


Negative Effects of Phthorimaea absoluta-Resistant Tomato Genotypes on the Zoophytophagous Biocontrol Agent, Orius laevigatus (Fieber) (Hemiptera: Anthocoridae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insects
2.3. Measure of O. laevigatus Fitness Components
2.4. Measure of Leaf Trichome Density
2.5. Correlation between O. laevigatus and P. absoluta Performance Variables
2.6. Data Analysis
3. Results
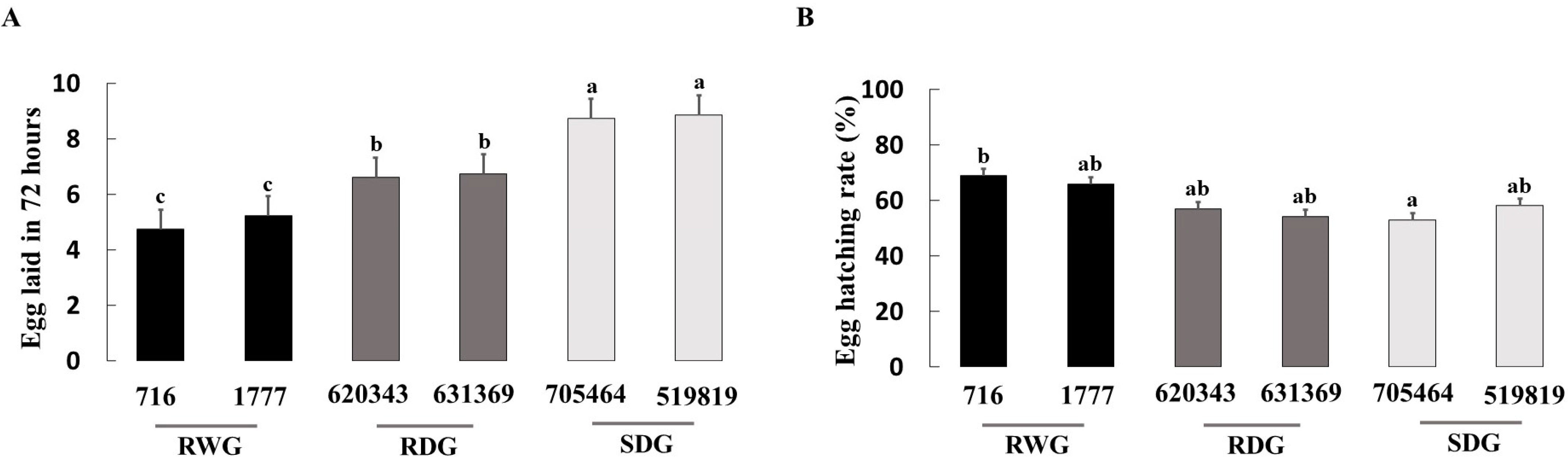
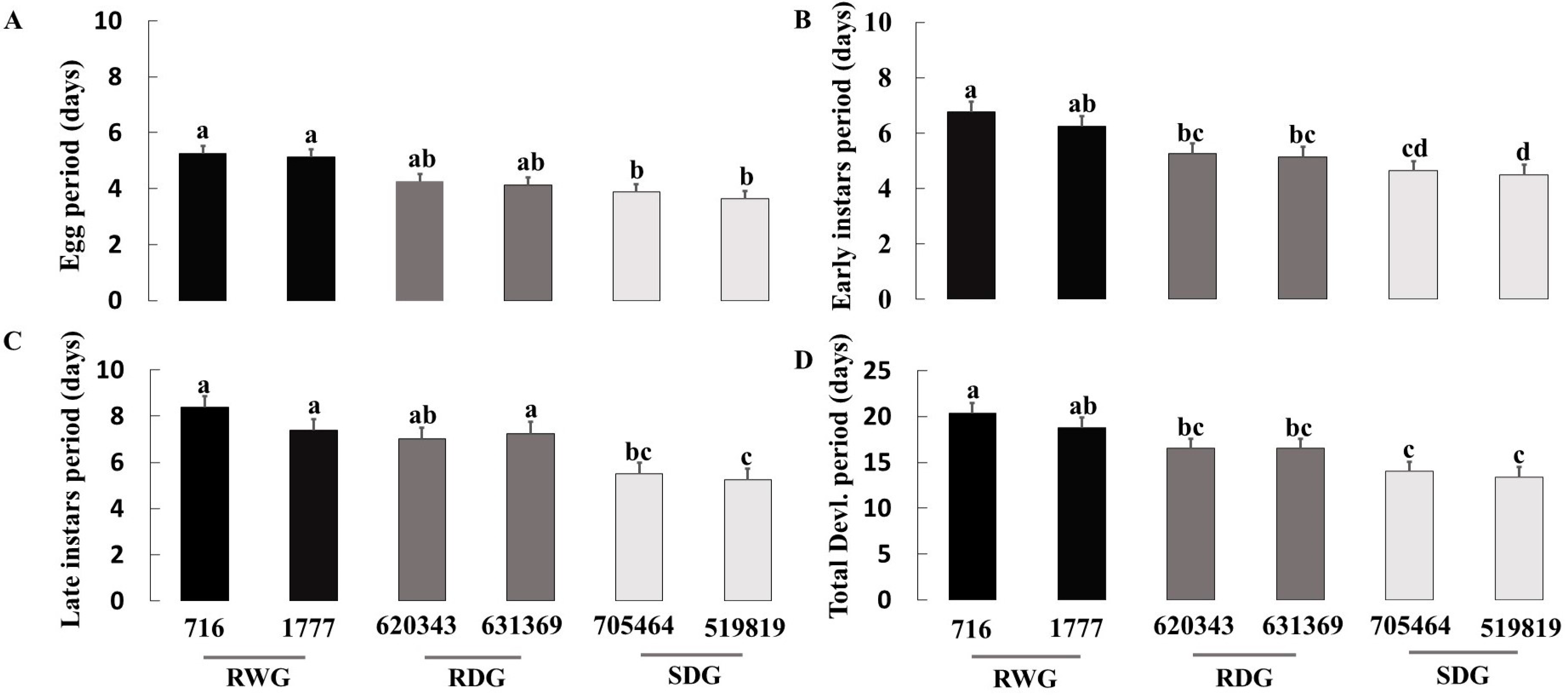
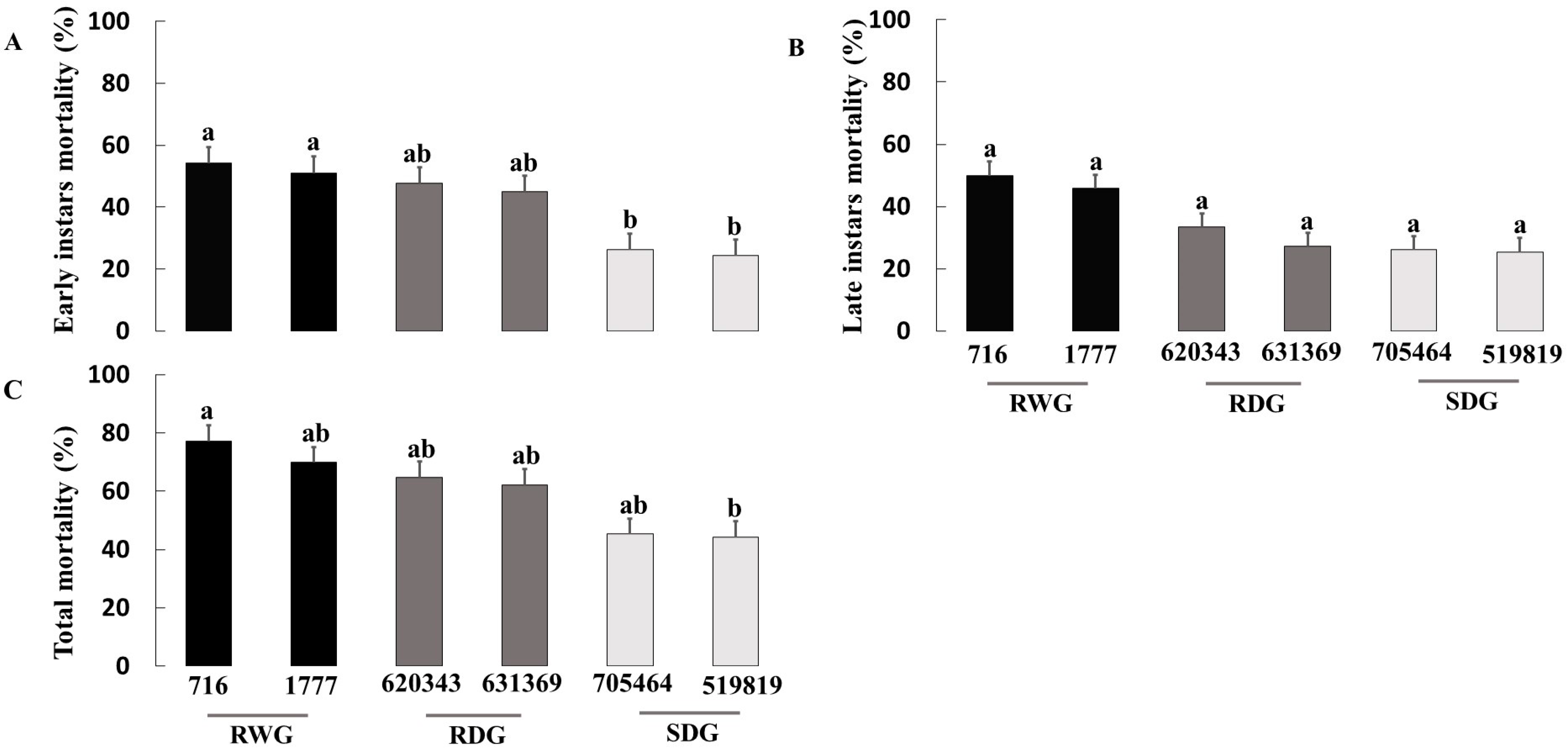
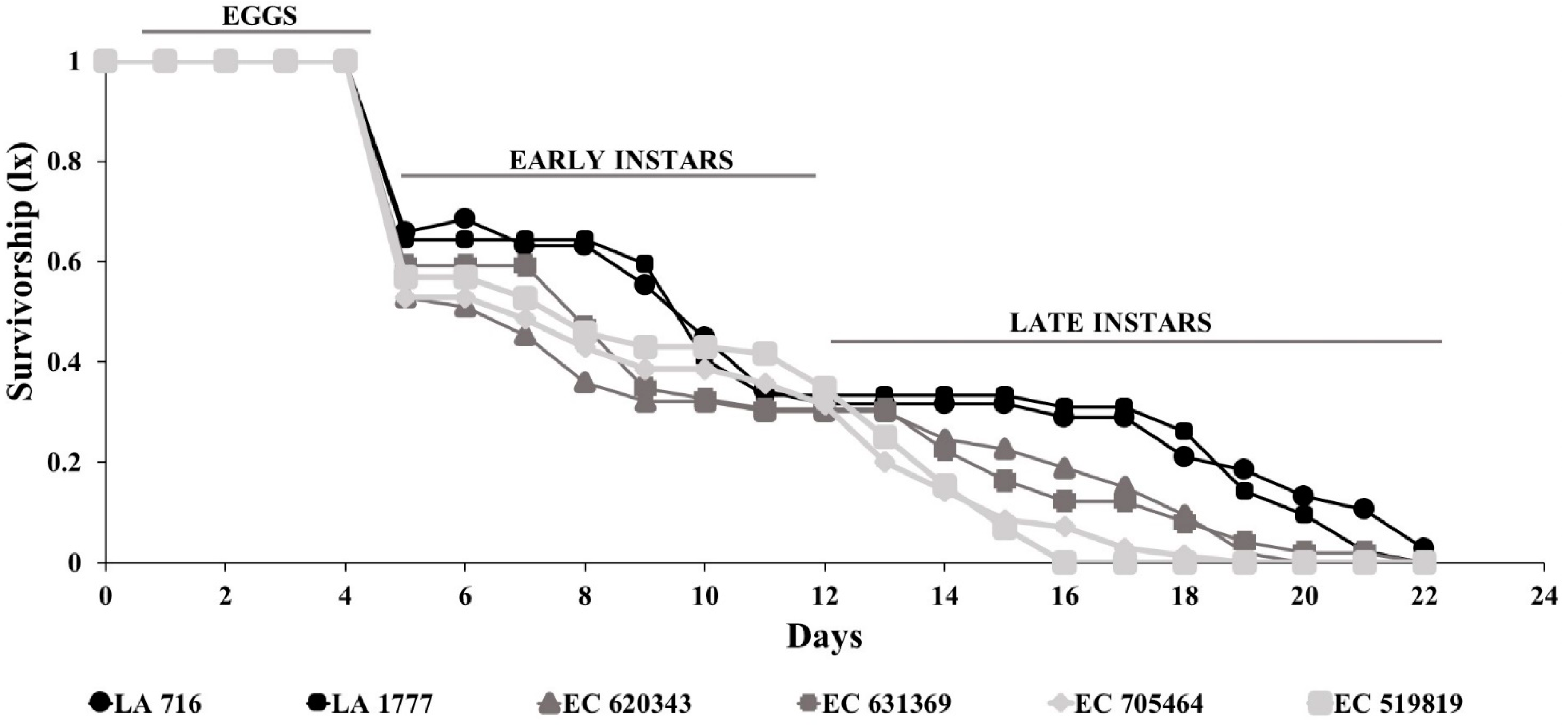
3.1. Measure of O. laevigatus Fitness Components
3.2. Trichome Density
3.3. Effect of Trichome Density on O. laevigatus Survival
3.4. Correlation between O. laevigatus and P. absoluta Performance Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stout, M.J. Host-Plant Resistance in Pest Management. In Integrated Pest Management; Abrol, D.P., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 1–21. [Google Scholar]
- Peterson, J.A.; Ode, P.J.; Oliveira-Hofman, C.; Harwood, J.D. Integration of plant defense traits with biological control of arthropod pests: Challenges and opportunities. Front. Plant. Sci. 2016, 7, 1794. [Google Scholar] [CrossRef] [PubMed]
- Bergman, J.M.; Tingey, W.M. Aspects of interaction between plant genotypes and biological control. Bull. Entomol. Soc. Am. 1979, 25, 275–279. [Google Scholar] [CrossRef]
- Verkerk, R.; Leather, S.; Wright, D.J. The potential for manipulating crop–pest–natural enemy interactions for improved insect pest management. Bull. Entomol. Res. 1998, 88, 493–501. [Google Scholar] [CrossRef]
- Gautam, S.; Mohankumar, S.; Kennedy, J.S. Induced host plant resistance in cauliflower by Beauveria bassiana. J. Entomol. Zool. Stud. 2016, 4, 476–482. [Google Scholar]
- Prabhukarthikeyan, R.; Saravanakumar, D.; Raguchander, T. Combination of endophytic Bacillus and Beauveria for the management of Fusarium wilt and fruit borer in tomato. Pest Manag. Sci. 2014, 70, 1742–1750. [Google Scholar] [CrossRef]
- Groot, A.T.; Dicke, M. Transgenic Crops in an Agro-Ecological Context: Multitrophic Aspects of Insect-Resistant Plants; Ponsen & Looyen: Wageningen, The Netherlands, 2001. [Google Scholar]
