
Side Effects of Kaolin and Bunch-Zone Leaf Removal on Generalist Predators in Vineyards



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Vineyards
2.2. Experimental Design
2.3. Samplings
2.4. Statistical Analyses
3. Results
3.1. Spiders and Generalist Predatory Insects in Vineyards
3.1.1. Spiders by the Drop Cloth Method
3.1.2. Predatory Arthropods by Yellow Sticky Traps
3.2. Effect of Kaolin and Bunch-Zone Leaf Removal on Predatory Insects and Spiders
3.2.1. Vineyard A
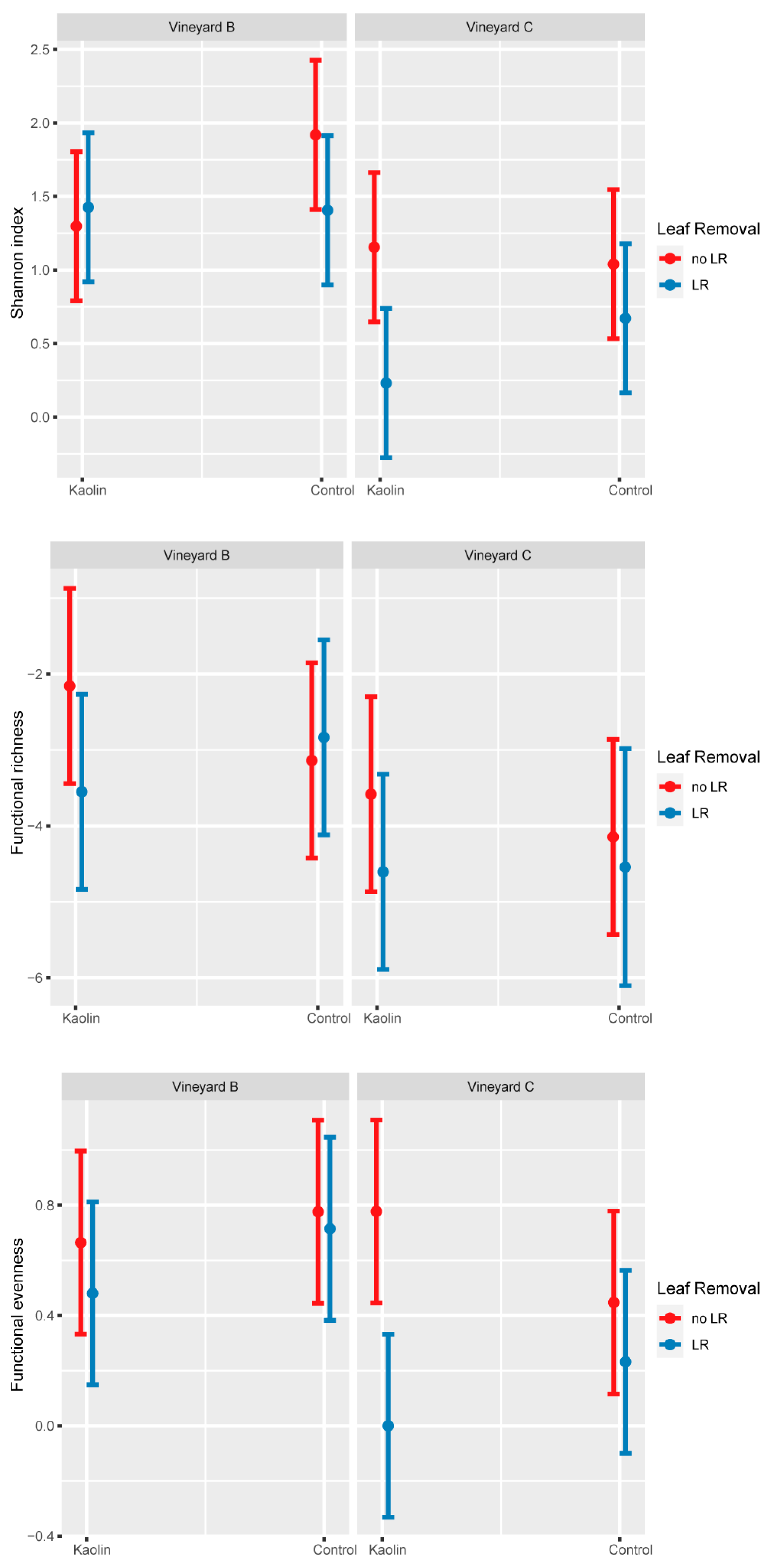
3.2.2. Vineyards B and C
3.2.3. Comparison between the Two Sampling Methods
4. Discussion
4.1. Spiders and Generalist Predatory Insects in Vineyards
4.2. Influence of Kaolin and Bunch-Zone Leaf Removal on Spiders and Generalist Predatory Insects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glenn, D.M.; Puterka, G.; Vanderzwet, T.; Byers, R.E.; Feldhake, C. Hydrophobic particle films: A new paradigm for suppression of arthropod pests and plant diseases. J. Econ. Entomol. 1999, 92, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Salerno, G.; Rebora, M.; Kovalev, A.; Gorb, E.; Gorb, S. Kaolin nano-powder effect on insect attachment ability. J. Pest Sci. 2020, 93, 315–327. [Google Scholar] [CrossRef]
- Vincent, C.; Hallman, G.; Panneton, B.; Fleurat-Lessard, F. Management of agricultural insects with physical control method. Annu. Rev. Entomol. 2003, 48, 261–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unruh, T.R.; Knight, A.L.; Upton, J.; Glenn, D.M.; Puterka, G.J. Particle films for suppression of the codling moth (Lepidoptera: Tortricidae) in apple and pear orchards. J. Econ. Entomol. 2000, 93, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Pascual, S.; Cobos, G.; Seris, E.; González-Núñez, M. Effects of processed kaolin on pests and non-target arthropods in a Spanish olive grove. J. Pest Sci. 2010, 83, 121–133. [Google Scholar] [CrossRef]
- Markό, V.; Blommers, L.H.M.; Bogya, S.; Helsen, H. Kaolin particle films suppress many apple pests, disrupt natural enemies and promote woolly apple aphid. J. Appl. Entomol. 2008, 132, 26–35. [Google Scholar] [CrossRef]
- Lo Verde, G.; Caleca, V.; Lo Verde, V. The use of kaolin to control Ceratitis capitata in organic citrus groves. Bull. Insectol. 2011, 64, 229–273. [Google Scholar]
- Marcotegui, A.; Sánchez-Ramos, I.; Pascual, S.; Fernández, C.E.; Cobos, G.; Armendáriz, I.; Cobo, A.; González-Núñez, M. Kaolin and potassium soap with thyme essential oil to control Monosteira unicostata and other phytophagous arthropods of almond trees in organic orchards. J. Pest Sci. 2015, 88, 753–765. [Google Scholar] [CrossRef]
- Xu, Y.; Dietrich, C.H.; Zhang, Y.-L.; Dmitriev, D.A.; Zhang, L.; Wang, Y.-M.; Lu, S.-H.; Qin, D.-Z. Phylogeny of the tribe Empoascini (Hemiptera: Cicadellidae: Typhlocybinae) based on morphological characteristics, with reclassification of the Empoasca generic group. Syst. Entomol. 2021, 46, 266–286. [Google Scholar] [CrossRef]
- Abedini, R.; Frarazmand, H.; Jebeleh, E.; Sirjani, M. Effect of kaolin clay (WP 95%) on grape leafhopper, Arboridia kermanshah Dlabola (Hem: Cicadellidae) in field condition. Plant Pest Res. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Tacoli, F.; Pavan, F.; Cargnus, E.; Tilatti, E.; Pozzebon, A.; Zandigiacomo, P. Efficacy and mode of action of kaolin in the control of Empoasca vitis and Zygina rhamni (Hemiptera: Cicadellidae) in vineyards. J. Econ. Entomol. 2017, 110, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Tacoli, F.; Mori, N.; Pozzebon, A.; Cargnus, E.; Da Vià, S.; Zandigiacomo, P.; Duso, C.; Pavan, F. Control of Scaphoideus titanus with natural products in organic vineyards. Insects 2017, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirello, P.; Marchesini, E.; Gherardo, P.; Raniero, D.; Rossetto, F.; Pozzebon, A.; Duso, C. The control of the American leafhopper Erasmoneura vulnerata (Fitch) in European vineyards: Impact of synthetic and natural insecticides. Insects 2021, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Sleezer, S.; Johnson, D.T.; Lewis, B.; Goggin, F.; Rothrock, C.; Savin, M. Foliar grape phylloxera, Daktulosphaira vitifoliae (Fitch), seasonal biology, predictive model and management in the Ozarks region of the United States. Acta Hortic. 2011, 904, 151–156. [Google Scholar] [CrossRef]
- Valizadeh, H.; Abbasipour, H.; Farazmand, H.; Askarianzadeh, A. Evaluation of kaolin application on oviposition control of the vine cicada, Psalmocharias alhageos in vineyards (Homoptera: Cicadidae). Entomol. Gen. 2013, 34, 279–286. [Google Scholar] [CrossRef]
