
An Acyl Carrier Protein Gene Affects Fatty Acid Synthesis and Growth of Hermetia illucens


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Sample Collection
2.2. Cloning of HiACP cDNAs
2.3. Sequence Analysis and Quantification of the HiACP Gene
2.4. RNAi Treatment
2.5. Fatty Acid Contents in H. illucens
2.6. Statistical Analysis
3. Results
3.1. Sequence Analysis of HiACP
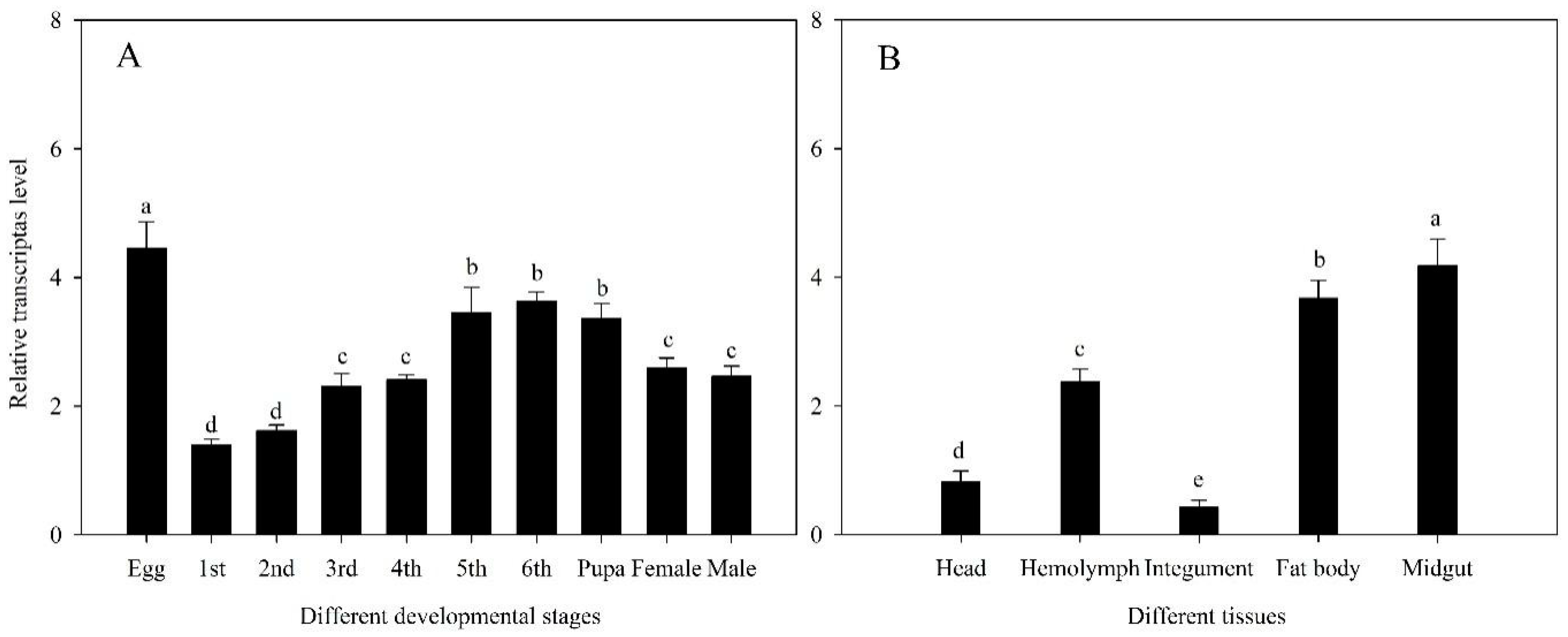
3.2. Expression at Growth Stages and Tissues
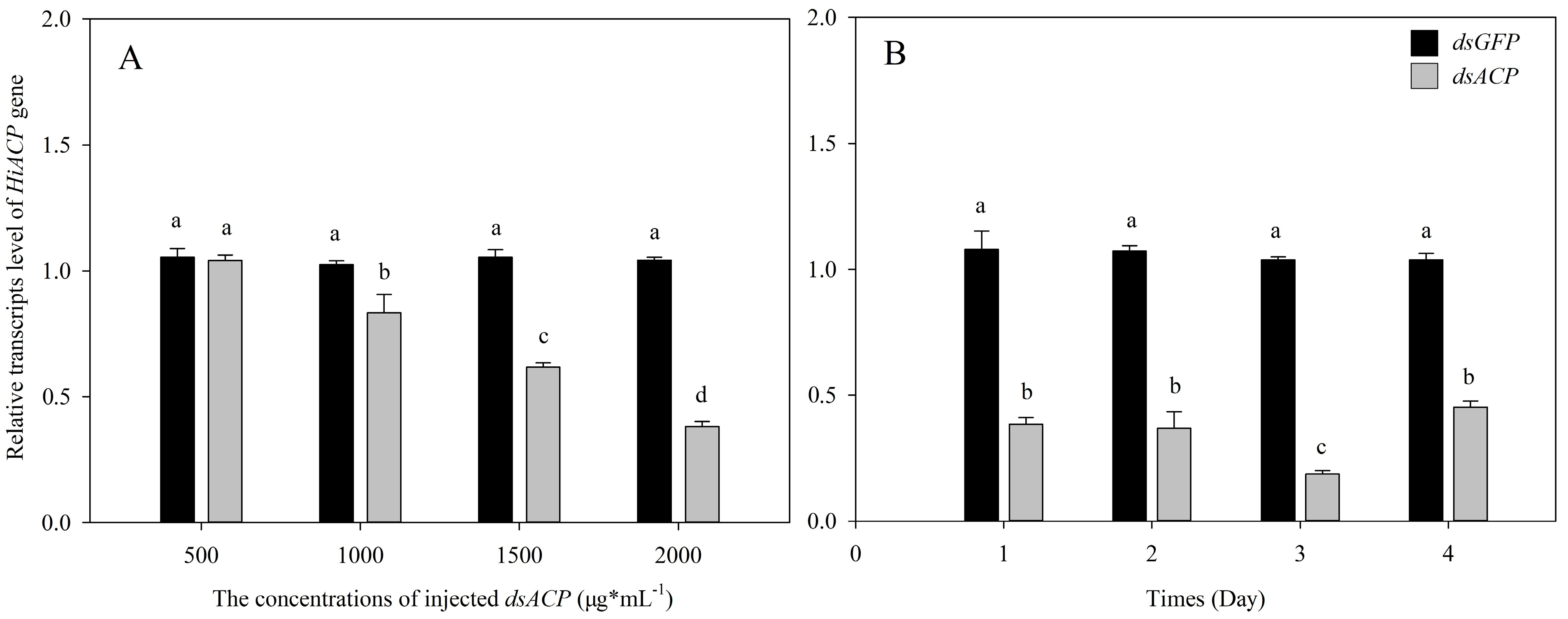
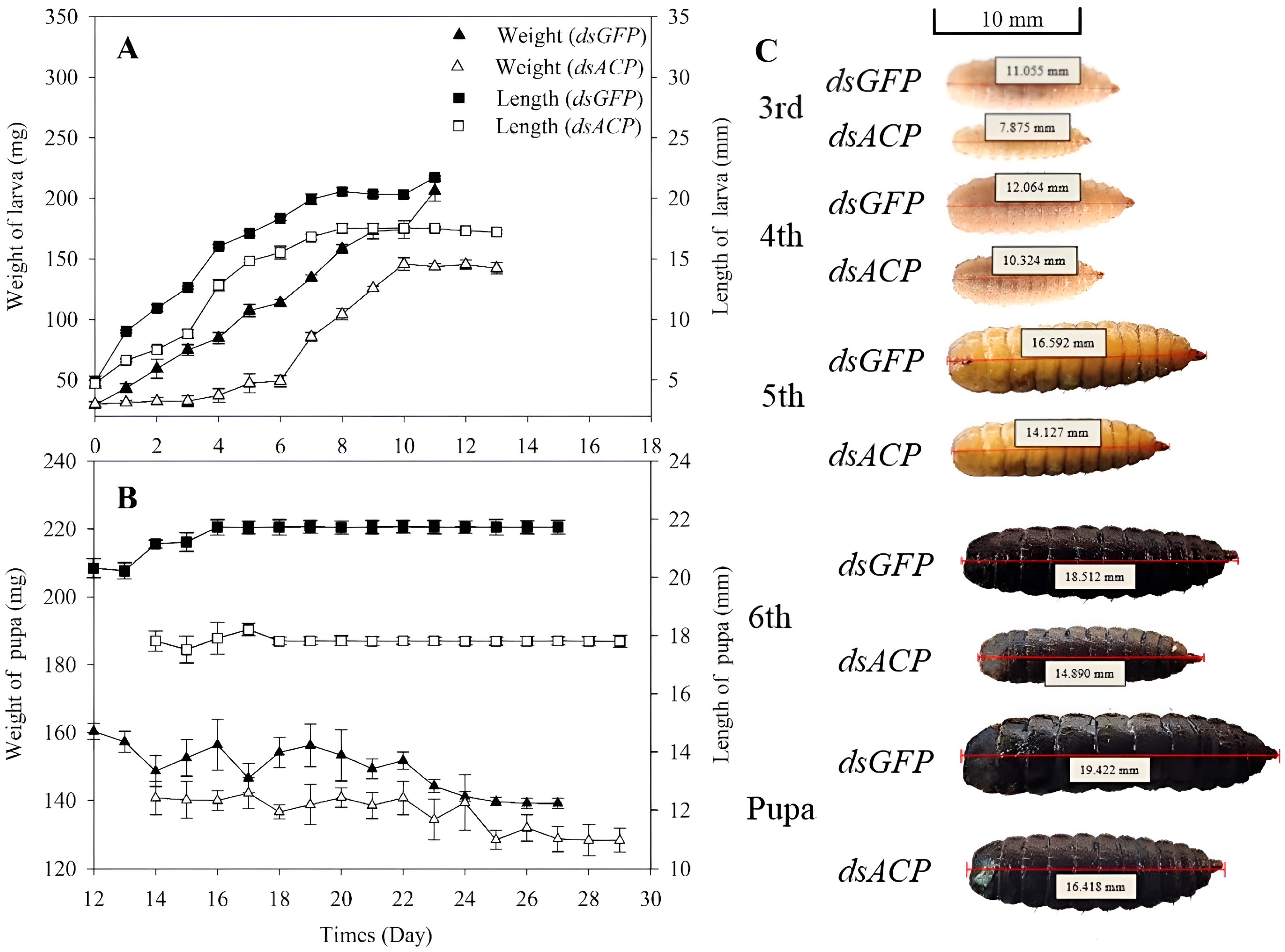
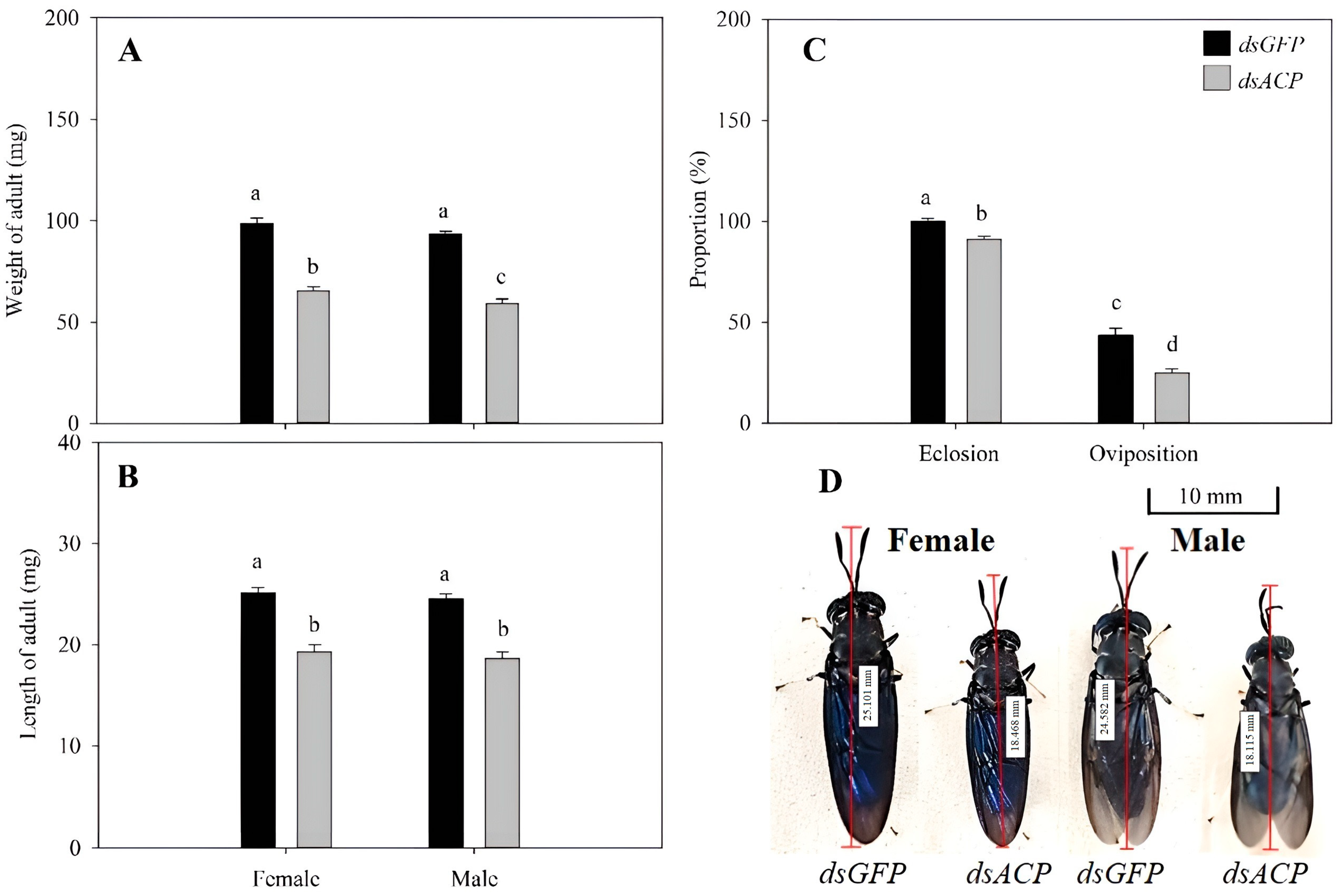
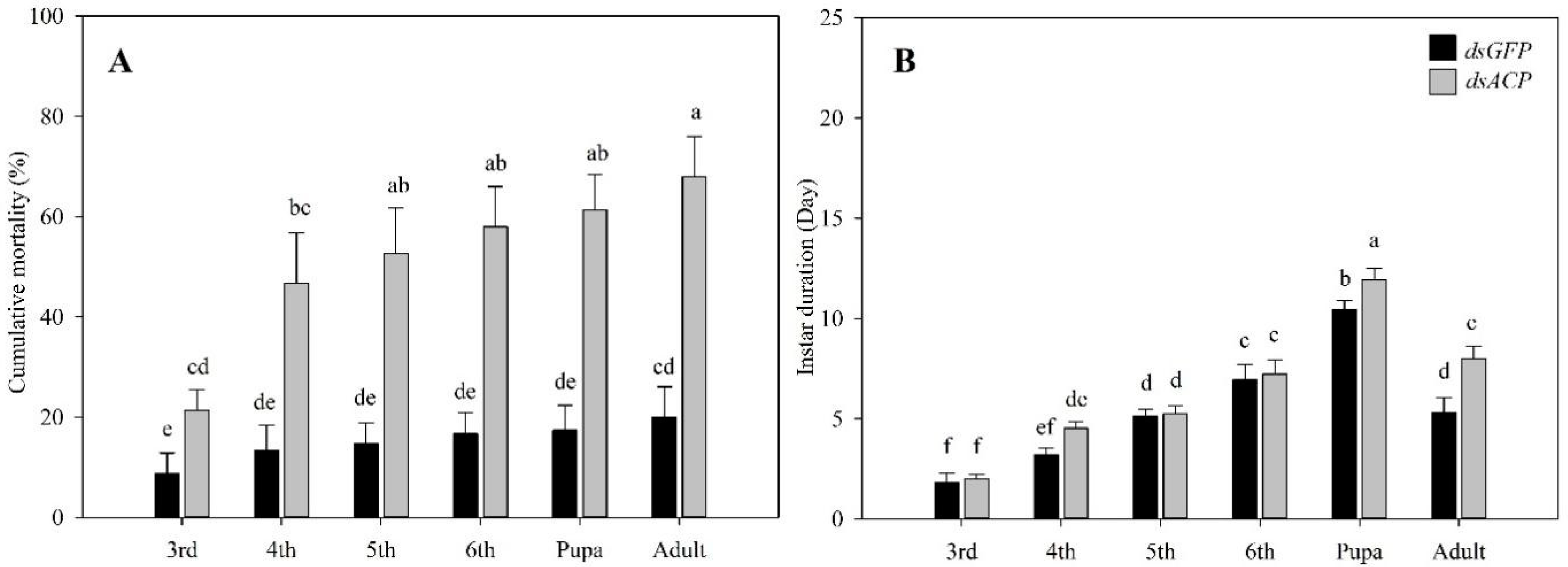
3.3. Effects of RNAi of HiACP
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G.; et al. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia-Pac. Entomol. 2017, 20, 2–38. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids (PLEFA) 2018, 132, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerkowniak, M.; Puckowski, A.; Stepnowski, P.; Gołębiowski, M. The use of chromatographic techniques for the separation and the identification of insect lipids. J. Chromatogr. B 2013, 35, 869–937. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.A.; Oonincx, B.; Laurent, S.; Margot, E.; Veenenbos, J.A. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Sci. 2020, 27, 3–22. [Google Scholar]
- Suh, J.M.; Gao, X.; McKay, J.; McKay, R.; Salo, Z.; Graff, J.M. Hedgehog signaling plays a conserved role in inhibiting fat formation. Cell Metab. 2006, 3, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.I.; Vogel, H.J. Current understanding of fatty acid biosynthesis and the acyl carrier protein. Biochem. J. 2010, 430, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Sherman, D.H. An Enzyme Assembly Line. Science 2008, 321, 1304–1305. [Google Scholar] [CrossRef]
- Huang, J.; Xue, C.; Wang, H.; Wang, L.; Schmidt, W.; Shen, R.; Lan, P. Genes of acyl carrier protein Family Show Different Expression Profiles and Overexpression of acyl carrier protein 5 Modulates Fatty Acid Composition and Enhances Salt Stress Tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 987–988. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hicks, W.M.; Silver, P.A.; Way, J.C. Engineering acyl carrier protein to enhance production of shortened fatty acids. Biotechnol. Biofuels 2016, 9, 24–25. [Google Scholar] [CrossRef] [Green Version]