- Tandon, P. Negative Aspects of Interaction between Host Plant Resistance and Biological Control and Its Implication in Integrated Pest Management of Crops. In Biocontrol Potential and its Exploitation in Sustainable Agriculture; Upadhyay, R.K., Mukerji, K.G., Chamola, B.P., Eds.; Springer: Boston, MA, USA, 2001; pp. 13–21. [Google Scholar]
- Hare, J.D. Effects of Plant Variation on Herbivore-Natural Enemy Interaction. In Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and Genetics; Fritz, R.S., Simms, E.L., Eds.; The University of Chicago Press: Chicago, IL, USA, 1992; pp. 278–298. [Google Scholar]
- Harvey, J.A.; Van Dam, N.M.; Gols, R. Interactions over four trophic levels: Foodplant quality affects development of a hyperparasitoid as mediated through a herbivore and its primary parasitoid. J. Anim. Ecol. 2003, 72, 520–531. [Google Scholar] [CrossRef]
- Price, P.W.; Bouton, C.E.; Gross, P.; McPheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Annu. Rev. Ecol. Syst. 1980, 11, 41–65. [Google Scholar] [CrossRef]
- Vet, L.E.; Dicke, M. Ecology of infochemical use by natural enemies in a tritrophic context. Annu. Rev. Entomol. 1992, 37, 141–172. [Google Scholar] [CrossRef]
- Dicke, M.; van Poecke, R.M.; de Boer, J.G. Inducible indirect defence of plants: From mechanisms to ecological functions. Basic Appl. Ecol. 2003, 4, 27–42. [Google Scholar] [CrossRef]
- Arimura, G.I.; Kost, C.; Boland, W. Herbivore-induced, indirect plant defences. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids. 2005, 1734, 91–111. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Turlings, T.C. Advances and challenges in the identification of volatiles that mediate interactions among plants and arthropods. Analyst 2006, 131, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Feeny, P. Plant Apparency and Chemical Defense. In Biochemical Interaction between Plants and Insects; Wallace, J.M., Mansell, R.L., Eds.; Springer: Boston, MA, USA, 1976; pp. 1–40. [Google Scholar]
- Thomas, M.; Waage, J. Interaction between Host-Plant Resistance and Biological Control. In Integration of Biological Control and Host Plant Resistance Breeding: A Scientific and Literature Review; Technical Centre for Agricultural and Rural Cooperation: Wageningen, The Netherlands, 1996; pp. 35–50. [Google Scholar]
- Kimura, S.; Sinha, N. Tomato (Solanum lycopersicum): A model fruit-bearing crop. Cold Spring Harb. Protoc. 2008, 11, pdb-emo105. [Google Scholar] [CrossRef]
