Critical issues explaining invasion success and damage levels in South America. J. Pest Sci. 2018, 92, 131–142. [Google Scholar] [CrossRef]
- Slippers, B.; Hurley, B.P.; Wingfield, M.J. Sirex Woodwasp: A model for evolving management paradigms of invasive forest pests. Annu. Rev. Entomol. 2015, 60, 601–619. [Google Scholar] [CrossRef]
- Kraus, S.; Monchanin, C.; Gomez-Moracho, T.; Lihoreau, M. Insect diet. In Encyclopedia of animal cognition and behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–9. [Google Scholar]
- Hull-Sanders, H.; Pepper, E.; Davis, K.; Trotter, R.T. Description of an establishment event by the invasive Asian longhorned beetle (Anoplophora glabripennis) in a suburban landscape in the northeastern United States. PLoS ONE. 2017, 12, e0181655. [Google Scholar] [CrossRef]
- Hajek, A.E.; Haavik, L.J.; Stephen, F.M. Biology and Ecology of Sirex noctilio in North America; USDA, Forest Service, U.S. Department of Argiculture, Forest Health Assessment and Applied Sciences Team: Morgantown, WV, USA, 2021. [Google Scholar]
- Zhao, J.; Ogura, N.; Isono, M. Artificial rearing of Anoplophora glabripennis. Proj. For. Prot. Inst. Ningxia. 1998, 5, 1–6. [Google Scholar]
- Dubois, T.; Hajek, A.E.; Smith, S. Methods for rearing the Asian longhorned beetle (Coleoptera: Cerambycidae) on artificial diet. Ann. Entomol. Soc. Am. 2002, 95, 223–230. [Google Scholar] [CrossRef]
- Keena, M.A. Pourable artificial diet for rearing Anoplophora glabripennis (Coleoptera: Cerambycidae) and methods to optimize larval survival and synchronize development. Ann. Entomol. Soc. Am. 2005, 98, 536–547. [Google Scholar] [CrossRef]
- Mason, C.J.; Scully, E.D.; Geib, S.M.; Hoover, K. Contrasting diets reveal metabolic plasticity in the tree-killing beetle, Anoplophora glabripennis (Cerambycidae: Lamiinae). Sci. Rep. 2016, 6, srep33813. [Google Scholar] [CrossRef]
- Favaro, R.; Lupi, D.; Jucker, C.; Cappellozza, S.; Faccoli, M. An artificial diet for rearing three exotic longhorn beetles invasive to Europe. Bull. Insectology 2017, 70, 91–99. [Google Scholar]
- Kaneuchi, T.; Sartain, C.V.; Takeo, S.; Horner, V.L.; Buehner, N.A.; Aigaki, T.; Wolfner, M.F. Calcium waves occur as Drosophila oocytes activate. Proc. Natl. Acad. Sci. USA. 2015, 112, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Boitano, S.; Sanderson, M.J.; Dirksen, E.R. A role for Ca2+-conducting ion channels in mechanically-induced signal transduction of airway epithelial cells. J. Cell Sci. 1994, 107, 3037–3044. [Google Scholar] [CrossRef]
- Davis, M.J.; Meininger, G.A.; Zawieja, D.C. Stretch-induced increases in intracellular calcium of isolated vascular smooth muscle cells. Am. J. Physiol. Circ. Physiol. 1992, 263, H1292–H1299. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, K.; Dechev, N.; Burke, R.D.; Park, E.J. Development of an autonomous biological cell manipulator with single-cell electroporation and visual servoing capabilities. IEEE Trans. Biomed. Eng. 2009, 56, 2064–2074. [Google Scholar] [CrossRef]
- Landman, W.A.; Malherbe, J.; Engelbrecht, F. South Africa’s present-day climate. In South African Risk and Vulnerability Atlas: Understanding the Social & Environmental Implications of Global Change; Mambo, J., Faccer, K., Eds.; AFRICAN SUN MeDIA: Stellenbosch, South Africa, 2017; pp. 7–12. [Google Scholar]
- Yousuf, F.; Carnegie, A.J.; Bedding, R.A.; Bashford, R.; Nicol, H.I.; Gurr, G.M. Effect of temperature on woodwasp (Sirex noctilio F.) development and parasitism by the entomopathogenic nematode, Deladenus siricidicola. Biol. Control. 2014, 79, 67–74. [Google Scholar] [CrossRef]
- Fischer, K.; Bot, A.N.M.; Zwaan, B.J.; Brakefield, P.M. Genetic and environmental sources of egg size variation in the butterfly Bicyclus anynana. Heredity 2004, 92, 163–169. [Google Scholar] [CrossRef]