- Pease, C.E.; Lόpez-Olguín, J.F.; Pérez-Moreno, I.; Marco-Mancebόn, V. Effects of kaolin on Lobesia botrana (Lepidoptera: Tortricidae) and its compatibility with the natural enemy, Trichogramma cacoeciae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2016, 109, 740–745. [Google Scholar] [CrossRef]
- Tacoli, F.; Cargnus, E.; Kiaeian Moosavi, F.; Zandigiacomo, P.; Pavan, F. Efficacy and mode of action of kaolin and its interaction with bunch-zone leaf removal against Lobesia botrana on grapevines. J. Pest Sci. 2019, 92, 465–475. [Google Scholar] [CrossRef]
- Linder, C.; Staehli, N.; Leumann, M.; Siegfried, W.; Kehrli, P. Physical barriers against Drosophila suzukii in viticulture. IOBC/WPRS Bull. 2018, 139, 35–39. [Google Scholar]
- Isaacs, R.; Mercader, R.J.; Wise, J.C. Activity of conventional and reduced-risk insecticides for protection of grapevines against the rose chafer, Macrodactylus subspinosus (Coleoptera: Scarabaeidae). J. Appl. Entomol. 2004, 128, 371–376. [Google Scholar] [CrossRef]
- Maier, R.M.; Williamson, R.C. Evaluation of kaolin clay as an alternative management tactic for Japanese beetle feeding damage in grape vineyards. J. Hortic. 2016, 3, 184. [Google Scholar] [CrossRef] [Green Version]
- Percival, D.C.; Fisher, K.H.; Sullivan, J.A. Use of fruit zone leaf removal with Vitis vinifera L. cv Riesling grapevines. II. Effects on fruit composition, yield, and occurrence of bunch rot (Botrytis cinerea Pers.). Am. J. Enol. Vitic. 1994, 45, 133–139. [Google Scholar] [CrossRef]
- Pavan, F.; Cargnus, E.; Kiaeianmoosavi, F.; Bigot, G.; Tacoli, F.; Zandigiacomo, P. Bunch-zone leaf removal of grapevines to prevent damage by Lobesia botrana and grey mould. Bull. Insectol. 2016, 69, 107–115. [Google Scholar]
- Kiaeian Moosavi, F.; Cargnus, E.; Pavan, F.; Zandigiacomo, P. Effects of grapevine bunch exposure to sunlight on berry surface temperature and Lobesia botrana (Lepidoptera: Tortricidae) egg laying, hatching and larval settlement. Agric. For. Entomol. 2018, 20, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Hosseinabad, A.; Khadivi, A. Foliar application of kaolin reduces the incidence of sunburn in ‘Thompson seedless’ grapevine. Eur. J. Hortic. Sci. 2019, 84, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Marc, P.; Canard, A.; Ysnel, F. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 1999, 74, 229–273. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [Green Version]
- Nyffeler, M.; Sunderland, K.D. Composition, abundance and pest control potential of spider communities in agroecosystems: A comparison of European and US studies. Agric. Ecosyst. Environ. 2003, 95, 579–612. [Google Scholar] [CrossRef]
- Harwood, J.D.; Obrycki, J.J. The role of alternative prey sustaining predator populations. In Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005; pp. 453–462. [Google Scholar]
- Duso, C.; Girolami, V. Ruolo degli Antocoridi nel controllo del Panonychus ulmi Koch nei vigneti. Boll. Ist. entomol. Univ. studi Bologna 1982, 37, 157–169. [Google Scholar]
- Isaia, M.; Bona, F.; Badino, G. Influence of landscape diversity and agricultural practices on spider assemblage in Italian vineyards of Langa Astigiana (Northwest Italy). Environ. Entomol. 2006, 35, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Thiéry, D.; Louâpre, P.; Muneret, L.; Rusch, A.; Sentenac, G.; Vogelweith, F.; Moreau, J. Biological protection against grape berry moths. A review. Agron. Sustain. Dev. 2018, 38, 15. [Google Scholar] [CrossRef] [Green Version]
- Costello, M.J.; Daane, K.M. Spider and leafhopper (Erythroneura spp.) response to vineyard ground cover. Environ. Entomol. 2003, 32, 1085–1098. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Vineyard proximity to riparian habitat influences Western grape leafhopper (Erythroneura elegantula Osborn) populations. Agric. Ecosyst. Environ. 2015, 211, 43–50. [Google Scholar] [CrossRef]
- Nobre, T.; Meierrose, C. The species composition, within-plant distribution, and possible predatory role of spiders (Araneae) in a vineyard in southern Portugal. In Proceedings of the 18th European Colloquium of Arachnology, High Tatras, Slovakia, 12–17 July 1999; Gajdoš, P., Pekar, S., Eds.; Ekológia (Bratislava), 19, suppl. 3/2000. pp. 193–200. [Google Scholar]
- Hogg, B.N.; Daane, K.M. Ecosystem services in the face of invasion: The persistence of native and non-native spiders in an agricultural landscape. Ecol. Appl. 2011, 21, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Hogg, B.N.; Daane, K.M. The roles of top and intermediate predators in herbivore suppression: Contrasting results from the field and laboratory. Ecol. Entomol. 2014, 39, 149–158. [Google Scholar] [CrossRef]