- Balaji, D.R.; Jeyarani, S.; Ramaraju, K.; Mohankumar, S.; Shanmugam. Occurrence of South American tomato pinworm, Tuta absoluta (Meyrick). (Lepidoptera: Gelechiidae): An invasive pest in Tamil Nadu, India. J. Entomol. Zool. Stud. 2018, 6, 657–662. [Google Scholar]
- Marchant, W.G.; Gautam, S.; Dutta, B.; Srinivasan, R. Whitefly-Mediated Transmission and Subsequent Acquisition of Highly Similar and Naturally Occurring Tomato Yellow Leaf Curl Virus Variants. Phytopathology 2022, 112, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandić, A.; Noris, E.; Matić, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the Mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- Jothi, G.; Pugalendhi, S.; Poornima, K.; Rajendran, G. Management of root-knot nematode in tomato Lycopersicon esculentum, Mill., with biogas slurry. Bioresour. Technol. 2003, 89, 169–170. [Google Scholar] [CrossRef]
- Vargas, C. Observaciones sobre la biologia y enemigos naturales de la polilla del tomate, Gnorimoschema absoluta (Meyrick).(Lep. Gelechiidae). Idesia 1970, 1, 75–110. [Google Scholar]
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches; Springer: Dordrecht, The Netherlands, 2005; pp. 381–412. [Google Scholar]
- Nombela, G.; Muñiz, M. Host plant resistance for the management of Bemisia tabaci: A multi-crop survey with emphasis on tomato. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P., Naranjo, S., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 357–383. [Google Scholar]
- Bottrell, D.G.; Barbosa, P.; Gould, F. Manipulating natural enemies by plant variety selection and modification: A realistic strategy. Annu. Rev. Entomol. 1998, 43, 347–367. [Google Scholar] [CrossRef]
- Pallipparambil, G.R.; Sayler, R.J.; Shapiro, J.P.; Thomas, J.M.; Kring, T.J.; Goggin, F.L. Mi-1.2, an R gene for aphid resistance in tomato, has direct negative effects on a zoophytophagous biocontrol agent, Orius insidiosus. J. Exp. Bot. 2015, 66, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Bottega, D.B.; de Souza, B.H.S.; Rodrigues, N.E.L.; Eduardo, W.I.; Barbosa, J.C.; Júnior, A.L.B. Resistant and susceptible tomato genotypes have direct and indirect effects on Podisus nigrispinus preying on Tuta absoluta larvae. Biol. Control 2017, 106, 27–34. [Google Scholar] [CrossRef]
