
Mating Competitiveness of Male Spodoptera frugiperda (Smith) Irradiated by X-rays

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Methods of Irradiating and Feeding Tested Insects
2.2. Field Corn Cage Coverage Methods
2.3. Experimental Methods
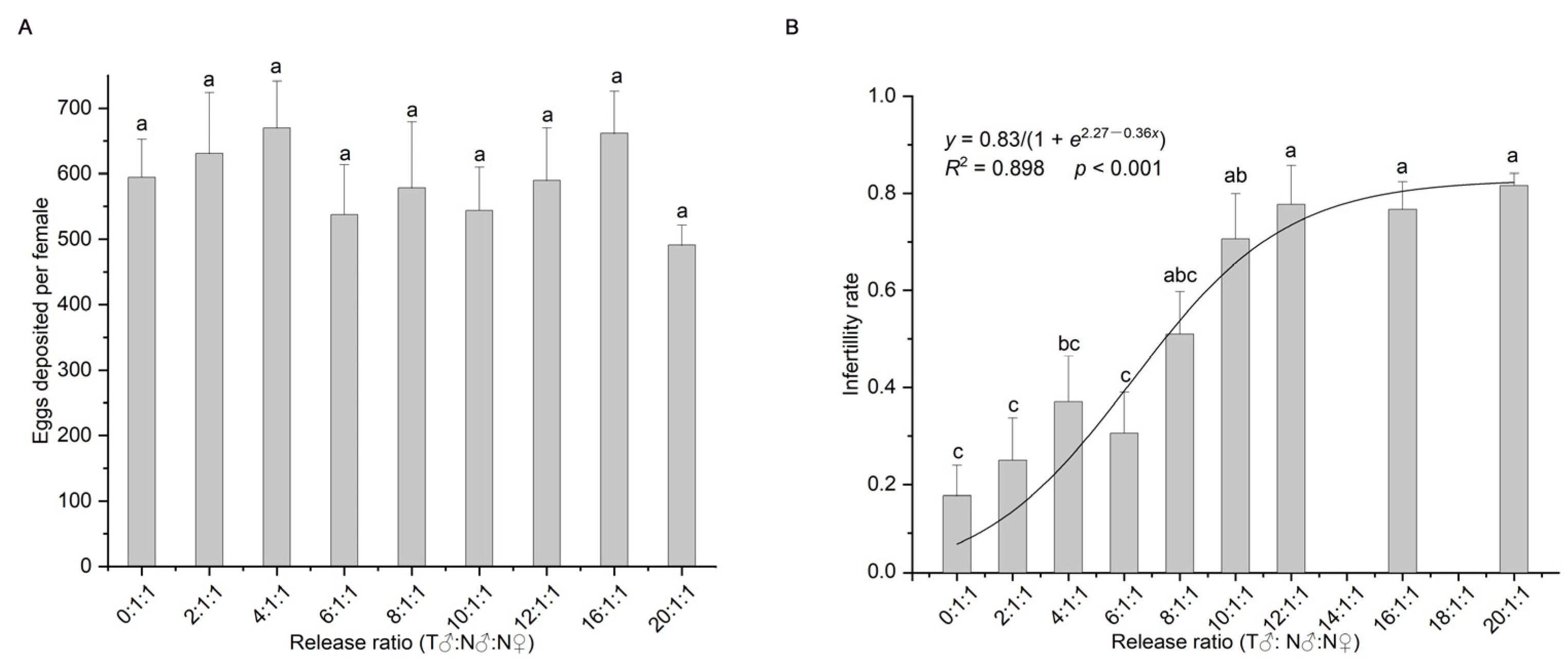
2.3.1. Effect of the Release Ratio of Irradiated to Non-Irradiated Males on the Sterility Rate of the Offspring
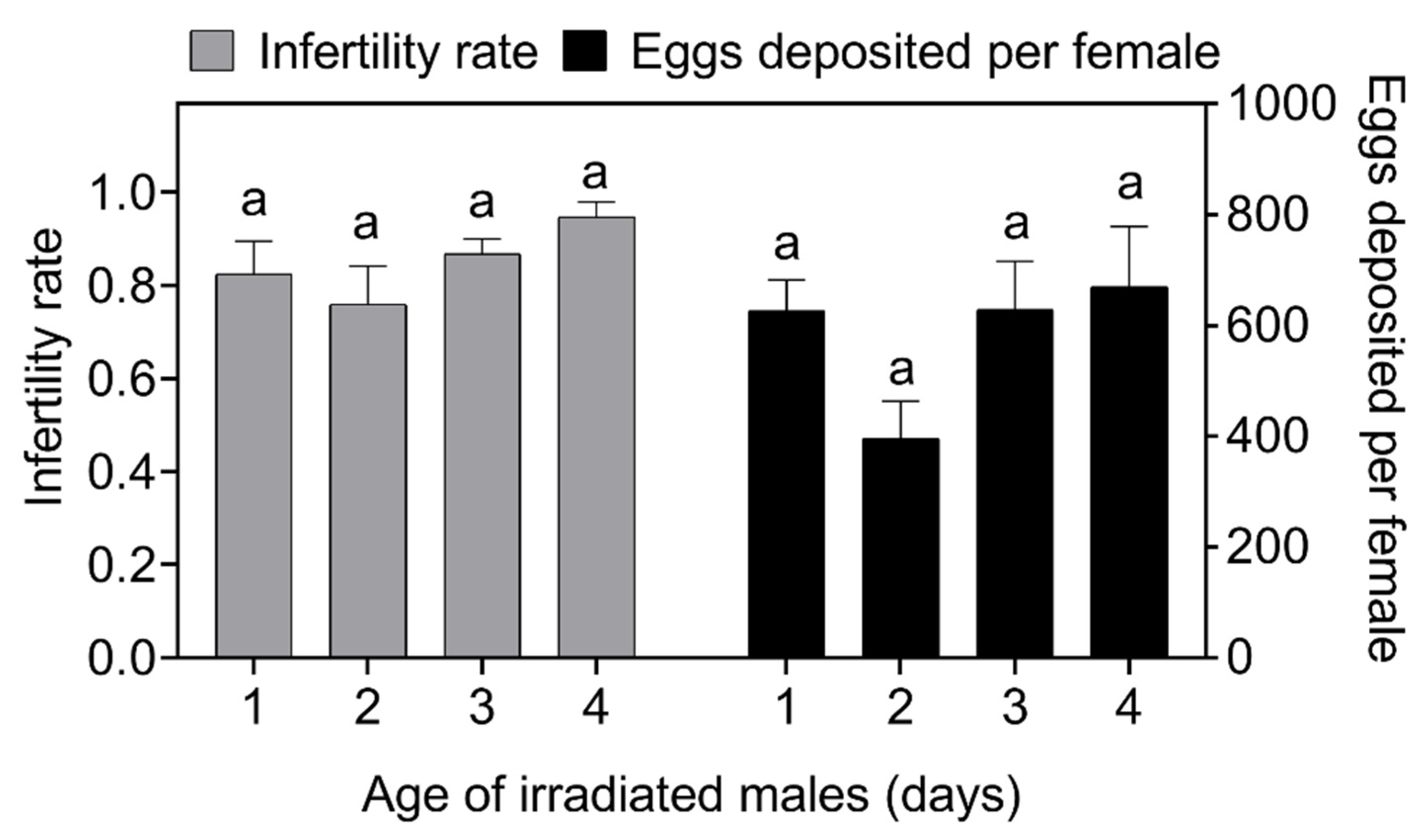
2.3.2. Selection of the Age of Irradiated Males at Release
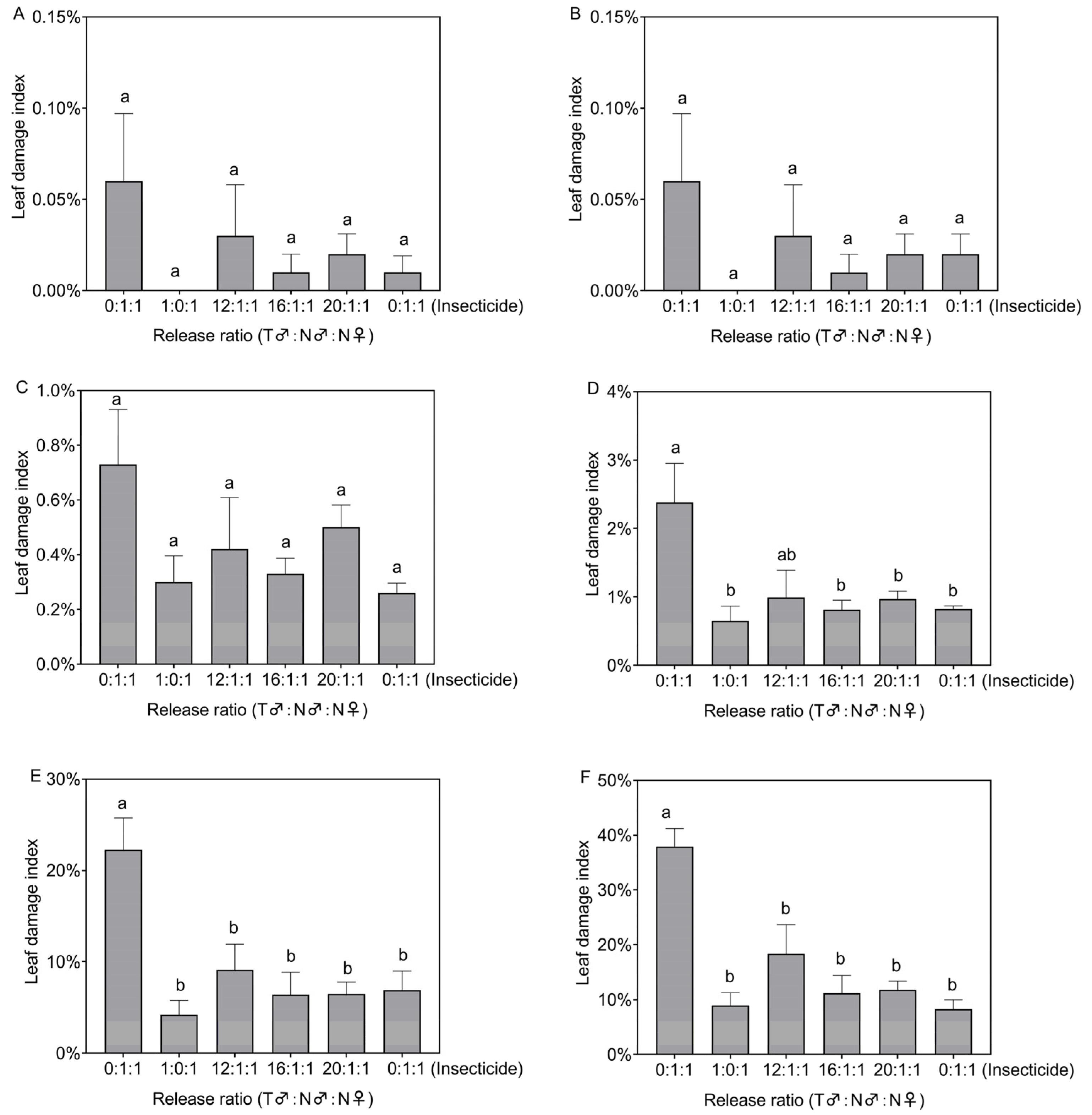
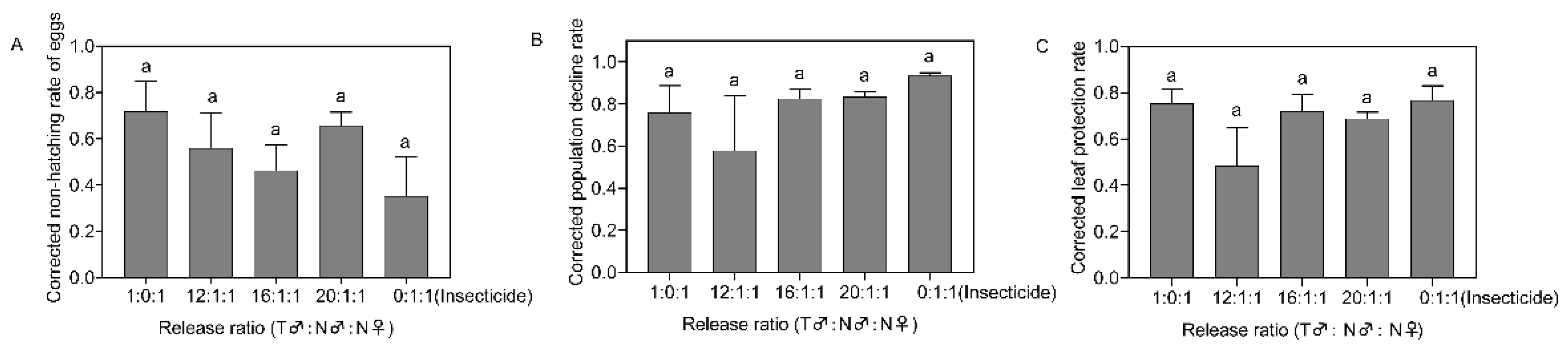
2.3.3. Confined-Field Release Test of Irradiated Males
2.4. Data Analysis
3. Results
3.1. Effect of the Release Ratio of Irradiated to Non-Irradiated Males on the Sterility Rate of Offspring
3.2. Selection of the Age of Irradiated Males at Release
3.3. Control Effects of Irradiated Males after Release in the Field Cages
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.; Gu, S.H.; Zhang, L.; Gao, X.W. Research status and prospects of Spodoptera frugiperda (Lepidoptera: Noctuidae) in China. Acta Entomol. Sin. 2020, 63, 624–638. [Google Scholar] [CrossRef]
- He, L.M.; Ge, S.S.; Chen, Y.C.; Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. The developmental threshold temperature, effective accumulated temperature and prediction model of developmental duration of fall armyworm, Spodoptera frugiperda. Plant Prot. 2019, 45, 18–26. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agr. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Marenco, R.J.; Foster, R.E.; Sanchez, C.A. Sweet corn response to fall armyworm (Lepidoptera: Noctuidae) damage during vegetative growth. J. Econ. Entomol. 1992, 84, 1285–1292. [Google Scholar] [CrossRef]
- Hruska, A.J.; Gladstone, S.M. Effect of period and level of infestation of the fall armyworm, Spodoptera frugiperda, on irrigated maize yield. Fla. Entomol. 1988, 71, 249–254. [Google Scholar] [CrossRef]
- Yang, X.; Wyckhuys, K.A.G.; Jia, X.; Nie, F.; Wu, K. Fall armyworm invasion heightens pesticide expenditure among Chinese smallholder farmers. J. Environ. Manag. 2021, 282, 111949. [Google Scholar] [CrossRef]
- Diez-Rodriguez, E.G.; Celso, O. Herança da resistência de Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) a lambda-Cialotrina. Neotrop. Entomol. 2001, 30, 311–316. [Google Scholar] [CrossRef]