- Altieri, M.A.; Ponti, L.; Nicholls, C.I. Manipulating vineyard biodiversity for improved insect pest management: Case studies from northern California. Int. J. Biodivers. Sci. Manag. 2005, 1, 191–203. [Google Scholar] [CrossRef]
- Hogg, B.N.; Daane, K.M. The role of dispersal from natural habitat in determining spider abundance and diversity in California vineyards. Agric. Ecosyst. Environ. 2010, 135, 260–267. [Google Scholar] [CrossRef]
- Miles, A.; Wilson, H.; Altieri, M.; Nicholls, C. Habitat diversity at the field and landscape level: Conservation biological control research in California viticulture. In Arthropod Management in Vineyards: Pest, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaac, R., Eds.; Springer Science + Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 159–189. [Google Scholar] [CrossRef]
- Judt, C.; Guzmán, G.; Gómez, J.A.; Cabezas, J.M.; Entrenas, J.A.; Winter, S.; Zaller, J.G.; Paredes, D. Diverging effects of landscape factors and inter-row management on the abundance of beneficial and herbivorous arthropods in Andalusian vineyards (Spain). Insects 2019, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Zanettin, G.; Bullo, A.; Pozzebon, A.; Burgio, G.; Duso, C. Influence of vineyard inter-row groundcover vegetation management on arthropod assemblages in the vineyards of north-eastern Italy. Insects 2021, 12, 349. [Google Scholar] [CrossRef]
- Lombardini, L.; Harris, M.K.; Glenn, D.M. Effects of particle film application on leaf gas exchange, water relations, nut yield, and insect populations in mature pecan trees. HortScience 2005, 40, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Karagounis, C.; Kourdoumbalos, A.K.; Margaritopoulos, J.T.; Nanos, G.D.; Tsitsipis, J.A. Organic farming-compatible insecticides against the aphid Myzus persicae (Sulzer) in peach orchards. J. Appl. Entomol. 2006, 130, 150–154. [Google Scholar] [CrossRef]
- Sackett, T.E.; Buddle, C.M.; Vincent, C. Effects of kaolin on the composition of generalist predator assemblages and parasitism of Choristoneura rosaceana (Lep.; Tortricidae) in apple orchards. J. Appl. Entomol. 2007, 131, 478–485. [Google Scholar] [CrossRef]
- Porcel, M.; Cotes, B.; Campos, M. Biological and behavioral effects of kaolin particle film on larvae and adults of Chrysoperla carnea (Neuroptera: Chrysopidae). Biol. Control. 2011, 59, 98–105. [Google Scholar] [CrossRef]
- Santos, R.L.; dos Santos Neves, R.C.; Colares, F.; Torres, J.B. Parasitoids of boll weevil Anthonomus grandis and resident predators in kaolin-treated cotton. Semin.-Ciênc. Agrár. 2013, 34, 3463–3474. [Google Scholar] [CrossRef]
- Markό, V.; Bogya, S.; Kondorosy, E.; Blommers, L.H.M. Side effects of kaolin particle films on apple orchard bug, beetle and spider communities. Int. J. Pest Manag. 2010, 56, 189–199. [Google Scholar] [CrossRef]
- González-Núñez, M.; Pascual, S.; Cobo, A.; Seris, E.; Cobos, G.; Fernández, C.E.; Sánchez-Ramos, I. Copper and kaolin sprays as tools for controlling the olive fruit fly. Entomol. Gen. 2021, 41, 97–110. [Google Scholar] [CrossRef]
- Moraefi, M.; Seddigh, S.; Hamrahi, A. Influence of concentration and time of kaolin application on agronomic traits, canola aphid (Brevicoryne brassicae L.) and its natural enemies. J. Agroecol. 2021, 12, 723–740. [Google Scholar] [CrossRef]
- Tacoli, F.; Cargnus, E.; Pozzebon, A.; Duso, C.; Tirello, P.; Pavan, F. Side effects of kaolin and bunch-zone leaf removal on predatory mite population (Acari: Phytoseiidae) occurring in vineyards. J. Econ. Entomol. 2019, 112, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth stages of the grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera). Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package, Version 1.0-12; 2014. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Van Willigen, B.; Ranke, J.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models, R Package, Version: 3-1; 2017. Available online: https://cran.r-project.org/package=nlme (accessed on 1 October 2021).
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Wildi, O. Data Analysis in Vegetation Ecology, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2013; p. 330. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Community Ecology Package; 2019; pp. 1–296. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; ISBN 0387874577. [Google Scholar]
- Ryan, T.A. Significance tests for multiple comparisons of proportions, variances, and other statistics. Psychol. Bull. 1960, 57, 318–328. [Google Scholar] [CrossRef] [PubMed]
- IBM Corp. IBM SPSS Statistics for Windows, Version 20.0; IBM Corp: Armonk, NY, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/index.html (accessed on 1 October 2021).