- Cronan, J.E. The chain-flipping mechanism of ACP (acyl carrier protein)-dependent enzymes appears universal. Biochem. J. 2014, 460, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Sulpizio, A.; Crawford, C.E.; Koweek, R.S.; Charkoudian, L.K. Probing the structure and function of acyl carrier proteins to unlock the strategic redesign of type II polyketide biosynthetic pathways. J. Biol. Chem. 2021, 296, 100328. [Google Scholar] [CrossRef]
- Mofid, M.R.; Finking, R.; Marahiel, M.A. Recognition of hybrid peptidyl carrier proteins/acyl carrier proteins in nonribosomal peptide synthetase modules by the 4’-phosphopantetheinyl transferases AcpS and Sfp. J. Biol. Chem. 2002, 277, 17023–17031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, E.; Hofmann, J. Microbial Type I Fatty Acid Synthases (FAS): Major Players in a Network of Cellular FAS Systems. Microbiol. Mol. Biol. 2004, 68, 501–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunphy, K.Y.; Senaratne, R.H.; Masuzawa, M.; Kendall, L.V.; Riley, L.W. Attenuation of Mycobacterium tuberculosis Functionally Disrupted in a Fatty Acyl–Coenzyme A Synthetase Gene fadD5. J. Infect. Dis. 2010, 201, 1232–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrechts, R.A.; Schepers, H.; Yu, Y.; van der Zwaag, M.; Autio, K.J.; Vieira-Lara, M.A.; Bakker, B.M.; Tijssen, M.A.; Hayflick, S.J.; Grzeschik, N.A.; et al. CoA-dependent activation of mitochondrial acyl carrier protein links four neurodegenerative diseases. EMBO Mol. Med. 2019, 11, 12–15. [Google Scholar] [CrossRef]
- Ragone, G.; Caizzi, R.; Moschetti, R. The Drosophila melanogaster gene for the NADH: Ubiquinone oxidoreductase acyl carrier protein: Developmental expression analysis and evidence for alternatively spliced forms. Mol. Gen. Genet. 1999, 261, 4–5. [Google Scholar] [CrossRef]
- Clements, J.; Olson, J.M.; Sanchez-Sedillo, B.; Bradford, B.; Groves, R.L. Changes in emergence phenology, fatty acid composition, and xenobiotic-metabolizing enzyme expression is associated with increased insecticide resistance in the Colorado potato beetle. Arch. Insect Biochem. Physiol. 2020, 103, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.A.; Ristow, B.; Rahayu, T.; Putra, N.S.; Yuwono, N.W.; Mategeko, B.; Smetana, S.; Saki, M.; Nawaz, A.; Nagdalian, A. Black soldier fly larvae (BSFL) and their affinity for organic waste processing. Waste Manag. 2022, 140, 1–13. [Google Scholar] [CrossRef]