- Gibbs, M.; Breuker, C.J.; VAN Dyck, H. Flight during oviposition reduces maternal egg provisioning and influences offspring development in Pararge aegeria (L.). Physiol. Entomol. 2010, 35, 29–39. [Google Scholar] [CrossRef]
- Queffelec, J.; Wooding, A.L.; Greeff, J.M.; Garnas, J.R.; Hurley, B.P.; Wingfield, M.J.; Slippers, B. Mechanisms that influence sex ratio variation in the invasive hymenopteran Sirex noctilio in South Africa. Ecol. Evol. 2019, 9, 7966–7973. [Google Scholar] [CrossRef]
- Markert, J.A.; Champlin, D.M.; Gutjahr-Gobell, R.; Grear, J.S.; Kuhn, A.; McGreevy, T.J.; Roth, A.; Bagley, M.J.; Nacci, D.E. Population genetic diversity and fitness in multiple environments. BMC Evol. Biol. 2010, 10, 205. [Google Scholar] [CrossRef]
- Hongwane, P.; Mitchell, G.; Kanzler, A.; Verryn, S.; Lopez, J.; Chirwa, P. Alternative pine hybrids and species to Pinus patula and P. radiata in South Africa and Swaziland. South. For. J. For. Sci. 2018, 80, 301–310. [Google Scholar] [CrossRef]
- Luo, D.; Lai, M.; Xu, C.; Shi, H.; Liu, X. Life history traits in a capital breeding pine caterpillar: Effect of host species and needle age. BMC Ecol. 2018, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Madden, J.L. Physiological aspects of host tree favourability for the woodwasp, Sirex noctilio F. Proc. Ecol. Soc. Aust. 1968, 3, 147–149. [Google Scholar]
- Wang, P.; Lu, P.-F.; Zheng, X.-L.; Chen, L.-Z.; Lei, C.-L.; Wang, X.-P. New artificial diet for continuous rearing of the bean pod borer, Maruca vitrata. J. Insect Sci. 2013, 13, 121. [Google Scholar] [CrossRef]
- Snodgrass, R.E. Insect metamorphosis. Smithson. Misc. Collect. 1984, 122, 1–124. [Google Scholar]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Wang, M.; Wang, L.; Liu, X.; Gao, C.; Ren, L.; Luo, Y. Gut structure and microbial communities in Sirex noctilio (Hymenoptera: Siricidae) and their predicted contribution to larval nutrition. Front. Microbiol. 2021, 12, 641141. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Amount |
|---|---|
| Distilled water | 257.81 mL |
| Agar | 8.20 g |
| Group 1 | |
| Raw wheat germ | 19.92 g |
| Torula yeast powder | 10.55 g |
| Group 2 | |
| Wesson salt mixture | 1.05 g |
| Sorbic acid | 0.73 g |
| Methyl paraben | 0.73 g |
| Sucrose | 5.86 g |
| Casein from bovine milk | 3.52 g |
| Sodium propionate | 0.45 g |
| Group 3 | |
| Cholesterol | 0.35 g |
| Autoclaved wheatgerm oil | 1.64 mL |
| Group 4 | |
| Choline chloride | 0.09 g |
| Vanderzant vitamin mixture for insects | 1.55 g |
| Vitamin A beadlets | 0.12 g |
| Group 5 | |
| Alpha cellulose | 45.70 g |
| Group 6 | |
| Streptomycin sulfate salt | 0.3 g |
| SABAX pour water | 2 ml |
| Group 7 | |
| Autoclaved pine sawdust | 50 g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Merwe, E.; Slippers, B.; Dittrich-Schröder, G. Mechanical Egg Activation and Rearing of First Instar Larvae of Sirex noctilio (Hymenoptera: Siricidae). Insects 2023, 14, 931. https://doi.org/10.3390/insects14120931
van der Merwe E, Slippers B, Dittrich-Schröder G. Mechanical Egg Activation and Rearing of First Instar Larvae of Sirex noctilio (Hymenoptera: Siricidae). Insects. 2023; 14(12):931. https://doi.org/10.3390/insects14120931
Chicago/Turabian Stylevan der Merwe, Elmarie, Bernard Slippers, and Gudrun Dittrich-Schröder. 2023. "Mechanical Egg Activation and Rearing of First Instar Larvae of Sirex noctilio (Hymenoptera: Siricidae)" Insects 14, no. 12: 931. https://doi.org/10.3390/insects14120931
APA Stylevan der Merwe, E., Slippers, B., & Dittrich-Schröder, G. (2023). Mechanical Egg Activation and Rearing of First Instar Larvae of Sirex noctilio (Hymenoptera: Siricidae). Insects, 14(12), 931. https://doi.org/10.3390/insects14120931