- Yu, S.J. Detection and biochemical characterization of insecticide resistance in fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1992, 85, 675–682. [Google Scholar] [CrossRef]
- Yu, S.J. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Phys. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Yu, S.J.; Nguyen, S.N.; Abo-Elghar, G.E. Biochemical characteristics of insecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Phys. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Zhao, S.Y.; Sun, X.X.; Zhang, H.W.; Yang, X.M.; Wu, K.M. Laboratory test on the control efficacy of common chemical insecticides against Spodoptera frugiperda. Plant Prot. 2019, 45, 10–14. [Google Scholar] [CrossRef]
- Zhang, D.D.; Xiao, Y.T.; Xu, P.J.; Yang, X.M.; Wu, Q.L.; Wu, K.M. Insecticide resistance monitoring for the invasive populations of fall armyworm, Spodoptera frugiperda in China. J. Integr. Agr. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Lachance, L.E.; Richard, R.D.; Proshold, F.I. Radiation response in the pink bollworm: A comparative study of sperm bundle production, sperm transfer, and oviposition response elicited by native and laboratory-reared males. Environ. Entomol. 1975, 4, 321–324. [Google Scholar] [CrossRef]
- Baumhover, A.H.; Graham, A.J.; Bitter, B.A.; Hopkins, D.E.; New, W.D.; Dudley, F.H.; Bushland, R.C. Screw-worm control through release of sterilized flies. J. Econ. Entomol. 1955, 48, 462–466. [Google Scholar] [CrossRef]
- Knipling, E.F. Use of insects for their own destruction. J. Econ. Entomol. 1960, 53, 415–420. [Google Scholar] [CrossRef]
- Hendrichs, J.; Oritiz, G.; Liedo, P.; Schwarz, A. Six years of successful medfly program in Mexico and Guatemala. In Fruit Flies of Economic Importance: Proceedings of the CEE/IOBC International Symposium: Athens, Netherlands, 16–19 November 1982; Cavalloro, R., Ed.; A. A. Balkema Publishers: Rotterdam, The Netherlands, 1982; pp. 353–365. [Google Scholar]
- Ito, Y.; Kakinohana, H.; Yamagishi, M.; Kohama, T. Eradication of the melon fly, Sactracera cucurbitae, from Okinawa, Japan, by means of the sterile insect technique, with special emphasis on the role of basic studies. J. Asia-Pac. Entomol. 2003, 6, 119–129. [Google Scholar] [CrossRef]