- Armer, C.A.; Wiedenmann, R.N.; Bush, D.R. Plant feeding site selection on soybean by the facultatively phytophagous predator Orius insidiosus. Entomol. Exp. Appl. 1998, 86, 109–118. [Google Scholar] [CrossRef]
- Eubanks, M.D.; Styrsky, J.D. Effects of plant feeding on the performance of omnivorous predators. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Wackers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, USA, 2005; pp. 148–177. [Google Scholar]
- Coll, M.; Ridgway, R.L. Functional and numerical responses of Orius insidiosus (Heteroptera: Anthocoridae) to its prey in different vegetable crops. Ann. Entomol. Soc. Am. 1995, 88, 732–738. [Google Scholar] [CrossRef]
- Bueno, V.H.; Lins, J.C., Jr.; Silva, D.B.; van Lenteren, J.C. Is predation of Tuta absoluta by three Neotropical mirid predators affected by tomato lines with different densities in glandular trichomes. Arthropod Plant Interact. 2019, 13, 41–48. [Google Scholar] [CrossRef]
- Wheeler, A.G., Jr.; Krimmel, B.A. Mirid (Hemiptera: Heteroptera) specialists of sticky plants: Adaptations, interactions, and ecological implications. Annu. Rev. Entomol. 2015, 60, 393–414. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, X.; Shen, H.; Xue, H.; Tian, T.; Zhang, Q.; Hu, J.; Tong, H.; Zhang, Y.; Su, Q. Flavonoid-producing tomato plants have a direct negative effect on the zoophytophagous biological control agent Orius sauteri. Insect Sci. 2022, 29, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Van Lenteren, J.C.; Alomar, O.; Ravensberg, W.J.; Urbaneja, A. Biological Control Agents for Control of Pests in Greenhouses. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer: Cham, Switzerland, 2020; Volume 9, pp. 409–439. [Google Scholar]
- Tan, X.-L.; Wang, S.; Liu, T.X. Acceptance and suitability of four plant substrates for rearing Orius sauteri (Hemiptera: Anthocoridae). Biocontrol Sci. Technol. 2014, 24, 291–302. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Z.; Liu, P.; Yin, Y.; Felton, G.W.; Shi, W.J. Influence of plant physical and anatomical characteristics on the ovipositional preference of Orius sauteri (Hemiptera: Anthocoridae). Insects 2021, 12, 326. [Google Scholar] [CrossRef]
- Guruswamy, M.; Marimuthu, M.; Coll, M. Life table analyses for the Tomato Leaf Miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae): Effects of plant genotype. Pest Manag. Sci. 2023. In press. [Google Scholar] [CrossRef]
