
Invasion Genetics of the Horse-Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A Case of Successful Involvement of Citizen Science in Studying an Alien Insect Pest

 ,
,  ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Volunteers’ Involvement
2.3. Insect Specimen Sampling
2.4. Molecular Genetic Analysis
2.5. Trophic Associations
3. Results
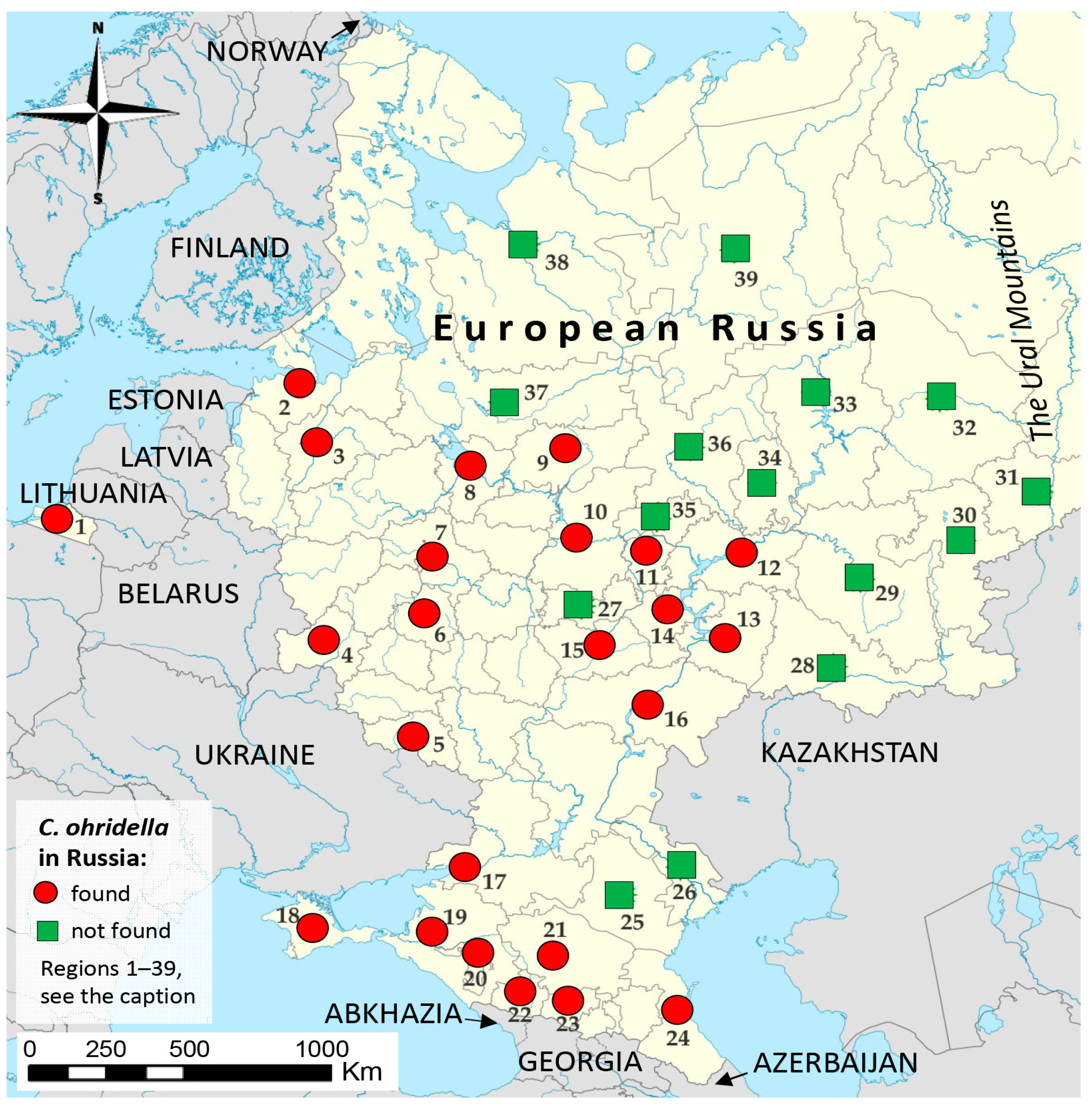
3.1. Sampling Efforts and the Detection of C. ohridella across the European Part of Russia
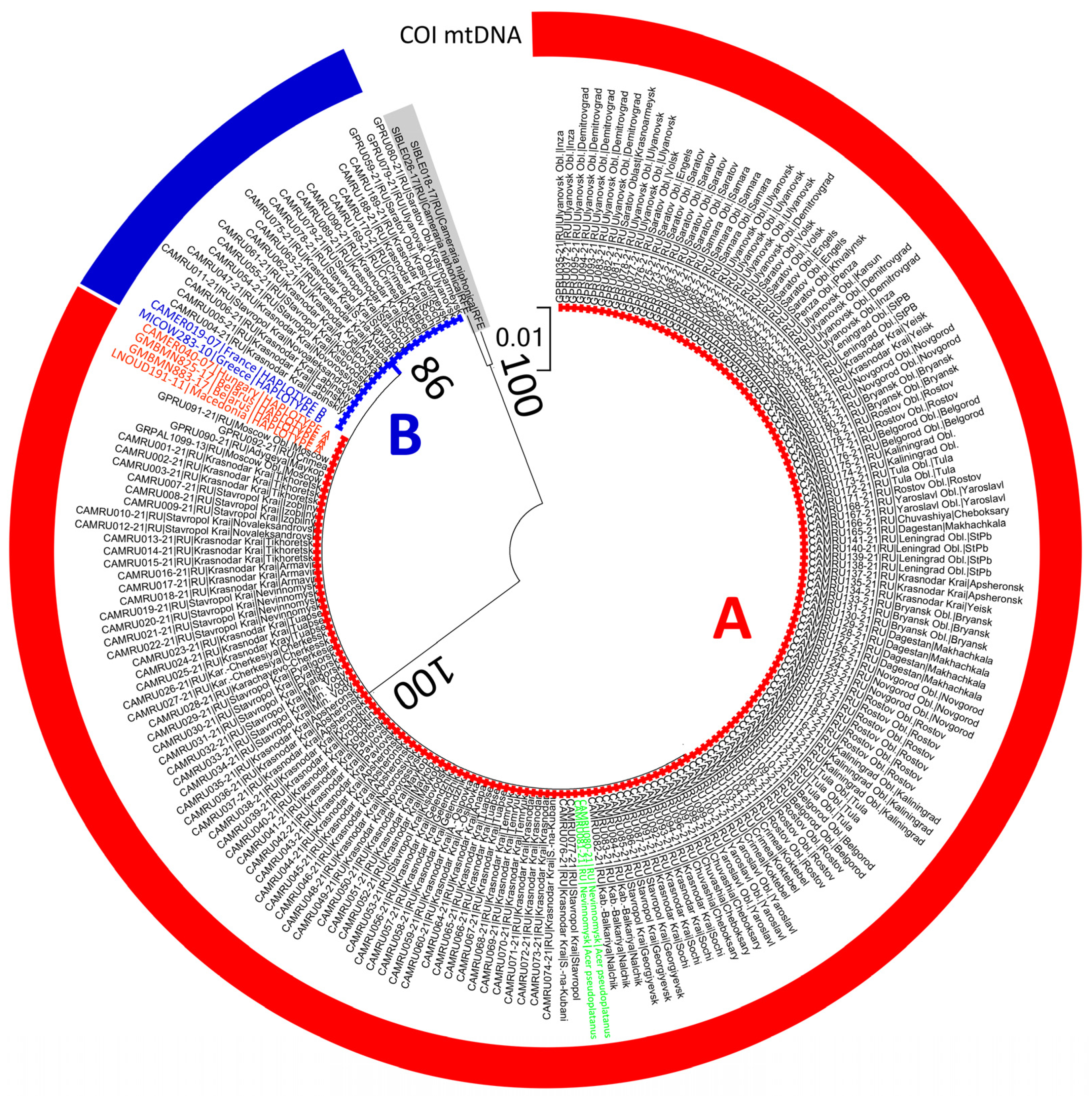
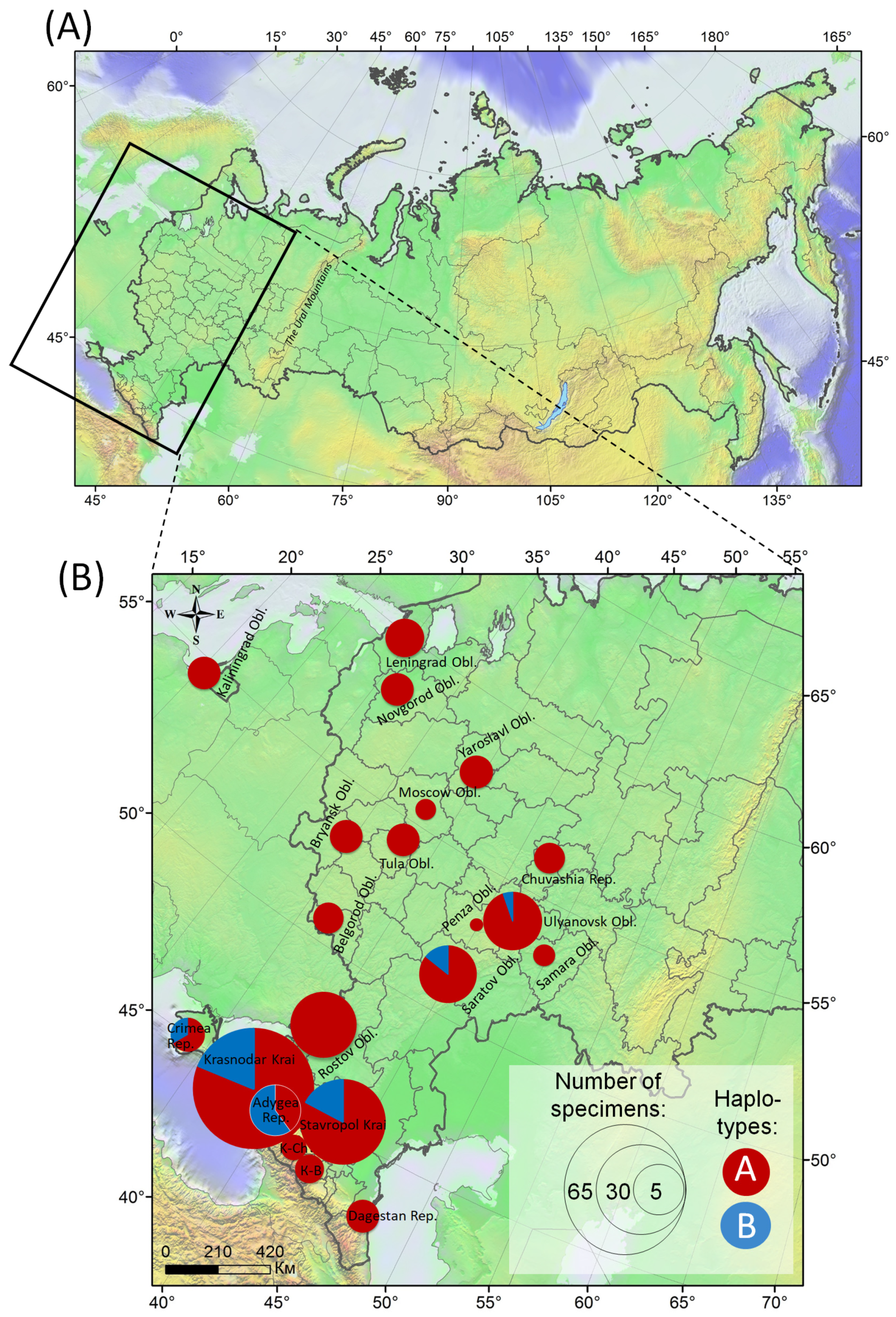
3.2. Genetic Variability of C. ohridella in the European Part of Russia
3.3. Trophic Associations of C. ohridella with Sapindaceae in Southern Russia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowry, E.; Rollinson, E.J.; Laybourn, A.J.; Scott, T.E.; Aiello-Lammens, M.E.; Gray, S.M.; Mickley, J.; Gurevitch, J. Biological invasions: A field synopsis, systematic review, and database of the literature. Ecol. Evol. 2012, 3, 182–196. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Elton, C.S.; Simberloff, D.; Ricciardi, A. The Ecology of Invasions by Animals and Plants, 2nd ed.; Springer: Cham, Switzerland, 2020; 181p. [Google Scholar]
- Kenis, M.; Auger-Rozenberg, M.-A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.W.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Liu, C.; Diagne, C.; Angulo, E.; Banerjee, A.-K.; Chen, Y.; Cuthbert, R.N.; Haubrock, P.J.; Kirichenko, N.; Pattison, Z.; Watari, Y.; et al. Economic costs of biological invasions in Asia. NeoBiota 2021, 67, 53–78. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. Available online: https://neobiota.pensoft.net/article/58196/ (accessed on 17 January 2023). [CrossRef]