- Koyama, J.; Kakinohana, H.; Miyatake, T. Eradication of the melon fly, Bactrocera cucurbitae, in Japan: Importance of behavior, ecology, genetics, and evolution. Annu. Rev. Entomol. 2004, 49, 331–349. [Google Scholar] [CrossRef]
- Fisher, K. Queensland fruit fly (Bactrocera tryoni): Eradication from western Australia. In Fruit Fly Pests a World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucie Press: St. Lucie, FL, USA, 2020; pp. 535–541. [Google Scholar]
- Proverbs, M.D.; Newton, J.R. Influence of gamma radiation on the development and fertility of the codling moth, Carpocapsa pomonella (L.) (Lepidoptera: Olethreutidae). Can. J. Zool. 1962, 40, 401–419. [Google Scholar] [CrossRef]
- North, D.T. Inherited sterility in Lepidoptera. Annu. Rev. Entomol. 1975, 20, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Aliniazee, M.T.; Long, L.E. Biology and Control of the Cherry Fruit Flies: A Worldwide Perspective; Agricultural Experiment Station, Oregon State University: Corvallis, OR, USA, 1995; pp. 1–110. [Google Scholar]
- Judd, G.J.R.; Gardiner, M.G.T. Towards eradication of codling moth in British Columbia by complimentary actions of mating disruption, tree banding and sterile insect technique: Five-year study in organic orchards. Crop Prot. 2005, 24, 718–733. [Google Scholar] [CrossRef]
- Henneberry, T.J. Integrated systems for control of the pink bollworm Pectinophora gossypiella in Cotton. In Area-Wide Control of Insect Pests From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 567–579. [Google Scholar]
- Tabashnik, B.E.; Liesner, L.R.; Ellsworth, P.C.; Unnithan, G.C.; Fabrick, J.A.; Naranjo, S.E.; Li, X.; Dennehy, T.J.; Antilla, L.; Staten, R.T.; et al. Transgenic cotton and sterile insect releases synergize eradication of pink bollworm a century after it invaded the United States. Proc. Natl. Acad. Sci. USA 2021, 118, e2019115118. [Google Scholar] [CrossRef] [PubMed]
- Lance, D.R.; Mcinnis, D.O. Biological basis of the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 113–142. [Google Scholar]
- Wu, K.M. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar] [CrossRef]
- Li, G.P.; Wu, K.M. Commercial strategy of transgenic insect-resistant maize in China. J. Plant Prot. 2022, 49, 17–32. [Google Scholar] [CrossRef]
- Jiang, S.; He, L.M.; He, W.; Zhao, H.Y.; Yang, X.M.; Yang, X.Q.; Wu, K.M. Effects of X-ray irradiation on the fitness of the established invasive pest fall armyworm Spodoptera frugiperda. Pest. Manag. Sci. 2022, 78, 2806–2815. [Google Scholar] [CrossRef]