- Van Huis, A.; Gasco, L. Insects as feed for livestock production. Science 2023, 379, 138–139. [Google Scholar] [CrossRef]
- Liew, C.S.; Mong, G.R.; Abdelfattah, E.A.; Raksasat, R.; Rawindran, H.; Kiatkittipong, W.; Mohamad, M.; Ramli, A.; Yunus, N.M.; Lam, M.K.; et al. Correlating black soldier fly larvae growths with soluble nutrients derived from thermally pre-treated waste activated sludge. Environ. Res. 2022, 210, 112923. [Google Scholar] [CrossRef] [PubMed]
- Naser El Deen, S.; van Rozen, K.; Elissen, H.; van Wikselaar, P.; Fodor, I.; van der Weide, R.; Hoek-van den Hil, E.F.; Rezaei Far, A.; Veldkamp, T. Bioconversion of Different Waste Streams of Animal and Vegetal Origin and Manure by Black Soldier Fly Larvae Hermetia illucens L.(Diptera: Stratiomyidae). Insects 2023, 14, 204. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.U.; Hollah, C.; Wiesotzki, K.; Rehman, R.U.; Zhang, J.; Zheng, L.; Nienaber, T.; Heinz, V.; Aganovic, K. Black soldier fly, Hermetia illucens as a potential innovative and environmentally friendly tool for organic waste management: A mini-review. Waste Manag. Res. 2023, 41, 81–97. [Google Scholar] [CrossRef]
- Romano, N.; Sinha, A.; Powell, A.; Fischer, H. Mineral composition in black soldier fly (Hermetia illucens) larvae and resulting frass from fruit and their peels. J. Insects Food Feed 2023, 9, 43–53. [Google Scholar] [CrossRef]
- Cheng, V.; Shoveller, A.K.; Huber, L.-A.; Kiarie, E.G. Comparative protein quality in black soldier fly larvae meal vs. soybean meal and fish meal using classical protein efficiency ratio (PER) chick growth assay model. Poult. Sci. 2023, 102, 102255. [Google Scholar]
- Shah, A.A.; Wanapat, M. Gryllus testaceus walker (crickets) farming management, chemical composition, nutritive profile, and their effect on animal digestibility. Entomol. Res. 2021, 51, 639–649. [Google Scholar] [CrossRef]
- Shah, A.A.; Totakul, P.; Matra, M.; Cherdthong, A.; Harnboonsong, Y.; Wanapat, M. Nutritional composition of various insects and potential uses as alternative protein sources in animal diets. Anim. Biosci. 2022, 35, 317. [Google Scholar] [CrossRef]
- Boykin, K.; Mitchell, M.A. The Value of Black Soldier Fly Larvae (Hermetia illucens) as a Food Source: A Review. J. Herpetol. Med. Surg. 2021, 31, 3–11. [Google Scholar] [CrossRef]