- Kirichenko, N.; Haubrock, P.J.; Cuthbert, R.N.; Akulov, E.; Karimova, E.; Shneyder, Y.; Liu, C.; Angulo, E.; Diagne, C.; Courchamp, F. Economic costs of biological invasions in terrestrial ecosystems in Russia. NeoBiota 2021, 67, 103–130. [Google Scholar] [CrossRef]
- Dang, Y.; Wei, K.; Wang, X.; Duan, J.J.; Jennings, D.E.; Poland, T.M. Introduced plants induce outbreaks of a native pest and facilitate invasion in the plants’ native range: Evidence from the emerald ash borer. J. Ecol. 2022, 110, 593–604. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2018, 92, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Unwelcome exchange: International trade as a direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, 666–679. [Google Scholar] [CrossRef]
- Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive insect pests of forests and urban trees in Russia: Origin, pathways, damage, and management. Forests 2022, 13, 521. [Google Scholar] [CrossRef]
- Hill, M.P.; Clusella-Trullas, S.; Terblanche, J.S.; Richardson, D.M. Drivers, impacts, mechanisms and adaptation in insect invasions. Biol. Invasions 2016, 18, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.E.; Callaham, M.A.; Stewart, J.E.; Warren, S.D. Invasive species response to natural and anthropogenic disturbance. In Invasive Species in Forests and