- Cayol, J.P.; Vilardi, J.; Rial, E.; Vera, M.T. New indices and method to measure the sexual compatibility and mating performance of Ceratitis capitata (Diptera: Tephritidae) laboratory-reared strains under field cage conditions. J. Econ. Entomol. 1999, 92, 140–145. [Google Scholar] [CrossRef]
- Caceres, C.; Robinson, A.; McInnis, D.; Shelly, T.; Jang, E.; Hendrichs, J. Quality management systems for fruit fly (Diptera: Tephritidae) sterile insect technique. Fla. Entomol. 2007, 90, 1–9. [Google Scholar] [CrossRef]
- Bloem, S.; Bloem, K.A.; Carpenter, J.E.; Calkins, C.O. Inherited sterility in codling moth (Lepidoptera: Tortricidae): Effect of substerilizing doses of radiation on field competitiveness. Environ. Entomol. 1999, 28, 669–674. [Google Scholar] [CrossRef]
- Woods, B.; McInnis, D.; Steiner, E.; Soopaya, A.; Lindsey, J.; Lacey, I.; Virdi, A.; Fogliani, R. Developing field cage tests to measure mating competitiveness of sterile light brown apple moths (Lepidoptera: Tortricidae) in western Australia. Fla. Entomol. 2016, 99, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Barrington, A.M.; Chhagan, A.; Stephens, A.E.A.; Burnip, G.M.; Charles, J.G.; Wee, S.L. Eradication of the Australian painted apple moth Teia anartoides in New Zealand: Trapping, inherited sterility, and male competitiveness. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 603–615. [Google Scholar]
- Firied, M. Determination of sterile insect competitiveness. J. Econ. Entomol. 1971, 64, 869–872. [Google Scholar] [CrossRef]
- Helinski, M.E.H.; Knols, B.G.J. The influence of late-stage pupal irradiation and increased irradiated: Un-irradiated male ratio on mating competitiveness of the malaria mosquito Anopheles arabiensis Patton. Bull. Entomol. Res. 2009, 99, 317–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, J.G.; Aguirre-Ibaniez, S.; Osorio, A.R.; Marina, C.E.; Gomez-Simuta, Y.; Tamayo-Escobar, R.; Dor, A.; Liedo, P.; Carvalho, D.O.; Williams, T. Sexual competitiveness and induced egg sterility by Aedes Aegypti and Aedes albopictus gamma-irradiated males: A laboratory and field study in Mexico. Insects 2021, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–788. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, Y.H.; Liu, H.; Cheng, D.F. A method used for distinguishing between the sexes of Scotogramma trifolii. Chin. J. Appl. Entomol. 2011, 48, 1879–1881. [Google Scholar] [CrossRef]