- Marchesini, E.; Montà, L.D. Observations on natural enemies of Lobesia botrana (Den. & Schiff.) (Lepidoptera Tortricidae) in Venetian vineyards. Boll. Di Zool. Agrar. E Di Bachic. 1994, 26, 201–230. [Google Scholar]
- Addante, R.; Di Gioia, S.; Calculli, C.; Pollice, A. The impact of spiders (Araneae) on Lobesia botrana (Denis & Schiffermüller) population density. IOBC/WPRS Bull. 2008, 36, 221–231. [Google Scholar]
- Gonçalves, F.; Carlos, C.; Crespi, A.; Villemant, C.; Trivellone, V.; Goula, M.; Canovai, R.; Zina, V.; Crespo, L.; Pinheiro, L.; et al. The functional agrobiodiversity in the Douro demarcated region viticulture: Utopia or reality? Arthropods as a case-study—A review. Ciênc. Téc. Vitivinícola 2019, 34, 102–114. [Google Scholar] [CrossRef]
- Sentenac, G. Les antagonistes naturels d’Empoasca vitis Göth en Bourgogne. In Etude de Faisabilité D’une Lutte Biologique par Augmentation; Mondiaviti: Bordeaux, France, 1 et 2 Décembre 2004; pp. 25–37. [Google Scholar]
- Huseynov, E.F. Natural prey of the lynx spider Oxyopes lineatus (Araneae: Oxyopidae). Entomol. Fenn. 2007, 18, 144–148. [Google Scholar] [CrossRef]
- Harwood, J.D.; Obrycki, J.J. Web-site selection strategies of linyphiid spiders in alfalfa: Implications for biological control. Biocontrol 2007, 52, 451–467. [Google Scholar] [CrossRef]
- Costello, M.J.; Daane, K.M. Spider (Araneae) species composition and seasonal abundance in San Joaquin Valley grape vineyards. Environ. Entomol. 1995, 24, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Hanna, R.; Zalom, F.G.; Roltsch, W.J. Relative impact of spider predation and cover crop on population dynamics of Erythroneura variabilis in a raisin grape vineyard. Entomol. Exp. Appl. 2003, 107, 177–191. [Google Scholar] [CrossRef]
- Hogg, B.N.; Daane, K.M. Diversity and invasion within a predator community: Impacts on herbivore suppression. J. Appl. Ecol. 2011, 48, 453–461. [Google Scholar] [CrossRef]
- Addante, R.; Pesarini, C.; Ranieri, G. Faunistic and ecological aspects of spiders (Araneae) on vineyards. IOBC/WPRS Bull. 2006, 29, 221–234. [Google Scholar]
- Wisniewska, J.; Prokopy, R.J. Pesticide effect on faunal composition, abundance, and body length of spiders (Araneae) in apple orchards. Environ. Entomol. 1997, 26, 763–766. [Google Scholar] [CrossRef]
- Mazzia, C.; Capowiez, Y.; Marliac, G.; Josselin, D.; Pasquet, A. Spinosad application in an apple orchard affects both the abundance of the spider Araneus diadematus and its web construction behaviour. Ecotoxicology 2020, 29, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Pekár, S. Foraging mode: A factor affecting the susceptibility of spiders (Araneae) to insecticide applications. Pestic. Sci. 1999, 55, 1077–1082. [Google Scholar] [CrossRef]
- Pekár, S. Effect of IPM practices and conventional spraying on spider population dynamics in an apple orchard. Agric. Ecosyst. Environ. 1999, 73, 155–166. [Google Scholar] [CrossRef]
- Nobre, T.; Meierrose, C.; De Oliveira, N.G. Comparison of sampling techniques for vineyard foliage spiders (Araneae). In Proceedings of the 18th European Colloquium of Arachnology, High Tatras, Slovakia, 12–17 July 1999; Gajdoš, P., Pekar, S., Eds.; Ekológia (Bratislava), 19, suppl. 3/2000. pp. 201–206. [Google Scholar]
- Trdan, S.; Andjus, L.; Raspudić, E.; Kač, M. Distribution of Aeolothrips intermedius Bagnall (Thysanoptera: Aeolothripidae) and its potential prey Thysanoptera species on different cultivated host plants. J. Pest Sci. 2005, 78, 217–226. [Google Scholar] [CrossRef]
- Daane, K.M.; Almeida, R.P.P.; Bell, V.A.; Walker, J.T.S.; Botton, M.; Fallahzadeh, M.; Mani, M.; Miano, J.L.; Sforza, R.; Walton, V.M.; et al. Biology and management of mealybugs in vineyards. In Arthropod Management in Vineyards: Pest, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaac, R., Eds.; Springer Science + Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 271–307. [Google Scholar] [CrossRef]
- Arzone, A.; Vidano, C.; Arnò, C. Predators and parasitoids of Empoasca vitis and Zygina rhamni (Rhynchota Auchrenorrhyncha). In Proceedings of the 6th Auchenorrhyncha Meeting, Turin, Italy, 7–11 September 1987; Vidano, C., Arzone, A., Eds.; CNR-IPRA: Turin, Italy, 1988; pp. 623–629. [Google Scholar]