Rangelands of the United States; Poland, T.M., Patel-Weynand, T., Finch, D.M., Miniat, C.F., Hayes, D.C., Lopez, V.M., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. Effect of climate change on introduced and native agricultural invasive insect pests in Europe. Insects 2021, 12, 985. [Google Scholar] [CrossRef]
- Moricca, S.; Bracalini, M.; Croci, F.; Corsinovi, S.; Tiberi, R.; Ragazzi, A.; Panzavolta, T. Biotic factors affecting ecosystem services in urban and peri-urban forests in Italy: The role of introduced and impending pathogens and pests. Forests 2018, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Solomou, A.D.; Topalidou, E.T.; Germani, R.; Argiri, A.; Karetsos, G. Importance, Utilization and health of urban forests: A review. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 47, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Maslyakov, V.Y.; Izhevsky, S.S. Invasions of Herbivorous Insects in the European Part of Russia; IGRAN: Moscow, Russia, 2011. (In Russian) [Google Scholar]
- Karpun, N.N. The Structure of Complexes of Harmful Organisms of Woody Plants in the Humid Subtropics of Russia and the Biological Justification of Protection Measures: Diss …; Dr. Biol. Sciences: Sochi, Russia, 2018. (In Russian) [Google Scholar]
- Shchurov, V.I.; Zamotailov, A.S. The first finds of the ash emerald borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in the Krasnodar Territory. In Proceedings of the Biological Diversity of the Caucasus and Southern Russia, XXIV International Scientific Conference, Magnus, Russia, 17–20 November 2022; pp. 558–565. (In Russian). [Google Scholar]