- Osouli, S.; Ahmadi, M.; Kalantarian, N. Radiation biology and inherited sterility in Helicoverpa armigera Hübner (Lepidoptera: Nuctuidae). Int. J. Trop. Insect Sci. 2021, 41, 2421–2429. [Google Scholar] [CrossRef]
- Mastrangelo, T.; Kovaleski, A.; Botteon, V.; Scopel, W.; Costa, M.D.L.Z. Optimization of the sterilizing doses and overflooding ratios for the South American fruit fly. PLoS ONE 2018, 13, e201026. [Google Scholar] [CrossRef]
- Shelly, T.E.; McInnis, D.O.; Rendon, P. Sterile insect technique and the Mediterranean fruit fly: Assessing the utility of aromatherapy in large field enclosures. Entomol. Exp. Appl. 2005, 116, 199–208. [Google Scholar] [CrossRef]
- Zhou, H.X.; Wang, F.H.; Liu, W.X.; Liu, X.J.; Li, Q. Study on population dynamics and damage of Lygus lucorwm Mayr in transgenie Bt cotton. Chin. J. Eco.-Agric. 2003, 11, 13–15. [Google Scholar]
- Zhang, L.X.; Liu, X.Z. The analysis and application of the two important characteristic points on logistic curve. Hebei J. For. Orch. Res. 1992, 7, 154–158. [Google Scholar]
- Bloem, S.; Carpenter, J.E. Evaluation of population suppression by irradiated lepidoptera and their progeny. Fla. Entomol. 2001, 84, 165–171. [Google Scholar] [CrossRef]
- Simmons, G.S.; Bloem, K.A.; Bloem, S.; Carpenter, J.E.; Suckling, D.M. Impact of moth suppression/eradication programmes using the sterile insect technique or inherited sterility. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 1007–1054. [Google Scholar]
- Hight, S.D.; Carpenter, J.E.; Bloem, S.; Bloem, K.A. Developing a sterile insect release program for Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae): Effective overflooding ratios and release-recapture field studies. Environ. Entomol. 2005, 34, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Steiner, L.F. A method of estimating the size of native populations of oriental, melon, and mediterranean fruit flies to establish the overflooding ratios required for sterile-male releases. J. Econ. Entomol. 1969, 62, 4–7. [Google Scholar] [CrossRef]