- Girolami, V.; Duso, C.; Refatti, E.; Osler, R. Lotta Integrata in Viticoltura, Malattie Della Vite; IRIPA-Coldiretti Edizioni: Treviso, Italy, 1989. [Google Scholar]
- Genini, M. Antagonistes de la cicadelle verte et des vers de la grappe dans le vignoble valaisan et les milieux environnants. Rev. suisse Vitic. Arboric. Hortic. 2000, 32, 153–162. [Google Scholar]
- Benhadi-Marín, J.; Pereira, J.A.; Santos, S.A.P. Effects of kaolin particle films on the life span of an orb-weaver spider. Chemosphere 2016, 144, 918–924. [Google Scholar] [CrossRef]
- Reichert, S.; Lockley, T. Spiders as biological control agents. Annu. Rev. Entomol. 1984, 29, 299–320. [Google Scholar] [CrossRef]
- Nyffeler, M.; Dean, D.A.; Sterling, W.L. Evaluation of the importance of the striped lynx spider, Oxyopes salticus (Araneae: Oxyopidae), as a predator in Texas cotton. Environ. Entomol. 1987, 16, 1114–1123. [Google Scholar] [CrossRef]
- Huseynov, E.F. Natural prey of the jumping spider Menemerus taeniatus (Araneae: Salticidae). Eur. J. Entomol. 2005, 102, 797–799. [Google Scholar] [CrossRef] [Green Version]
- Bengochea, P.; Amor, F.; Saelices, R.; Hernando, S.; Budia, F.; Adán, Á.; Medina, P. Kaolin and copper-based products applications: Ecotoxicology on four natural enemies. Chemosphere 2013, 91, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, N.; Abdallah, S.B. Laboratory evaluation of side-effects of kaolin on two predator species found on olive groves. Tunis. J. Plant Prot. 2016, 11, 83–90. [Google Scholar]
- Tacoli, F.; Bell, V.A.; Cargnus, E.; Pavan, F. Insecticidal activity of natural products against vineyard mealybugs (Hemiptera: Pseudococcidae). Crop. Prot. 2018, 111, 50–57. [Google Scholar] [CrossRef]
- Joubert, P.H.; Grové, T.; De Beer, M.S.; Steyn, W.P. Evaluation of kaolin (Surround® Wp) in an IPM program on mangoes in South Africa. Acta Hortic. 2004, 645, 493–499. [Google Scholar] [CrossRef]
- Skouras, P.J.; Stathas, G.J.; Demopoulos, V.; Louloudakis, G.; Margaritopoulos, J.T. The effect of five insecticides on the predators Coccinella septempunctata and Hippodamia variegata. Phytoparasitica 2019, 47, 197–205. [Google Scholar] [CrossRef]
- Bengochea, P.; Saelices, R.; Amor, F.; Adán, Á.; Budia, F.; del Estal, P.; Viñuela, E.; Medina, P. Non-target effects of kaolin and coppers applied on olive trees for the predatory lacewing Chrysoperla carnea. Biocontrol Sci. Technol. 2014, 24, 625–640. [Google Scholar] [CrossRef]
- Bestete, L.R.; Torres, J.B.; Pereira, F.F. Harmonious interaction of kaolin and two insect predator species in plant protection. Int. J. Pest Manag. 2018, 64, 166–172. [Google Scholar] [CrossRef]
- Strapazzon, A.; Pavan, F.; Cristoferi, K. Monitoraggio dei fitomizi della vite con trappole cromotropiche: Risposta di Drepanothrips reuteri Uzel al colore giallo ed alle dimensioni delle trappole. Frustula entomologica 1990, 11, 9–18. [Google Scholar]
- Pavan, F.; Cargnus, E.; Tacoli, F.; Zandigiacomo, P. Standardization and criticism of sampling procedures using sticky card traps: Monitoring sap-sucking insect pests and Anagrus atomus inhabiting European vineyards. Bull. Insectol. 2021, 74, 291–306. [Google Scholar]
- Pennington, T.; Kolb, S.; Kaiser, J.; Hoffmann, C.; Entling, M.H. Does minimal pruning and reduced fungicide use impact spiders in the grapevine canopy? J. Arachnol. 2019, 47, 381–384. [Google Scholar] [CrossRef]
- Legner, E.F.; Oatman, E.R. Spiders on apple in Wisconsin and their abundance in natural and two artificial environments. Can. Entomol. 1964, 96, 1202–1207. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Date | Timing | BBCH |
|---|---|---|
| Vineyards A and B—2015 | ||
| 17 June | LR | 73 |
| 18 June | First kaolin spraying | 75 |
| 24 June | Second kaolin spraying | 75 |
| Vineyard A—2016 | ||
| 10 June | LR and First kaolin spraying | 71 |
| 11–19 June | Rain: about 90 mm in 8 days http://www.osmer.fvg.it/archivio.php (accessed on 20 June 2016) | 71–74 |
| 20 June | Second kaolin spraying | 74 |
| 28 June | Third kaolin spraying | 77 |
| Vineyard C—2016 | ||
| 10 June | LR and First kaolin spraying | 71 |
| 24 June | Second kaolin spraying | 77 |
| Indices and Functional Guilds | Factors | F | D.F. | p |
|---|---|---|---|---|
| Species diversity (Shannon index H’) | Intercept | 63.305 | 1, 14 | <0.0001 |