- Anang, D.M.; Rusul, G.; Bakar, J.; Ling, F.H. Effects of lactic acid and lauricidin on the survival of Listeria monocytogenes, Salmonella enteritidis and Escherichia coli O157: H7 in chicken breast stored at 4 °C. Food Control 2006, 18, 8–24. [Google Scholar] [CrossRef]
- Lawal, K.G.; Kavle, R.R.; Akanbi, T.O.; Mirosa, M.; Agyei, D. Enrichment in specific fatty acids profile of Tenebrio molitor and Hermetia illucens larvae through feeding. Future Foods 2021, 3, 4–6. [Google Scholar] [CrossRef]
- Danieli, P.P.; Lussiana, C.; Gasco, L.; Amici, A.; Ronchi, B. The Effects of Diet Formulation on the Yield, Proximate Composition, and Fatty Acid Profile of the Black Soldier Fly (Hermetia illucens L.) Prepupae Intended for Animal Feed. Animals 2019, 9, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, O.M. Dried Hermetia illucens larvae (Diptera: Stratiomy-idae) as a feed additive for poultry. J. Ga. Entomol. Soc. 1973, 173, 16–20. [Google Scholar]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)–Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Boyen, F.; Bohez, L.; Pasmans, F.; Volf, J.; Sevcik, M.; Rychlik, I.; Haesebrouck, F.; Ducatelle, R. Medium-chain fatty acids decrease colonization and invasion through HILA suppression shortly after infection of chickens with Salmonella enterica serovar Enteritidis. Appl. Environ. Microbiol. 2004, 70, 3582–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alagappan, S.; Hoffman, L.C.; Mantilla, S.M.O.; Mikkelsen, D.; James, P.; Yarger, O.; Cozzolino, D. Near Infrared Spectroscopy as a Traceability Tool to Monitor Black Soldier Fly Larvae (Hermetia illucens) Intended as Animal Feed. Appl. Sci. 2022, 12, 8168. [Google Scholar] [CrossRef]
- Barrera, J.; Salinas, E.R.; Rivera, A.R.; Malalis, J.S.; Bagaforo, E.R.O.; Araya, J.P.; Sulay, D.L.D. Productivity and Nutritive Value of Black Soldier Fly (Hermetia illucens) Larvae Fed with Different Waste Substrates. Int. J. Multidiscip. Appl. Bus. Educ. Res. 2023, 4, 588–599. [Google Scholar] [CrossRef]
- De Matteis, V.; Cascione, M.; De Giorgi, M.L.; Leporatti, S.; Rinaldi, R. Encapsulation of Thermo-Sensitive Lauric Acid in Silica Shell: A Green Derivate for Chemo-Thermal Therapy in Breast Cancer Cell. Molecules 2019, 24, 2034. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 146–176. [Google Scholar] [CrossRef]