- Suckling, D.; Cristofaro, M.; Roselli, G.; Levy, M.; Cemmi, A.; Mazzoni, V.; Stringer, L.; Zeni, V.; Ioriatti, C.; Anfora, G. The competitive mating of irradiated brown marmorated stink bugs, Halyomorpha halys, for the sterile insect technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmeyr, J.H.; Carpenter, J.E.; Bloem, S. Developing the sterile insect technique for Cryptophlebia leucotreta (Lepidoptera: Tortricidae): Influence of radiation dose and release ratio on fruit damage and population growth in field cages. J. Econ. Entomol. 2005, 98, 1924–1929. [Google Scholar] [CrossRef]
- Rendon, P.; McInnis, D.; Lance, D.; Stewart, J. Medfly (Diptera: Tephritidae) genetic sexing: Large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef]
- Meza-Hernandez, J.S.; Diaz-Fleischer, F. Comparison of sexual compatibility between laboratory and wild Mexican fruit flies under laboratory and field conditions. J. Econ. Entomol. 2006, 99, 1979–1986. [Google Scholar] [CrossRef]
- Parker, A.; Mehta, K. Sterile insect technique: A model for dose optimization for improved sterile insect quality. Fla. Entomol. 2007, 90, 88–95. [Google Scholar] [CrossRef]
- Zhang, D.; Zheng, X.; Xi, Z.; Bourtzis, K.; Gilles, J.R.L. Combining the sterile insect technique with the incompatible insect technique: I-impact of Wolbachia infection on the fitness of triple-and double- infected strains of Aedes albopictus. PLoS ONE 2015, 10, e121126. [Google Scholar] [CrossRef] [Green Version]
- Brooks, R.; Kemp, D.J. Can older males deliver the good genes? Trends Ecol. Evol. 2001, 16, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Lingren, P.D.; Warner, W.B.; Henneberry, T.J. Influence of delayed mating on egg production, egg viability, mating, and longevity of female pink bollworm (Lepidoptera: Gelechiidae). Environ. Entomol. 1988, 17, 86–89. [Google Scholar] [CrossRef]
- Milonas, P.G.; Andow, D.A. Virgin male age and mating success in Ostrinia nubilalis (Lepidoptera: Crambidae). Anim. Behav. 2010, 79, 509–514. [Google Scholar] [CrossRef]
- Avent, T.D.; Price, T.A.R.; Wedell, N. Age-based female preference in the fruit fly Drosophila pseudoobscura. Anim. Behav. 2008, 75, 1413–1421. [Google Scholar] [CrossRef]
- Somashekar, K.; Krishna, M.S. Evidence of female preference for older males in Drosophila bipectinata. Zool. Stud. 2011, 50, 1–15. Available online: http://zoolstud.sinica.edu.tw/Journals/50.1/1.pdf (accessed on 20 December 2022).