| kaolin | 1.790 | 1, 14 | 0.202 | |
| LR | 0.536 | 1, 14 | 0.476 | |
| year | 0.039 | 1, 14 | 0.847 | |
| kaolin:LR | 0.611 | 1, 14 | 0.448 | |
| kaolin:year | 0.050 | 1, 14 | 0.826 | |
| year:LR | 0.594 | 1, 14 | 0.454 | |
| kaolin:LR:year | 0.485 | 1, 14 | 0.498 | |
| Functional richness | Intercept | 64.774 | 1, 14 | <0.0001 |
| kaolin | 1.307 | 1, 14 | 0.272 | |
| LR | 0.001 | 1, 14 | 0.982 | |
| year | 4.024 | 1, 14 | 0.065 | |
| kaolin:LR | 0.907 | 1, 14 | 0.357 | |
| kaolin:year | 0.007 | 1, 14 | 0.934 | |
| year:LR | 0.681 | 1, 14 | 0.423 | |
| kaolin:LR:year | 1.398 | 1, 14 | 0.257 | |
| Functional evenness | Intercept | 25.068 | 1, 14 | 0.000 |
| kaolin | 1.982 | 1, 14 | 0.181 | |
| LR | 0.139 | 1, 14 | 0.715 | |
| year | 0.587 | 1, 14 | 0.456 | |
| kaolin:LR | 4.024 | 1, 14 | 0.065 | |
| kaolin:year | 0.017 | 1, 14 | 0.899 | |
| year:LR | 0.097 | 1, 14 | 0.760 | |
| kaolin:LR:year | 0.676 | 1, 14 | 0.425 | |
| Functional distance | Intercept | 45.589 | 1, 13 | <0.0001 |
| kaolin | 0.272 | 1, 13 | 0.611 | |
| LR | 0.048 | 1, 13 | 0.830 | |
| year | 0.100 | 1, 13 | 0.757 | |
| kaolin:LR | 1.426 | 1, 13 | 0.254 | |
| kaolin:year | 1.841 | 1, 13 | 0.198 | |
| year:LR | 0.288 | 1, 13 | 0.600 | |
| kaolin:LR:year | 0.288 | 1, 13 | 0.601 | |
| Body size | Intercept | 92.915 | 1, 13 | <0.0001 |
| kaolin | 0.181 | 1, 13 | 0.677 | |
| LR | 1.039 | 1, 13 | 0.327 | |
| year | 0.857 | 1, 13 | 0.372 | |
| kaolin:LR | 0.952 | 1, 13 | 0.347 | |
| kaolin:year | 0.516 | 1, 13 | 0.485 | |
| year:LR | 0.822 | 1, 13 | 0.381 | |
| kaolin:LR:year | 0.135 | 1, 13 | 0.719 | |
| log (hunter % + 0.1) | Intercept | 4.178 | 1, 13 | 0.062 |
| kaolin | 1.313 | 1, 13 | 0.273 | |
| LR | 0.001 | 1, 13 | 0.975 | |
| year | 0.405 | 1, 13 | 0.536 | |
| kaolin:LR | 0.094 | 1, 13 | 0.764 | |
| kaolin:year | 0.026 | 1, 13 | 0.873 | |
| year:LR | 1.613 | 1, 13 | 0.226 | |
| kaolin:LR:year | 2.174 | 1, 13 | 0.164 |
| Taxon | Kaolin | LR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Kaolin | F | D.F. | p | No LR | LR | F | D.F. | p | |
| 2015 | ||||||||||
| Total spiders | 5.0 ± 1.4 | 3.8 ± 1.4 | 2.308 | 1, 12 | 0.155 | 4.4 ± 1.4 | 4.4 ± 1.4 | 0.000 | 1, 12 | 1.000 |
| Web-builders | 1.1 ± 1.0 | 1.0 ± 0.7 | 0.034 | 1, 12 | 0.857 | 1.3 ± 1.0 | 0.9 ± 0.7 | 0.303 | 1, 12 | 0.592 |
| Araneidae | 0.9 ± 0.9 | 0.6 ± 0.6 | 0.509 | 1, 12 | 0.489 | 0.9 ± 0.6 | 0.6 ± 0.6 | 0.057 | 1, 12 | 0.816 |
| Linyphiidae | 0.1 ± 0.3 | 0.1 ± 0.3 | − | − | − | 0.3 ± 0.3 | 0.0 ± 0.0 | − | − | − |
| Theridiidae | 0.1 ± 0.3 | 0.3 ± 0.3 | − | − | − | 0.1 ± 0.3 | 0.3 ± 0.3 | − | − | − |
| Hunters | 3.9 ± 1.3 | 2.8 ± 1.4 | 3.522 | 1, 12 | 0.085 | 3.1 ± 1.3 | 3.5 ± 1.4 | 0.391 | 1, 12 | 0.543 |
| Thomisidae | 1.8 ± 1.0 | 2.1 ± 1.2 | 0.370 | 1, 12 | 0.554 | 1.9 ± 1.1 | 2.0 ± 1.1 | 0.041 | 1, 12 | 0.843 |
| Gnaphosidae | 0.3 ± 0.3 | 0.3 ± 0.3 | − | 0.3 ± 0.3 | 0.3 ± 0.3 | − | − | − | ||
| Oxyopidae | 1.5 ± 1.1 | 0.1 ± 0.3 | 6.153 | 1, 12 | 0.029 | 0.5 ± 0.5 | 1.1 ± 1.0 | 1.271 | 1, 12 | 0.282 |
| Salticidae | 0.4 ± 0.4 | 0.3 ± 0.5 | − | − | − | 0.5 ± 0.6 | 0.1 ± 0.3 | − | − | − |
| 2016 | ||||||||||
| Total spiders | 4.4 ± 1.6 | 3.8 ± 1.6 | 0.311 | 1, 12 | 0.587 | 3.3 ± 1.6 | 4.9 ± 1.6 | 2.104 | 1, 12 | 0.173 |
| Web-builders | 1.4 ± 0.9 | 0.9 ± 0.7 | 0.889 | 1, 12 | 0.364 | 0.9 ± 0.8 | 1.4 ± 0.8 | 0.889 | 1, 12 | 0.364 |
| Araneidae | 1.1 ± 0.6 | 0.3 ± 0.3 | 7.000 | 1, 12 | 0.021 | 0.5 ± 0.5 | 0.9 ± 0.6 | 1.286 | 1, 12 | 0.279 |
| Linyphiidae | 0.3 ± 0.3 | 0.5 ± 0.5 | − | − | − | 0.4 ± 0.4 | 0.4 ± 0.4 | − | − | − |
| Theridiidae | 0.0 ± 0.0 | 0.1 ± 0.3 | − | − | − | 0.0 ± 0.0 | 0.1 ± 0.3 | − | − | − |
| Hunters | 3.0 ± 1.6 | 2.9 ± 1.7 | 0.014 | 1, 12 | 0.908 | 2.4 ± 1.4 | 3.5 ± 1.8 | 1.120 | 1, 12 | 0.311 |
| Thomisidae | 1.1 ± 1.0 | 2.3 ± 1.4 | 2.505 | 1, 12 | 0.139 | 1.1 ± 0.8 | 2.3 ± 1.5 | 2.505 | 1, 12 | 0.139 |
| Gnaphosidae | 0.1 ± 0.3 | 0.3 ± 0.3 | − | − | − | 0.1 ± 0.3 | 0.3 ± 0.3 | − | − | − |
| Miturgidae | 0.1 ± 0.3 | 0.0 ± 0.0 | − | − | − | 0.0 ± 0.0 | 0.1 ± 0.3 | − | − | − |
| Oxyopidae | 0.3 ± 0.3 | 0.0 ± 0.0 | − | − | − | 0.1 ± 0.3 | 0.1 ± 0.3 | − | − | − |
| Salticidae | 1.3 ± 0.8 | 0.3 ± 0.3 | 5.647 | 1, 12 | 0.035 | 0.9 ± 0.7 | 0.6 ± 0.5 | 0.063 | 1, 12 | 0.563 |
| Sparassidae | 0.1 ± 0.3 | 0.1 ± 0.3 | − | − | − | 0.1 ± 0.3 | 0.1 ± 0.3 | − | − | − |