- Pehlivan, S. Influence of the eggs of Ephestia kuehniella (Lepidoptera: Pyralidae) reared on different diets on the performance of the predatory bug Orius laevigatus (Hemiptera: Anthocoridae). Eur. J. Entomol. 2021, 118, 51–56. [Google Scholar] [CrossRef]
- De Clercq, P.; Coudron, T.A.; Riddick, E.W.; Morales-Ramos, J.A.; Rojas, G.; Shapiro, D.I. Production of Heteropteran Predators. In Mass Production of Beneficial Organisms; Morales-Ramos, J.A., Rojas, M.G., Shapiro-Ilan, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 57–100. [Google Scholar]
- Maiti, R.; Bidinger, F.; Reddy, K.S.; Gibson, P.; Davis, J. Nature and Occurrence of Trichomes in Sorghum Lines with Resistance to the Sorghum Shootfly; RP 01485, 1980; International Crops Research Institute for the Semi-Arid Tropics: Hyderabad, India, 1980; pp. 1–36. [Google Scholar]
- Inbar, M.; Gerling, D. Plant-mediated interactions between whiteflies, herbivores, and natural enemies. Annu. Rev. Entomol. 2008, 53, 431–448. [Google Scholar] [CrossRef]
- Styrsky, J.D.; Kaplan, I.; Eubanks, M.D. Plant trichomes indirectly enhance tritrophic interactions involving a generalist predator, the red imported fire ant. Biol. Control 2006, 36, 375–384. [Google Scholar] [CrossRef]
- Kennedy, G.G. Tomato, pests, parasitoids, and predators: Tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 2003, 48, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Riddick, E.W.; Simmons, A.M. Do plant trichomes cause more harm than good to predatory insects. Pest Manage. Sci. 2014, 70, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Paspati, A.; Rambla, J.L.; Gresa, M.P.L.; Arbona, V.; Gómez-Cadenas, A.; Granell, A.; González-Cabrera, J.; Urbaneja, A. Tomato trichomes are deadly hurdles limiting the establishment of Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Biol. Control 2021, 157, 104572. [Google Scholar] [CrossRef]
- Tian, D.; Tooker, J.; Peiffer, M.; Chung, S.H.; Felton, G.W. Role of trichomes in defense against herbivores: Comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 2012, 236, 1053–1066. [Google Scholar] [CrossRef]
- Lucini, T.; Faria, M.V.; Rohde, C.; Resende, J.T.V.; de Oliveira, J.R.F. Acylsugar and the role of trichomes in tomato genotypes resistance to Tetranychus urticae. Arthropod Plant Interact. 2015, 9, 45–53. [Google Scholar] [CrossRef]