- Mariam, Q.; David, T.L.; Conrad, S. METAGENOTE: A simplified web platform for metadata annotation of genomic samples and streamlined submission to NCBI’s sequence read archive. BMC Bioinform. 2020, 21, 55–67. [Google Scholar]
- Gasteiger, E. Protein Identifcation and Analysis Tools on the ExPASy Server. Proteom. Protoc. Handb. 2005, 8, 571–607. [Google Scholar]
- Chou, K.C.; Shen, H.B. Euk-mPLoc: A fusion classifer for large-scale eukaryotic protein subcellular location prediction by incorporating multiple sites. J. Proteome Res. 2007, 6, 1728–1734. [Google Scholar] [CrossRef] [PubMed]
- Irina, V.U.; Marina, A.K.; Ilya, G.K. New insights into the organisation of the oxidative phosphorylation system in the example of pea shoot mitochondria. Biochim. Biophys. Acta (BBA)-Bioenerg. 2020, 1861, 380–386. [Google Scholar]
- Janice, L.M.; Anne, K.G.; Mary, W.M. Use of SOMPA in Identification of Gifted African-American Children. J. Educ. Gift. 1992, 15, 13–21. [Google Scholar]
- Rebecca, A.H.; Janet, S.D.; Beverly, S.C. Prolactin induces Jak2 phosphorylation of RUSHY195. Mol. Cell. Endocrinol. 2011, 338, 129–149. [Google Scholar]
- Haghighi, M.; Khanahmad, H.; Palizban, A. Selection and Characterization of Single-Stranded DNA Aptamers Binding Human B-Cell Surface Protein CD20 by Cell-SELEX. Molecules 2018, 23, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanyan, Z.; Ning, S.; Fei, L. Activation of mitogen-activated protein kinases in satellite glial cells of the trigeminal ganglion contributes to substance P-mediated inflammatory pain. Int. J. Oral Sci. 2019, 11, 213–222. [Google Scholar]
- Mazumdar, J.; Wilson, E.; Masek, K. Apicoplast fatty acid synthesis is essential for organelle biogenesis and parasite survival in Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2006, 103, 13192–13197. [Google Scholar] [CrossRef] [Green Version]
- Manchester, K.L. Value of A260/A280 ratios for measurement of purity of nucleic acids. BioTechniques 1995, 19, 25–31. [Google Scholar]
- Veronika, U.; Olga, B.; Hanka, V. Photoperiod regulates growth of male accessory glands through juvenile hormone signaling in the linden bug, Pyrrhocoris apterus. Insect Biochem. Mol. Biol. 2016, 70, 118–120. [Google Scholar]