- Jones, T.M.; Balmford, A.; Quinnell, R.J. Adaptive female choice for middle-aged mates in a lekking sandy. Proc. R. Soc. Ser. B 2000, 267, 681–686. [Google Scholar] [CrossRef]
- Liu, X.P.; Xu, J.; He, H.M.; Kuang, X.J.; Xue, F.S. Male age affects female mate preference and reproductive performance in the cabbage beetle, Colaphellus bowringi. J. Insect Behav. 2011, 24, 83–93. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Diamantidis, A.D.; Nakas, C.T.; Carey, J.R.; Papadopoulos, N.T. Dual reproductive cost of aging in male medflies: Dramatic decrease in mating competitiveness and gradual reduction in mating performance. J. Insect Physiol. 2011, 57, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Shelly, T.E.; Edu, J.; Pahio, E. Female medflies mate selectively with young males but gain no apparent fitness benefits. J. Insect Behav. 2011, 24, 55–66. [Google Scholar] [CrossRef]
- Unnithan, G.C.; Paye, S.O. Mating, longevity, fecundity, and egg fertility of Chilo partellus (Lepidoptera: Pyralidae): Effects of delayed or successive matings and their relevance to pheromonal control methods. Environ. Entomol. 1991, 20, 150–155. [Google Scholar] [CrossRef]
- Lai, M.; Zhang, S.Y.; Zhang, Y.F.; Liu, X.P. Male age affects female mating preference but not fitness in the monandrous moth Dendrolimus punctatus Walker (Lepidoptera: Lasiocampidae). Physiol. Entomol. 2020, 45, 22–29. [Google Scholar] [CrossRef]
- Rogers, C.E.; Marti, O.G. Effects of age at first mating on the reproductive potential of the fall armyworm (Lepidoptera: Noctuidae). Environ. Entomol. 1994, 23, 322–325. [Google Scholar] [CrossRef]
- Kapranas, A.; Collatz, J.; Michaelakis, A.; Milonas, P. Review of the role of sterile insect technique within biologically-based pest control—An appraisal of existing regulatory frameworks. Entomol. Exp. Appl. 2022, 170, 385–393. [Google Scholar] [CrossRef]
- Thistlewood, H.M.A.; Judd, G.J.R. Twenty-five years of research experience with the sterile insect technique and area-wide management of codling moth, Cydia pomonella (L.), in Canada. Insects 2019, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, K.; Banks, D.; Adang, M.J. Toxicity, binding, and permeability analyses of four Bacillus thuringiensis Cry1 delta-endotoxins using brush border membrane vesicles of Spodoptera exigua and Spodoptera frugiperda. Appl. Environ. Microbiol. 1999, 65, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Zhang, L.; Liao, C.; Wu, K.; Xiao, Y. Research progress of resistance mechanism and management techniques of fall armyworm Spodoptera frugiperda to insecticides and Bt crops. Plant Dis. Pests 2019, 10, 10–17. [Google Scholar] [CrossRef]
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V. Managing fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt maize and insecticides in southern Brazil. Pest. Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef]
- Buntin, G.D. Corn expressing Cry1Ab or Cry1F endotoxin for fall armyworm and corn earworm (Lepidoptera: Noctuidae) management in field corn for grain production. Fla. Entomol. 2008, 91, 523–530. [Google Scholar] [CrossRef]