| Taxon | Kaolin | LR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Kaolin | F | D.F. | p | No LR | LR | F | D.F. | p | |
| Insects | ||||||||||
| Aeolothrips sp. | 59.6 ± 10.5 | 71.0 ± 20.2 | 0.444 | 1, 12 | 0.518 | 38.0 ± 8.4 | 92.6 ± 15.6 | 10.230 | 1, 12 | 0.008 |
| Scymninae | 3.1 ± 0.5 | 6.4 ± 1.0 | 8.380 | 1, 12 | 0.013 | 3.9 ± 10.9 | 5.6 ± 1.1 | 2.430 | 1, 12 | 0.145 |
| Coccinellidae non Scymninae | 3.9 ± 0.6 | 2.8 ± 0.6 | 1.827 | 1, 12 | 0.201 | 2.6 ± 0.6 | 4.0 ± 0.6 | 2.729 | 1, 12 | 0.124 |
| Chrysoperla carnea s.l. | 2.3 ± 0.69 | 1.0 ± 0.4 | 3.000 | 1, 12 | 0.109 | 1.8 ± 0.4 | 1.5 ± 0.7 | 0.120 | 1, 12 | 0.735 |
| Orius sp. | 2.6 ± 0.8 | 1.0 ± 0.3 | 3.596 | 1, 12 | 0.082 | 2.4 ± 0.7 | 1.3 ± 0.6 | 1.723 | 1, 12 | 0.214 |
| Spiders | ||||||||||
| Total | 15.8 ± 2.5 | 12.5 ± 1.4 | 1.257 | 1, 12 | 0.284 | 12.4 ± 21.5 | 15.9 ± 2.4 | 1,457 | 1, 12 | 0.251 |
| Web-builders | 6.6 ± 1.3 | 5.0 ± 0.5 | 1.317 | 1, 12 | 0.274 | 4.8 ±0.5 | 6.9 ± 1.3 | 2.252 | 1, 12 | 0.159 |
| Hunters | 9.1 ± 1.4 | 7.5 ± 1.2 | 0.719 | 1, 12 | 0.413 | 7.6 ± 1.3 | 9.0 ± 1.3 | 0.515 | 1, 12 | 0.487 |
| Thomisidae | 4.9 ± 1.0 | 3.6 ± 0.7 | 1.181 | 1, 12 | 0.298 | 3.1 ± 0.7 | 5.4 ± 1.9 | 3.827 | 1, 12 | 0.074 |
| Gnaphosidae | 0.4 ± 0.3 | 0.3 ± 0.2 | 0.176 | 1, 12 | 0.682 | 0.1 ± 0.1 | 0.5 ± 0.3 | 1.588 | 1, 12 | 0.232 |
| Oxyopidae | 0.5 ± 0.3 | 0.9 ± 0.4 | 0.574 | 1, 12 | 0.463 | 0.5 ± 0.3 | 0.9 ± 0.4 | 0.574 | 1, 12 | 0.463 |
| Salticidae | 3.4 ± 0.9 | 2.0 ± 0.8 | 1.435 | 1, 12 | 0.254 | 3.4 ± 0.9 | 2.0 ± 0.7 | 1.435 | 1, 12 | 0.254 |
| Indices and Functional Guilds | Factors | F | D.F. | p |
|---|---|---|---|---|
| Species diversity (Shannon index H’) | Intercept | 176.940 | 1, 14 | <0.0001 |
| kaolin | 1.880 | 1, 14 | 0.192 | |
| LR | 6.171 | 1, 14 | 0.026 | |
| year | 19.129 | 1, 14 | 0.001 | |
| kaolin:LR | 0.016 | 1, 14 | 0.902 | |
| kaolin:year | 0.167 | 1, 14 | 0.689 | |
| year:LR | 1.811 | 1, 14 | 0.200 | |
| kaolin:LR:year | 3.143 | 1, 14 | 0.098 | |
| Functional richness | Intercept | 155.792 | 1, 13 | <0.0001 |
| kaolin | 0.068 | 1, 13 | 0.798 | |
| LR | 1.903 | 1, 13 | 0.191 | |
| year | 9.958 | 1, 13 | 0.008 | |
| kaolin:LR | 2.115 | 1, 13 | 0.170 | |
| kaolin:year | 0.008 | 1, 13 | 0.928 | |
| year:LR | 0.025 | 1, 13 | 0.877 | |
| kaolin:LR:year | 0.413 | 1, 13 | 0.532 | |
| Functional evenness | Intercept | 85.379 | 1, 14 | <0.0001 |
| kaolin | 0.312 | 1, 14 | 0.585 | |
| LR | 7.818 | 1, 14 | 0.014 | |
| year | 7.094 | 1, 14 | 0.019 | |
| kaolin:LR | 2.389 | 1, 14 | 0.145 | |
| kaolin:year | 1.006 | 1, 14 | 0.333 | |
| year:LR | 2.845 | 1, 14 | 0.114 | |
| kaolin:LR:year | 0.985 | 1, 14 | 0.338 | |
| Functional distance | Intercept | 133.834 | 1, 13 | <0.0001 |
| kaolin | 0.224 | 1, 13 | 0.644 | |
| LR | 2.945 | 1, 13 | 0.110 | |
| year | 4.493 | 1, 13 | 0.054 | |
| kaolin:LR | 0.038 | 1, 13 | 0.849 | |
| kaolin:year | 0.013 | 1, 13 | 0.913 | |
| year:LR | 0.566 | 1, 13 | 0.465 | |
| kaolin:LR:year | 0.377 | 1, 13 | 0.550 | |
| Body size | Intercept | 172.178 | 1, 13 | <0.0001 |
| kaolin | 1.332 | 1, 13 | 0.269 | |
| LR | 3.646 | 1, 13 | 0.079 | |
| year | 0.838 | 1, 13 | 0.377 | |
| kaolin:LR | 0.002 | 1, 13 | 0.968 | |
| kaolin:year | 0.270 | 1, 13 | 0.612 | |
| year:LR | 0.441 | 1, 13 | 0.518 | |
| kaolin:LR:year | 0.022 | 1, 13 | 0.886 | |
| log (hunter % + 0.1) | Intercept | 18.915 | 1, 13 | 0.001 |
| kaolin | 0.055 | 1, 13 | 0.818 | |
| LR | 0.154 | 1, 13 | 0.701 | |
| year | 2.916 | 1, 13 | 0.112 | |
| kaolin:LR | 1.762 | 1, 13 | 0.207 | |
| kaolin:year | 0.483 | 1, 13 | 0.499 | |
| year:LR | 2.805 | 1, 13 | 0.118 | |
| kaolin:LR:year | 1.408 | 1, 13 | 0.257 |
| Taxon | Kaolin | LR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Kaolin | F | D.F. | p | No LR | LR | F | D.F. | p | |
| Vineyard B | ||||||||||
| Total spiders | 9.9 ± 2.4 | 10.1 ± 3.0 | 0.029 | 1, 12 | 0.868 | 10.4 ± 32.4 | 9.6 ± 3.0 | 0.261 | 1, 12 | 0.619 |
| Web-builders | 3.8 ± 1.4 | 3.9 ± 2.0 | 0.014 | 1, 12 | 0.907 | 4.1 ± 1.8 | 3.5 ± 1.7 | 0.352 | 1, 12 | 0.564 |
| Araneidae | 2.5 ± 1.3 | 1.5 ± 1.2 | 0.591 | 1, 12 | 0.457 | 2.3 ± 1.2 | 1.8 ± 1.2 | 0.024 | 1, 12 | 0.880 |
| Linyphiidae | 0.9 ± 0.8 | 1.6 ± 1.5 | 0.591 | 1, 12 | 0.457 | 1.3 ± 1.4 | 1.3 ± 1.0 | 0.000 | 1, 12 | 1.000 |
| Theridiidae | 0.4 ± 0.4 | 0.8 ± 0.8 | 0.000 | 1, 12 | 1.000 | 0.6 ± 0.6 | 0.5 ± 0.6 | 0.667 | 1, 12 | 0.430 |