- Bitew, M.K. Significant role of wild genotypes of tomato trichomes for Tuta absoluta resistance. J. Plant Genet. Breed. 2018, 2, 104. [Google Scholar]
- Simmons, A.T.; Gurr, G.M. Trichomes of Lycopersicon species and their hybrids: Effects on pests and natural enemies. Agric. For. Entomol. 2005, 7, 265–276. [Google Scholar] [CrossRef]
- Simmons, A.T.; Gurr, G.M. The effect on the biological control agent Mallada signata of trichomes of F1 Lycopersicon esculentum × L. cheesmanii f. minor and L. esculentum × L. pennellii hybrids. Biol. Control 2006, 38, 174–178. [Google Scholar] [CrossRef]
- Maiti, R.; Satya, P.; Rajkumar, D.; Ranaswamy, A. Anatomical changes in crops under adaptation. In Crop Plant Anatomy; Cabi: Wallingford, UK, 2012; p. 269. [Google Scholar]
- Coll, M. Feeding and ovipositing on plants by an omnivorous insect predator. Oecologia 1996, 105, 214–220. [Google Scholar] [CrossRef]
- Barber, G.W. Orius Insidiosus (Say), an Important Natural Enemy of the Corn Ear Worm; US Department of Agriculture: Washington, DC, USA, 1936.
- Agrawal, A.A.; Klein, C.N. What omnivores eat: Direct effects of induced plant resistance on herbivores and indirect consequences for diet selection by omnivores. J. Anim. Ecol. 2000, 69, 525–535. [Google Scholar] [CrossRef]
- Perdikis, D.; Lykouressis, D. Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 2000, 17, 55–60. [Google Scholar] [CrossRef]
- Lee, J.C.; Heimpel, G.E. Floral resources impact longevity and oviposition rate of a parasitoid in the field. J. Anim. Ecol. 2008, 77, 565–572. [Google Scholar] [CrossRef]
- Nomikou, M.; Janssen, A.; Schraag, R.; Sabelis, M.W. Phytoseiid predators suppress populations of Bemisia Tabaci on cucumber plants with alternative food. Exp. Appl. Acarol. 2002, 27, 57–68. [Google Scholar] [CrossRef]
- Perdikis, D.C.; Lykouressis, D.P. Life table and biological characteristics of Macrolophus Pygmaeus when feeding on Myzus Persicae and Trialeurodes Vaporariorum. Entomol. Exp. Appl. 2002, 102, 261–272. [Google Scholar] [CrossRef]
- Eubanks, M.D.; Denno, R.F. The ecological consequences of variation in plants and prey for an omnivorous insect. Ecology 1999, 80, 1253–1266. [Google Scholar] [CrossRef]
- Wanner, H.; Gu, H.; Dorn, S. Nutritional value of floral nectar sources for flight in the parasitoid wasp, Cotesia Glomerata. Physiol. Entomol. 2006, 31, 127–133. [Google Scholar] [CrossRef]