- Šefrová, H. Invasions of Lithocolletinae species in Europe—Causes, kinds, limits and ecological impact (Lepidoptera, Gracillariidae). Ekol. Bratisl. 2003, 22, 132–142. [Google Scholar]
- Kirichenko, N.; Augustin, S.; Kenis, M. Invasive leafminers on woody plants: A global review of pathways, impact and management. J. Pest Sci. 2019, 92, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Csóka, G. Recent invasions of five species of leafmining Lepidoptera in Hungary. In Proceedings of the Integrated Management and Dynamics of Forest Defoliating Insects, Victoria, BC, Canada, 15–19 August 1999; Liebhold, A.M., McManus, M.L., Otvos, I.S., Fosbroke, S.L.C., Eds.; U.S. Department of Agriculture, Forest Service, Northeastern Research Station: Newtown Square, PA, USA, 2001. NE-277. pp. 31–36. [Google Scholar]
- Csóka, G. Leaf Mines and Leaf Miners; Agroinform: Budapest, Hungary, 2003. [Google Scholar]
- Valade, R.; Kenis, M.; Hernandez-Lopez, A.; Augustin, S.; Mari Mena, N.; Magnoux, E.; Rougerie, R.; Lakatos, F.; Roques, A.; Lopez-Vaamonde, C. Mitochondrial and microsatellite DNA markers reveal a Balkan origin for the highly invasive horse-chestnut leaf miner Cameraria ohridella (Lepidoptera, Gracillariidae). Mol. Ecol. 2009, 18, 3458–3470. [Google Scholar] [CrossRef]
- Lees, D.C.; Lack, H.W.; Rougerie, R.; Hernandez-Lopez, A.; Raus, T.; Avtzis, N.; Augustin, S.; Lopez-Vaamonde, C. Tracking origins of invasive herbivores using herbaria and archival DNA: The case of the horse-chestnut leafminer. Front. Ecol. Environ. 2011, 9, 322–328. [Google Scholar] [CrossRef]