| Hunters | 6.1 ± 2.1 | 6.3 ± 2.2 | 0.009 | 1, 12 | 0.926 | 6.3 ± 2.1 | 6.1 ± 2.3 | 0.009 | 1, 12 | 0.926 |
| Thomisidae | 2.9 ± 1.7 | 3.8 ± 2.2 | 0.452 | 1, 12 | 0.514 | 3.1 ± 1.7 | 3.5 ± 2.1 | 0.083 | 1, 12 | 0.778 |
| Gnaphosidae | 0.5 ± 0.7 | 1.0 ± 0.9 | 0.558 | 1, 12 | 0.469 | 0.9 ± 0.8 | 0.6 ± 0.8 | 0.140 | 1, 12 | 0.715 |
| Oxyopidae | 1.4 ± 1.1 | 1.0 ± 0.7 | 0.252 | 1, 12 | 0.625 | 1.0 ± 0.8 | 1.4 ± 1.1 | 0.252 | 1, 12 | 0.625 |
| Salticidae | 0.9 ± 0.9 | 0.5 ± 0.5 | 0.325 | 1, 12 | 0.579 | 0.8 ± 10.8 | 0.6 ± 0.6 | 0.036 | 1, 12 | 0.852 |
| Sparassidae | 0.5 ± 0.6 | 0.0 ± 0.0 | − | − | − | 0.5 ± 0.6 | 0.0 ± 0.0 | − | − | − |
| Vineyard C | ||||||||||
| Total spiders | 4.4 ± 1.2 | 3.1 ± 1.4 | 2.793 | 1, 12 | 0.121 | 3.9 ± 1.3 | 3.6 ± 1.3 | 0.310 | 1, 12 | 0.588 |
| Web-builders | 1.8 ± 0.9 | 1.4 ± 0.9 | 0.474 | 1, 12 | 0.504 | 1.5 ± 0.9 | 1.6 ± 0.9 | 0.053 | 1, 12 | 0.822 |
| Araneidae | 1.3 ± 0.8 | 0.9 ± 0.5 | 0.692 | 1, 12 | 0.422 | 1.0 ± 0.6 | 1.1 ± 0.7 | 0.077 | 1, 12 | 0.786 |
| Linyphiidae | 0.5 ± 0.6 | 0.5 ± 0.5 | − | − | − | 0.5 ± 0.5 | 0.5 ± 0.6 | − | − | − |
| Hunters | 2.6 ± 1.0 | 1.8 ± 0.9 | 1.862 | 1, 12 | 0.197 | 2.4 ± 1.0 | 2.0 ± 1.0 | 0.828 | 1, 12 | 0.381 |
| Thomisidae | 0.4 ± 0.4 | 0.3 ± 0.3 | − | − | − | 0.1 ± 0.3 | 0.5 ± 0.5 | − | − | − |
| Gnaphosidae | 0.0 ± 0.0 | 0.1 ± 0.3 | − | − | − | 0.1 ± 0.3 | 0.0 ± 0.0 | − | − | − |
| Miturgidae | 0.1 ± 0.3 | 0.0 ± 0.0 | − | − | − | 0.0 ± 0.0 | 0.1 ± 0.3 | − | − | − |
| Oxyopidae | 1.0 ± 0.7 | 0.6 ± 0.5 | 1.080 | 1, 12 | 0.319 | 0.9 ± 0.6 | 0.8 ± 0.7 | 0.120 | 1, 12 | 0.735 |
| Salticidae | 1.1 ± 0.7 | 0.8 ± 0.8 | 0.176 | 1, 12 | 0.682 | 1.3 ± 1.0 | 0.6 ± 0.5 | 1.588 | 1, 12 | 0.232 |
| Taxon | Kaolin | LR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Kaolin | F | D.F. | p | No LR | LR | F | D.F. | p | |
| Insects | ||||||||||
| Aeolothrips sp. | 55.6 ± 12.4 | 92.5 ± 21.2 | 2.020 | 1, 12 | 0.181 | 64.5 ± 19.0 | 83.6 ± 17.7 | 0.543 | 1, 12 | 0.475 |
| Scymninae | 5.9 ± 6.9 | 9.5 ± 8.7 | 2.422 | 1, 12 | 0.146 | 16.3 ± 3.1 | 22.1 ± 5.2 | 1.061 | 1, 12 | 0.323 |
| Coccinellidae non-Scymninae | 6.8 ± 1.4 | 8.4 ± 1.3 | 0.610 | 1, 12 | 0.450 | 7.3 ± 1.6 | 7.9 ± 1.2 | 0.090 | 1, 12 | 0.769 |
| Chrysoperla carnea s.l. | 2.6 ± 0.9 | 2.0 ± 0.5 | 0.316 | 1, 12 | 0.584 | 1.9 ± 0.8 | 2.8 ± 0.6 | 0.620 | 1, 12 | 0.446 |
| Orius sp. | 3.0 ± 0.8 | 0.9 ± 0.4 | 5.070 | 1, 12 | 0.044 | 2.1 ± 0.9 | 1.8 ± 0.5 | 0.158 | 1, 12 | 0.698 |
| Spiders | ||||||||||
| Total spiders | 19.4 ± 2.8 | 20.8 ± 2.9 | 0.104 | 1, 12 | 0.753 | 19.1 ± 3.3 | 21.0 ± 2.3 | 0.193 | 1, 12 | 0.668 |
| Web-builders | 7.1 ± 1.4 | 8.8 ± 1.5 | 0.497 | 1, 12 | 0.494 | 7.6 ± 2.0 | 8.3 ± 1.0 | 0.073 | 1, 12 | 0.791 |
| Hunters | 12.2 ± 2.2 | 12.0 ± 1.6 | 0.008 | 1, 12 | 0.931 | 11.5 ± 31.7 | 12.8 ± 2.1 | 0.194 | 1, 12 | 0.668 |
| Thomisidae | 6.1 ± 2.0 | 6.4 ± 1.0 | 0.011 | 1, 12 | 0.918 | 5.9 ± 1.2 | 6.6 ± 1.9 | 0.099 | 1, 12 | 0.758 |
| Gnaphosidae | 1.0 ± 0.3 | 0.5 ± 0.3 | 1.714 | 1, 12 | 0.215 | 0.8 ± 0.3 | 0.8 ± 0.3 | 0.000 | 1, 12 | 1.000 |
| Oxyopidae | 2.0 ± 0.6 | 2.5 ± 0.7 | 0.289 | 1, 12 | 0.601 | 2.4 ± 0.8 | 2.1 ± 0.4 | 0.072 | 1, 12 | 0.793 |
| Salticidae | 3.0 ± 0.8 | 2.5 ± 0.7 | 0.211 | 1, 12 | 0.655 | 2.3 ± 0.8 | 3.3 ± 0.7 | 0.842 | 1, 12 | 0.377 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cargnus, E.; Tacoli, F.; Boscutti, F.; Zandigiacomo, P.; Pavan, F. Side Effects of Kaolin and Bunch-Zone Leaf Removal on Generalist Predators in Vineyards. Insects 2023, 14, 126. https://doi.org/10.3390/insects14020126
Cargnus E, Tacoli F, Boscutti F, Zandigiacomo P, Pavan F. Side Effects of Kaolin and Bunch-Zone Leaf Removal on Generalist Predators in Vineyards. Insects. 2023; 14(2):126. https://doi.org/10.3390/insects14020126
Chicago/Turabian StyleCargnus, Elena, Federico Tacoli, Francesco Boscutti, Pietro Zandigiacomo, and Francesco Pavan. 2023. "Side Effects of Kaolin and Bunch-Zone Leaf Removal on Generalist Predators in Vineyards" Insects 14, no. 2: 126. https://doi.org/10.3390/insects14020126
APA StyleCargnus, E., Tacoli, F., Boscutti, F., Zandigiacomo, P., & Pavan, F. (2023). Side Effects of Kaolin and Bunch-Zone Leaf Removal on Generalist Predators in Vineyards. Insects, 14(2), 126. https://doi.org/10.3390/insects14020126