- Snyder, W.E.; Wise, D.H. Contrasting trophic cascades generated by a community of generalist predators. Ecology 2001, 82, 1571–1583. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Intraguild predation diminished in complex-structured vegetation: Implications for prey suppression. Ecology 2002, 83, 643–652. [Google Scholar] [CrossRef]
- Bruno, J.F.; O’Connor, M.I. Cascading effects of predator diversity and omnivory in a marine food web. Ecol. Lett. 2005, 8, 1048–1056. [Google Scholar] [CrossRef]
- Eubanks, M.D.; Denno, R.F. Host plants mediate omnivore–herbivore interactions and influence prey suppression. Ecology 2000, 81, 936–947. [Google Scholar]
- Janssen, A.; Willemse, E.; Van Der Hammen, T. Poor host plant quality causes omnivore to consume predator eggs. J. Anim. Ecol. 2003, 72, 478–483. [Google Scholar] [CrossRef]
- Badii, M.H.; Hernández-Ortiz, E.; Flores, A.E.; Landeros, J.A. Prey stage preference and functional response of Euseius Hibisci to Tetranychus Urticae (Acari: Phytoseiidae, Tetranychidae). Exp. Appl. Acarol. 2004, 34, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Spellman, B.; Brown, M.; Mathews, C.R. Effect of floral and extrafloral resources on predation of Aphis spiraecola by Harmonia axyridis on apple. BioControl 2006, 51, 715–724. [Google Scholar] [CrossRef]
- Robinson, K.A.; Jonsson, M.; Wratten, S.D.; Wade, M.R.; Buckley, H.L. Implications of floral resources for predation by an omnivorous lacewing. Basic Appl. Ecol. 2008, 9, 172–181. [Google Scholar] [CrossRef]
- Coll, M. Feeding on Non-Prey Resources by Natural Enemies. In Relationships of Natural Enemies and Non-Prey Foods; Lundgren, J.G., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 9–23. [Google Scholar]
- Gillespie, D.; McGregor, R.R. The functions of plant feeding in the omnivorous predator Dicyphus hesperus: Water places limits on predation. Ecol. Entomol. 2000, 25, 380–386. [Google Scholar] [CrossRef]
- Ågren, G.I.; Stenberg, J.A.; Björkman, C. Omnivores as plant bodyguards—A model of the importance of plant quality. Basic Appl. Ecol. 2012, 13, 441–448. [Google Scholar] [CrossRef]
- van Emden, H.F. Host plant-aphidophaga interactions. Agric. Ecosys. Environ. 1995, 52, 3–11. [Google Scholar] [CrossRef]