- Emílio, C.M.; Simone, S.; Ana, C. Growth, mortality and reproductive traits of diploid and triploid Pacific oysters in Southern Brazil. Aquac. Res. 2020, 51, 65–74. [Google Scholar]
- Kelly, V.; Beskin, C.D.; Holcomb, J.A. Larval digestion of different manure types by the black soldier fly (Diptera: Stratiomyidae) impacts associated volatile emissions. Waste Manag. 2018, 74, 141–148. [Google Scholar]
- Milad, G.; Sayed, M.G.; Saeed, M. Highly porous nanostructured copper oxide foam fiber as a sorbent for head space solid-phase microextraction of BTEX from aqueous solutions. Microchem. J. 2019, 145, 462–467. [Google Scholar]
- Yogesh, M.K.; Mohammad, R.I.; Tracy, J.B. Process development and simulation of glycerol-free biofuel from canola oil and dimethyl carbonate. Fuel Process. Technol. 2013, 114, 143–160. [Google Scholar]
- Gorrens, E.; Lecocq, A.; De Smet, J. The Use of Probiotics during Rearing of Hermetia illucens: Potential, Caveats, and Knowledge Gaps. Microorganisms 2023, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, X.; Zhang, J.; Cheng, P.; Xu, Z.; Sun, W.; Zhong, Y.; Wang, Y.; Yu, G.; Liu, H. Microplastics existence intensified bloom of antibiotic resistance in livestock feces transformed by black soldier fly. Environ. Pollut. 2023, 317, 120845. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Park, J.O.; Tanner, L.; Nagano, Y.; Rabinowitz, J.D.; Shvartsman, S.Y. Energy budget of Drosophila embryogenesis. Curr. Biol. 2019, 29, 566–567. [Google Scholar] [CrossRef]
- Allman, M.J.; Fraser, J.E.; Ritchie, S.A.; Joubert, D.A.; Simmons, C.P.; Flores, H.A. Wolbachia’s Deleterious Impact on Aedes aegypti Egg Development: The Potential Role of Nutritional Parasitism. Insects 2020, 11, 735. [Google Scholar] [CrossRef]
- Atella, G.C.; Shahabuddin, M. Differential partitioning of maternal fatty acid and phospholipid in neonate mosquito larvae. J. Exp. Biol. 2002, 205, 3623–3630. [Google Scholar] [CrossRef]
- Ohnishi, A.; Kaji, M.; Hashimoto, K.; Matsumoto, S. Screening for the genes involved in bombykol biosynthesis: Identification and functional characterization of Bombyx mori acyl carrier protein (BmACP). Front. Endocrinol. 2011, 2, 92–93. [Google Scholar] [CrossRef] [Green Version]
- Renobales, D.; Woodin, M.; Blomquist, T.S. Drosophila melanogaster fatty acid synthetase. Insect Biochem. 1986, 16, 887–894. [Google Scholar] [CrossRef]