- Lopez-Vaamonde, C.; Kirichenko, N.; Cama, A.; Doorenweerd, C.; Godfray, H.C.J.; Guiguet, A.; Gomboc, S.; Huemer, P.; Landry, J.-F.; Laštuvka, A.; et al. Evaluating DNA Barcoding for Species Identification and Discovery in European Gracillariid Moths. Front. Ecol. Evol. 2021, 9, 626752. [Google Scholar] [CrossRef]
- Tomiczek, C.; Krehan, H. The horse chestnut leafmining moth (Cameraria ohridella): A new pest in Central Europe. Arboric. J. 1998, 24, 144–148. [Google Scholar]
- Rakov, A.G. Horse Chestnut Leaf Miner and Other Invasive Dendrophilous Phyllophagous under the Conditions of Their Range Formation in the European Part of Russia, Diss. …; Cand. biol. Sciences: Moscow, Russia, 2015. (In Russian) [Google Scholar]
- Anikin, V.V. Present day bio-invasions in the Volga-Ural Region: From the South to the North or from the East to the West? Cameraria ohridella (Lepidoptera: Gracillariidae) in the Lower and Middle Volga. Zootaxa 2019, 4624, 583–588. [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Orlinski, A.D. New insect pests of forest plantations. Plant Prot. Quar. 2004, 4, 33. (In Russian) [Google Scholar]
- Golosova, M.A.; Gninenko, Y.I. The appearance of the horse-chestnut leafminer on horse chestnut in Moscow. For. Bull. 2006, 2, 43–46. Available online: https://www.elibrary.ru/download/elibrary_9273118_59679849.pdf (accessed on 4 December 2022). (In Russian).