- Bernardi, O.; Amado, D.; Sousa, R.S.; Segatti, F.; Fatoretto, J.; Burd, A.D.; Omoto, C. Baseline susceptibility and monitoring of Brazilian populations of Spodoptera frugiperda (Lepidoptera: Noctuidae) and Diatraea saccharalis (Lepidoptera: Crambidae) to Vip3Aa20 insecticidal protein. J. Econ. Entomol. 2014, 107, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Fatoretto, J.C.; Michel, A.P.; Silva Filho, M.C.; Silva, N. Adaptive potential of fall armyworm (Lepidoptera: Noctuidae) limits Bt trait durability in Brazil. J. Integr. Pest. Manag. 2017, 8, 17. [Google Scholar] [CrossRef]
- Li, G.P.; Ji, T.J.; Zhao, S.Y.; Feng, H.Q.; Wu, K.M. High-dose assessment of transgenic insect-resistant maize events against major lepidopteran pests in China. Plants 2022, 11, 3125. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, D.; Li, D.; Zhao, S.; Wang, C.; Xiao, Y.; Xu, D.; Yang, Y.; Li, G.; Wang, L.; et al. Expression profiles of Cry1Ab protein and its insecticidal efficacy against the invasive fall armyworm for Chinese domestic GM maize DBN9936. J. Integr. Agr. 2021, 20, 792–803. [Google Scholar] [CrossRef]
- Wang, W.H.; Zhang, D.D.; Zhao, S.Y.; Wu, K.M. Susceptibilities of the invasive fall armyworm (Spodoptera frugiperda) to the insecticidal proteins of Bt maize in China. Toxins 2022, 14, 507. [Google Scholar] [CrossRef]
- Ge, S.S.; Chu, B.; He, W.; Jiang, S.; Lv, C.Y.; Gao, L.Y.; Sun, X.T.; Yang, X.M.; Wu, K.M. Wheat-bran-based artificial diet for mass culturing of the fall armyworm, Spodoptera frugiperda Smith (Lepidoptera: Noctuidae). Insects 2022, 13, 1177. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Release Ratio | Hatching Rate of Eggs (%) | Total Number of Eggs | |||||
|---|---|---|---|---|---|---|---|
| T♂:N♂:N♀ 1 | 3 Days after Release | 5 Days after Release | 7 Days after Release | 9 Days after Release | 14 Days after Release | 21 Days after Release | |
| 0:1:1 (blank control) | 2.22 ± 1.34 a 2 | 0.89 ± 0.52 a | 47.37 ± 8.38 a | 76.69 ± 11.83 a | 79.01 ± 10.24 a | 79.01 ± 10.24 a | 5447.00 ± 1306.96 a |
| 1:0:1 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 7.54 ± 3.64 b | 17.82 ± 6.57 b | 18.98 ± 6.60 b | 18.98 ± 6.60 b | 3704.00 ± 760.05 a |
| 12:1:1 | 0.43 ± 0.43 a | 0.30 ± 0.30 a | 20.61 ± 6.13 ab | 29.89 ± 8.91 b | 31.05 ± 9.15 b | 31.05 ± 9.15 b | 4225.50 ± 1620.53 a |
| 16:1:1 | 0.16 ± 0.16 a | 0.05 ± 0.05 a | 19.59 ± 7.40 ab | 39.13 ± 7.54 ab | 40.18 ± 6.94 b | 40.18 ± 6.94 b | 3595.00 ± 927.59 a |
| 20:1:1 | 2.19 ± 1.30 a | 0.80 ± 0.46 a | 14.24 ± 3.23 b | 25.89 ± 6.79 b | 27.57 ± 6.67 b | 27.57 ± 6.67 b | 4246.75 ± 696.65 a |
| 0:1:1 (Insecticide) 3 | 7.01 ± 7.01 a | 3.59 ± 3.50 a | 33.43 ± 8.14 ab | 43.70 ± 6.29 ab | 46.04 ± 6.31 ab | 46.04 ± 6.31 ab | 3172.75 ± 713.26 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Sun, X.-T.; Ge, S.-S.; Yang, X.-M.; Wu, K.-M. Mating Competitiveness of Male Spodoptera frugiperda (Smith) Irradiated by X-rays. Insects 2023, 14, 137. https://doi.org/10.3390/insects14020137
Jiang S, Sun X-T, Ge S-S, Yang X-M, Wu K-M. Mating Competitiveness of Male Spodoptera frugiperda (Smith) Irradiated by X-rays. Insects. 2023; 14(2):137. https://doi.org/10.3390/insects14020137
Chicago/Turabian StyleJiang, Shan, Xiao-Ting Sun, Shi-Shuai Ge, Xian-Ming Yang, and Kong-Ming Wu. 2023. "Mating Competitiveness of Male Spodoptera frugiperda (Smith) Irradiated by X-rays" Insects 14, no. 2: 137. https://doi.org/10.3390/insects14020137
APA StyleJiang, S., Sun, X. -T., Ge, S. -S., Yang, X. -M., & Wu, K. -M. (2023). Mating Competitiveness of Male Spodoptera frugiperda (Smith) Irradiated by X-rays. Insects, 14(2), 137. https://doi.org/10.3390/insects14020137