- Yang, L.-F.; Siriamornpun, S.; Li, D. Polyunsaturated fatty acid content of edible insects in thailand. J. Food Lipids 2006, 13, 277–285. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Gómez, C.; Avendaño, C.; Harmsen, I.; Ortiz, D.; Ceballos, R.; Villamizar-Sarmiento, M.G.; Oyarzun-Ampuero, F.; Wacyk, J.; Valenzuela, C. House fly (Musca domestica) larvae meal as an ingredient with high nutritional value: Microencapsulation and improvement of organoleptic characteristics. Food Res. Int. 2021, 145, 110423. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Bueno, R.P.; González-Fernández, M.J.; Sánchez-Muros-Lozano, M.J.; García-Barroso, F.; Guil-Guerrero, J.L. Fatty acid profiles and cholesterol content of seven insect species assessed by several extraction systems. Eur. Food Res. Technol. 2016, 242, 1471–1477. [Google Scholar] [CrossRef]
- Bbosa, T.; Tamale Ndagire, C.; Muzira Mukisa, I.; Fiaboe, K.K.; Nakimbugwe, D. Nutritional Characteristics of Selected Insects in Uganda for Use as Alternative Protein Sources in Food and Feed. J. Insect Sci. 2019, 19, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Poekes, L.; Legry, V.; Schakman, O. Defective adaptive thermogenesis contributes to metabolic syndrome and liver steatosis in obese mice. Clin. Sci. 2017, 131, 146–152. [Google Scholar] [CrossRef]
- Alabaster, A.; Isoe, J.; Zhou, G. Deficiencies in acetyl-CoA carboxylase and fatty acid synthase 1 differentially affect eggshell formation and blood meal digestion in Aedes aegypti. Insect Biochem. Mol. Biol. 2011, 41, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Gondim, K.C.; Atella, G.C.; Pontes, E.G.; Majerowicz, D. Lipid metabolism in insect disease vectors. Insect Biochem. Mol. Biol. 2018, 101, 108–123. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; George, J.; Reddy, G.V.; Zeng, X.; Guerrero, A. Latest Developments in Insect Sex Pheromone Research and Its Application in Agricultural Pest Management. Insects 2021, 12, 484. [Google Scholar] [CrossRef]
- Pei, X.J.; Fan, Y.L.; Bai, Y.; Bai, T.T.; Schal, C.; Zhang, Z.F.; Chen, N.; Li, S.; Liu, T.X. Modulation of fatty acid elongation in cockroaches sustains sexually dimorphic hydrocarbons and female attractiveness. PLoS Biol. 2021, 19, e3001330. [Google Scholar] [CrossRef]