- Zhuravleva, E.N. The first appearance of the horse chestnut leaf miner Cameraria ohridella (Lepidoptera: Gracillariidae) on the horse chestnut in the territory of Bolshoy Sochi. In The Kataev Memorial Readings—VIII. Pests and Diseases of Woody Plants in Russia, Proceedings of an International Conference, St. Petersburg, Russia, 18–20 November 2014; Musolin, D.L., Selikhovkin, A.V., Eds.; St. Petersburg State Forest Technical University: St. Petersburg, Russia, 2014; p. 32. (In Russian) [Google Scholar]
- Shutko, A.P.; Tuturzhans, L.V. Adventive insect pests of wood plants of the Stavropol city. Uspekhi Sovrem. Estestvozn. 2018, 2, 184–189. (In Russian) [Google Scholar]
- Encarnação, J.; Teodósio, M.A.; Morais, P. Citizen science and biological invasions: A review. Front. Environ. Sci. 2021, 8, 602980. [Google Scholar] [CrossRef]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- De Groot, M.; Pocock, M.J.O.; Bonte, J.; Fernandez-Conradi, P.; Valdés-Correcher, E. Citizen science and monitoring forest pests: A beneficial alliance? Curr. For. Rep. 2023, 9, 15–32. [Google Scholar] [CrossRef]
- Pocock, M.J.O.; Evans, D.M. The success of the horse chestnut leafminer, Cameraria ohridella, in the UK revealed with hypothesis led citizen science. PLoS ONE 2014, 9, e86226. [Google Scholar] [CrossRef]
- Brown, N.; Pérez-Sierra, A.; Crow, P.; Parnell, S. The role of passive surveillance and citizen science in plant health. CABI Agric. Biosci. 2020, 1, 17. [Google Scholar] [CrossRef]
- Deschka, G.; Dimić, N. Cameraria ohridella n. sp. aus Mazedonien, Jugoslawien (Lepidoptera, Lithocelletidae). Acta Entomol. Jugosl. 1986, 22, 11–23. [Google Scholar]
- De Waard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: Analytical methods. In Methods in Molecular Biology: Environmental Genetics; Cristopher, M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2008. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 9.3; Environmental Systems Research Institute: Redlands, CA, USA, 2008; Available online: http://www.esri.com/software/arcgis/eval-help/arcgis-93 (accessed on 5 December 2022).
- De Groot, M.; O’Hanlon, R.; Bullas-Appleton, E.; Csóka, G.; Csiszár, Á.; Faccoli, M.; Gervasini, E.; Kirichenko, N.; Korda, M.; Marinšek, A.; et al. Challenges and solutions in early detection, rapid response and communication about potential invasive alien species in forests. Manag. Biol. Invasions 2020, 11, 637–660. [Google Scholar] [CrossRef]
- Larson, E.R.; Graham, B.M.; Achury, R.; Coon, J.J.; Daniels, M.K.; Gambrell, D.K.; Jonasen, K.L.; King, G.D.; LaRacuente, N.; Perrin-Stowe, Y.I.N.; et al. From eDNA to citizen science: Emerging tools for the early detection of invasive species. Front. Ecol. Environ. 2020, 18, 194–202. [Google Scholar] [CrossRef]
- Golani, D.; Azzurro, E.; Corsini-Foka, M.; Falautano, M.; Andaloro, F.; Bernardi, G. Genetic bottlenecks and successful biological invasions: The case of a recent Lessepsian migrant. Biol. Lett. 2007, 3, 541–545. [Google Scholar] [CrossRef]
- Puillandre, N.; Dupas, S.; Dangles, O.; Zeddam, J.-L.; Capdevielle-Dulac, C.; Barbin, K.; Torres-Leguizamon, M.; Silvain, J.-F. Genetic bottleneck in invasive species: The potato tuber moth adds to the list. Biol. Invasions 2008, 10, 319–333. [Google Scholar] [CrossRef]