- Shannag, H.; Obeidat, W.M. Interaction between plant resistance and predation of Aphis fabae (Homoptera: Aphididae) by Coccinella septempunctata (Coleoptera: Coccinellidae). Ann. Appl. Biol. 2008, 152, 331–337. [Google Scholar] [CrossRef]
- Bartlett, R. Negative interactions between chemical resistance and predators affect fitness in soybeans. Ecol. Entomol. 2008, 33, 673–678. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | r Value | p Value |
|---|---|---|
| Glandular trichome vs. total mortality of O. laevigatus | 0.47 | 0.003 |
| Total trichomes vs. total mortality of O. laevigatus | 0.49 | <0.001 |
| Total trichomes vs. early larval mortality of O. laevigatus | 0.55 | <0.001 |
| Glandular trichome vs. early larval mortality of O. laevigatus | 0.43 | 0.002 |
| Parameters | r Value | p Value |
|---|---|---|
| Egg period of P. absoluta vs. egg period of O. laevigatus | 0.96 | 0.003 |
| Development time of early stages of P. absoluta vs. development time of early stages of O. laevigatus | 0.89 | 0.019 |
| Development time of late stages of P. absoluta vs. development time of late stages of O. laevigatus | 0.92 | 0.011 |
| Total mortality of P. absoluta vs. total mortality of O. laevigatus | 0.86 | 0.030 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guruswamy, M.; Marimuthu, M.; Coll, M. Negative Effects of Phthorimaea absoluta-Resistant Tomato Genotypes on the Zoophytophagous Biocontrol Agent, Orius laevigatus (Fieber) (Hemiptera: Anthocoridae). Insects 2023, 14, 160. https://doi.org/10.3390/insects14020160
Guruswamy M, Marimuthu M, Coll M. Negative Effects of Phthorimaea absoluta-Resistant Tomato Genotypes on the Zoophytophagous Biocontrol Agent, Orius laevigatus (Fieber) (Hemiptera: Anthocoridae). Insects. 2023; 14(2):160. https://doi.org/10.3390/insects14020160
Chicago/Turabian StyleGuruswamy, Megha, Murugan Marimuthu, and Moshe Coll. 2023. "Negative Effects of Phthorimaea absoluta-Resistant Tomato Genotypes on the Zoophytophagous Biocontrol Agent, Orius laevigatus (Fieber) (Hemiptera: Anthocoridae)" Insects 14, no. 2: 160. https://doi.org/10.3390/insects14020160
APA StyleGuruswamy, M., Marimuthu, M., & Coll, M. (2023). Negative Effects of Phthorimaea absoluta-Resistant Tomato Genotypes on the Zoophytophagous Biocontrol Agent, Orius laevigatus (Fieber) (Hemiptera: Anthocoridae). Insects, 14(2), 160. https://doi.org/10.3390/insects14020160