- Loh, T.C.; Fong, L.; Foo, Y.H. Utilisation of Earthworm Meal in Partial Replacement of Soybean and Fish Meals in Diets of Broilers. Appl. Anim. Res. 2009, 36, 363–376. [Google Scholar] [CrossRef]
- Hiroto, M.; Motonori, K.; Masafumi, I. Hormonal regulation of the death commitment in programmed cell death of the silkworm anterior silk glands. J. Insect Physiol. 2012, 58, 67–75. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Primer | Reverse Primer | Product Length (bp) | Purpose |
|---|---|---|---|---|
| HiACP | AAAAATGTCGCTGTCACAGGGT | CAACAAAACGATGAACCCGC | 501 | cDNA |
| dsACP | T7-CGCTGTTTATTGAGCCGTGC | T7-TTCTCGGCATCTGAATCGGG | 395 | RNAi |
| dsGFP | T7-AGATCCGCCACAACATCGAG | T7-GTCCATGCCGAGAGTGATCC | 204 | |
| qHiACP | GATGCTCCATCGACAATGCG | GGACGCGTTCACGAATTTCT | 137 | qRT-PCR |
| β-actin | AGGAGACGAAGCACAAAGCA | AGTCCAAAGCGACGTAGCAG | 150 |
| Fatty Acid | dsGFP | dsACP |
|---|---|---|
| C10:0 | 0.21 ± 0.00 | 0.17 ± 0.01 * |
| C12:0 | 9.94 ± 0.08 | 7.93 ± 0.06 ** |
| C14:0 | 2.31 ± 0.02 | 1.91 ± 0.01 ** |
| C15:0 | 0.11 ± 0.01 | 0.06 ± 0.02 |
| C16:0 | 5.24 ± 0.23 | 4.94 ± 0.23 |
| C16:1 | 1.03 ± 0.04 | 0.82 ± 0.01 ** |
| C18:0 | 0.94 ± 0.06 | 0.88 ± 0.03 |
| C18:1 | 5.09 ± 0.04 | 4.6 ± 0.14 * |
| C18:2 | 2.84 ± 0.18 | 2.76 ± 0.28 |
| C18:3 | 0.31 ± 0.03 | 0.27 ± 0.05 |
| SFA | 18.75 ± 0.15 | 15.89 ± 0.25 ** |
| MUFA | 6.12 ± 0.07 | 5.42 ± 0.12 ** |
| PUFA | 3.15 ± 0.21 | 3.03 ± 0.32 |
| Total FA | 28.02 ± 0.17 | 24.34 ± 0.24 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Liu, J.; Li, B.; Wang, S.; Chen, B.; Zhang, D. An Acyl Carrier Protein Gene Affects Fatty Acid Synthesis and Growth of Hermetia illucens. Insects 2023, 14, 300. https://doi.org/10.3390/insects14030300
Peng X, Liu J, Li B, Wang S, Chen B, Zhang D. An Acyl Carrier Protein Gene Affects Fatty Acid Synthesis and Growth of Hermetia illucens. Insects. 2023; 14(3):300. https://doi.org/10.3390/insects14030300
Chicago/Turabian StylePeng, Xiaoyan, Jiawen Liu, Baoling Li, Shengyin Wang, Bosheng Chen, and Dayu Zhang. 2023. "An Acyl Carrier Protein Gene Affects Fatty Acid Synthesis and Growth of Hermetia illucens" Insects 14, no. 3: 300. https://doi.org/10.3390/insects14030300
APA StylePeng, X., Liu, J., Li, B., Wang, S., Chen, B., & Zhang, D. (2023). An Acyl Carrier Protein Gene Affects Fatty Acid Synthesis and Growth of Hermetia illucens. Insects, 14(3), 300. https://doi.org/10.3390/insects14030300