- Akimov, I.A.; Zerova, M.D.; Gershenzon, Z.S.; Narolsky, N.B.; Kokhanets, A.M.; Sviridov, S.V. The first report on the appearance of the horse-chestnut leaf miner Cameraria ohridella (Lepidoptera, Gracillariidae) on the horse chestnut Aesculus hippocastanum (Hippocastanaceae) in Ukraine. Vestn. Zool. 2003, 37, 41–52. (In Russian) [Google Scholar]
- Ivinskis, P.; Rimšaitė, J. The horse-chestnut leafminer (Cameraria Ohridella Deschka & Dimic 1986) Lepidoptera, Gracillariidae in Lithuania. Acta Zool. Litu. 2006, 16, 323–327. [Google Scholar] [CrossRef]
- Metla, Z.; Voitkāne, S.; Sešķēna, R.; Petrova, V.; Jankevica, L. Presence of entomopathogenic fungi and bacteria in Latvian population of horse-chestnut leaf miner Cameraria ohridella. Acta Biol. Univ. Daugavp. 2013, 13, 69–76. [Google Scholar]
- Augustin, S.; Guichard, S.; Heitland, W.; Freise, J.; Svatos, A.; Gilbert, M. Monitoring and dispersal of the invading Gracillariidae Cameraria ohridella. J. Appl. Entomol. 2009, 133, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.; Grégoire, J.-C.; Freise, J.F.; Heitland, W. Long-distance dispersal and human population density allow the prediction of invasive patterns in the horse chestnut leaf miner Cameraria ohridella. J. Anim. Ecol. 2004, 73, 459–468. [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Orlinski, A.D. New phytophages of tree plantations. Plant Prot. Quar. 2004, 4, 33. (In Russian) [Google Scholar]
- Chenikalova, E.V. The chestnut moth of Cameraria ohridella (Deschka et Dimić, 1986) and boxwood moth Cydalima perspectalis (Walker, 1859) in the tree stands of Stavropol and its environs. In Dendrobiotic Invertebrates and Fungi and Their Role in Forest Ecosystems (The Kataev Memorial Readings XI), Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; St. Petersburg State Forest Technical University: St. Petersburg, Russia, 2020; pp. 347–348. (In Russian) [Google Scholar]
- Martynov, V.V.; Nikulina, T.V.; Shokhin, I.V.; Terskov, E.N. Contributions to the fauna of invasive insects of Ciscaucasia. Field Biol. J. 2020, 2, 99–122. [Google Scholar] [CrossRef]
- Selikhovkin, A.V.; Drenkhan, R.; Mandelshtam, M.Y.; Musolin, D.L. Invasions of insect pests and fungal pathogens of woody plants into the northwestern part of European Russia. Earth Sci. 2020, 65, 263–283. (In Russian) [Google Scholar] [CrossRef]
- Golosova, M.A.; Gninenko, Y.I.; Golosova, E.I. The Horse-Chestnut Leaf Miner Cameraria ohridella Is a Dangerous Quarantine Pest in Urban Landscaping; VPRS MOBB, MGUL, VNIILM: Moscow, Russia, 2008. (In Russian) [Google Scholar]
- Péré, C.; Augustin, S.; Turling, T.C.J.; Kenis, M. The invasive alien leaf miner Cameraria ohridella and the native tree Acer pseudoplatanus: A fatal attraction? Agric. For. Entomol. 2010, 12, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Freise, J.F.; Heitland, W.; Tosevski, I. Parasitism of the horse chestnut leaf miner, Cameraria ohridella Deschka and Dimic (Lep., Gracillariidae), in Serbia and Macedonia. J. Pest Sci. 2002, 75, 152–157. [Google Scholar] [CrossRef]
- Pitkin, B. Cameraria ohridella Deschka & Dimic, 1986 on Acer platanoides. In The Leaf and Steam Mines of British Flies and Other Insects; Pitkin, B., Ellis, W., Plant, C., Edmunds, R., Eds.; 2022. UK. Available online: http://www.leafmines.co.uk/html/Lepidoptera/C.ohridella.htm (accessed on 4 December 2022).
- Acevedo-Rodríguez, P.; Wurdack, K.J.; Ferrucci, M.S.; Johnson, G.; Dias, P.; Coelho, R.G.; Somner, G.V.; Steinmann, V.W.; Zimmer, E.A.; Strong, M.T. Generic relationships and classification of tribe Paullinieae (Sapindaceae) with a new concept of supertribe Paulliniodae. Syst. Bot. 2017, 42, 96–114. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Locality (City, Village) | Damage Level * | |||
|---|---|---|---|---|---|
| Low 1–25% | Moderate 26–50% | High 51–75% | Severe >75% | ||
| Krasnodar Krai | |||||
| 1 | Anapa | ||||
| 2 | Apsheronsk | ||||
| 3 | Armavir | ||||
| 4 | Arkhipo-Osipovka | ||||
| 5 | Gelendzhik | ||||
| 6 | Krasnodar | ||||
| 7 | Kropotkin | ||||
| 8 | Labinsk | ||||
| 9 | Novomikhailovsky | ||||
| 10 | Novorossiysk | ||||
| 11 | Slavyansk-na-Kubani | ||||
| 12 | Sochi, Adler district | ||||
| 13 | Sochi, Central district | ||||
| 14 | Stanitsa Pavlovskaya | ||||
| 15 | Temryuk | ||||
| 16 | Tikhoretsk | ||||
| 17 | Tuapse | ||||
| 18 | Khadyzhensk | ||||
| 19 | Khopersky | ||||
| The Republic of Adygea | |||||
| 20 | Maykop | ||||
| Stavropol Krai | |||||
| 21 | Izobilny | ||||
| 22 | Novoaleksandrovsk | ||||
| 23 | Nevinnomyssk | ||||
| 24 | Pyatigorsk | ||||
| 25 | Mineralnye Vody | ||||
| 26 | Kislovodsk | ||||
| 27 | Stavropol | ||||
| 28 | Georgievsk | ||||
| The Republic of Kabardino-Balkaria | |||||
| 29 | Nalchik | ||||
| The Republic of Karachay-Cherkessia | |||||
| 30 | Cherkessk | ||||
| TOTAL number of localities by the damage level | 2 (7 %) | 4 (13 %) | 10 (33 %) | 14 (47 %) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirichenko, N.I.; Karpun, N.N.; Zhuravleva, E.N.; Shoshina, E.I.; Anikin, V.V.; Musolin, D.L. Invasion Genetics of the Horse-Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A Case of Successful Involvement of Citizen Science in Studying an Alien Insect Pest. Insects 2023, 14, 117. https://doi.org/10.3390/insects14020117
Kirichenko NI, Karpun NN, Zhuravleva EN, Shoshina EI, Anikin VV, Musolin DL. Invasion Genetics of the Horse-Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A Case of Successful Involvement of Citizen Science in Studying an Alien Insect Pest. Insects. 2023; 14(2):117. https://doi.org/10.3390/insects14020117
Chicago/Turabian StyleKirichenko, Natalia I., Natalia N. Karpun, Elena N. Zhuravleva, Elena I. Shoshina, Vasily V. Anikin, and Dmitrii L. Musolin. 2023. "Invasion Genetics of the Horse-Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A Case of Successful Involvement of Citizen Science in Studying an Alien Insect Pest" Insects 14, no. 2: 117. https://doi.org/10.3390/insects14020117
APA StyleKirichenko, N. I., Karpun, N. N., Zhuravleva, E. N., Shoshina, E. I., Anikin, V. V., & Musolin, D. L. (2023). Invasion Genetics of the Horse-Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A Case of Successful Involvement of Citizen Science in Studying an Alien Insect Pest. Insects, 14(2), 117. https://doi.org/10.3390/insects14020117