


Metabonomic Analysis of Silkworm Midgut Reveals Differences between the Physiological Effects of an Artificial and Mulberry Leaf Diet


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Artificial Feed and Mulberry Leaves
2.2. Silkworms Rearing
2.3. Silkworm Feeding Trial
2.4. Sample Preparation and Collection
2.5. Metabolite Extraction
2.6. LC-MS/MS Analysis
2.7. Processing and Statistical Analysis of Metabolomics Data
3. Results
3.1. Phenotypic Observations and Metabolite Analysis
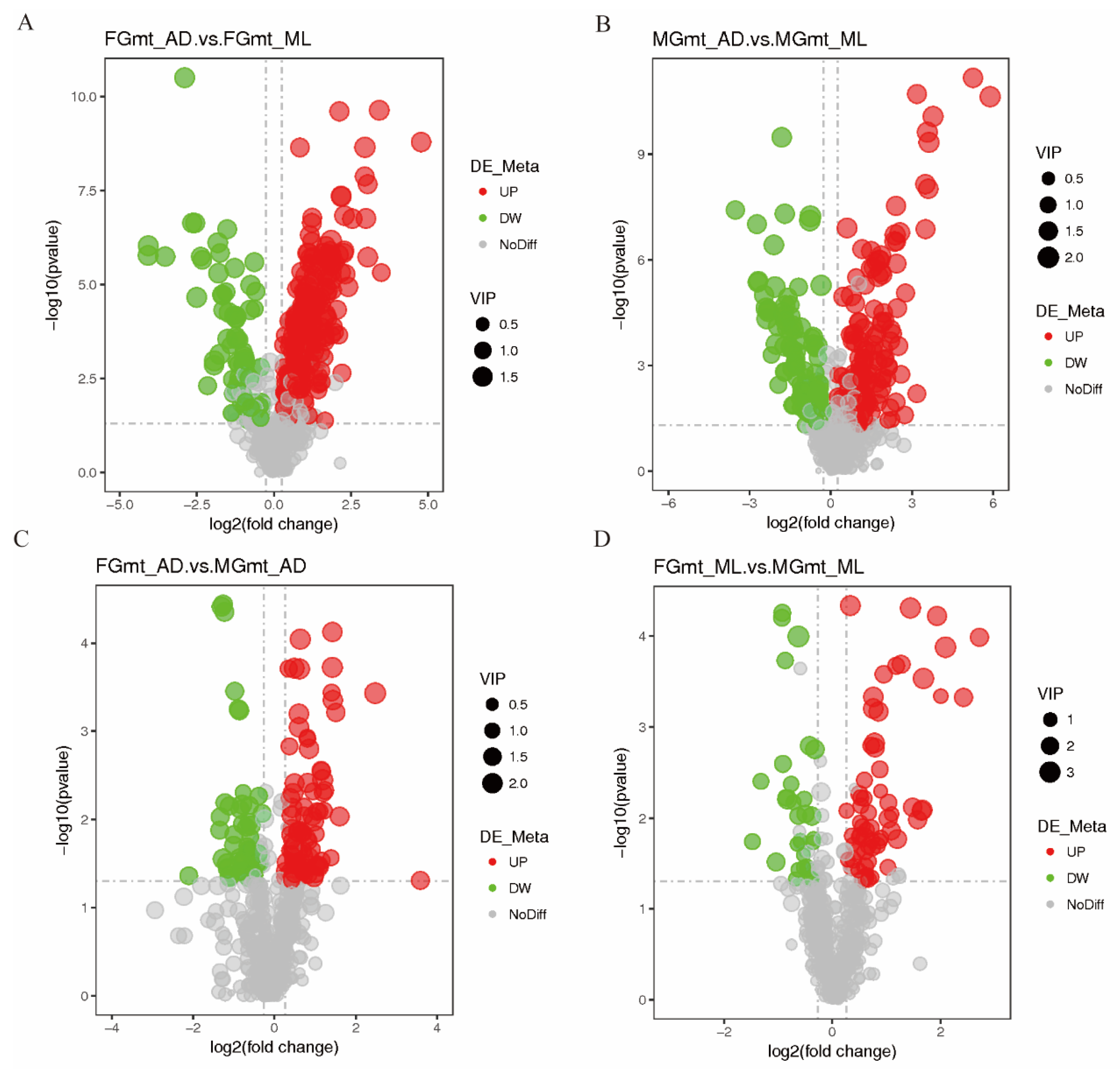
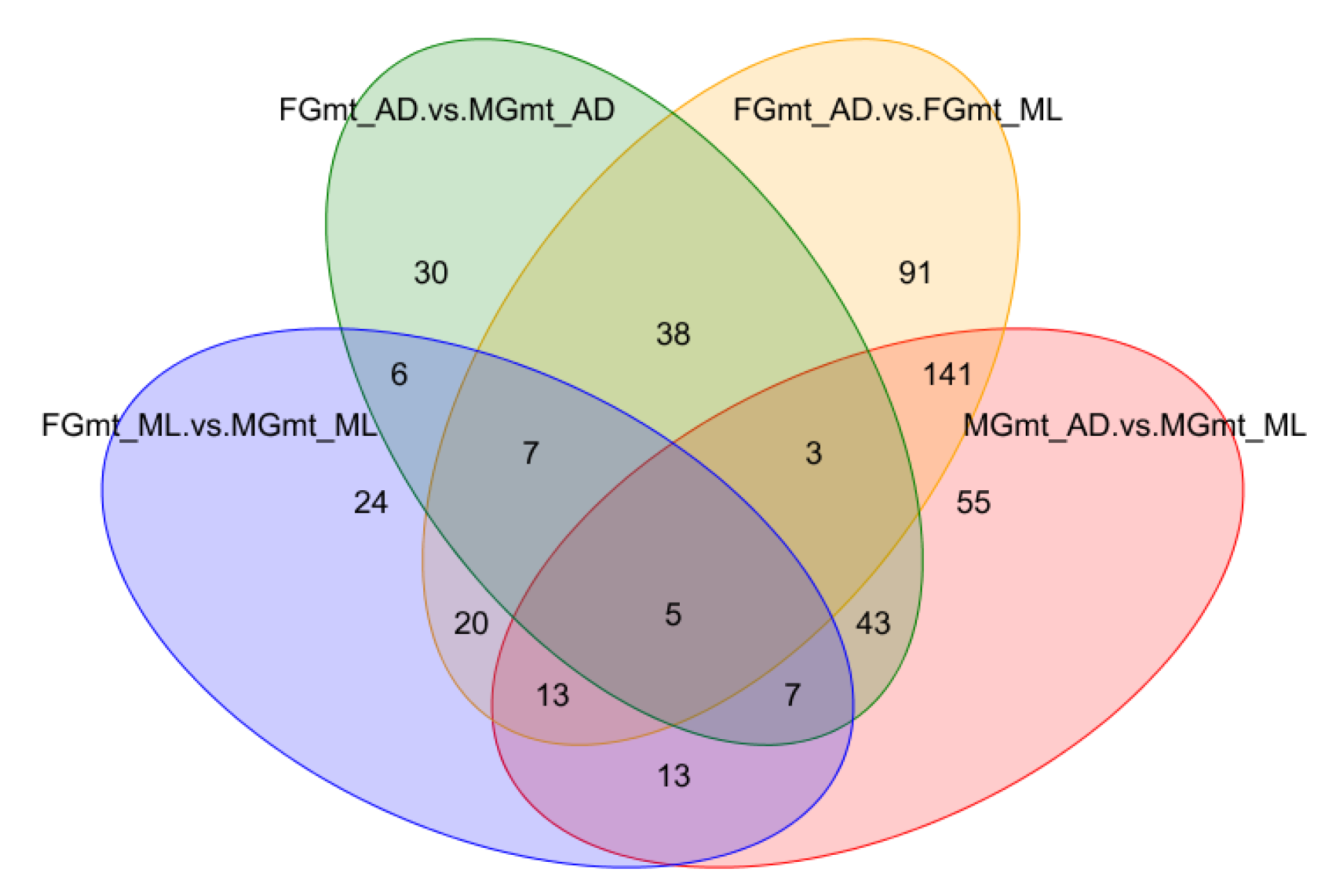
3.2. Differential Metabolite Screening
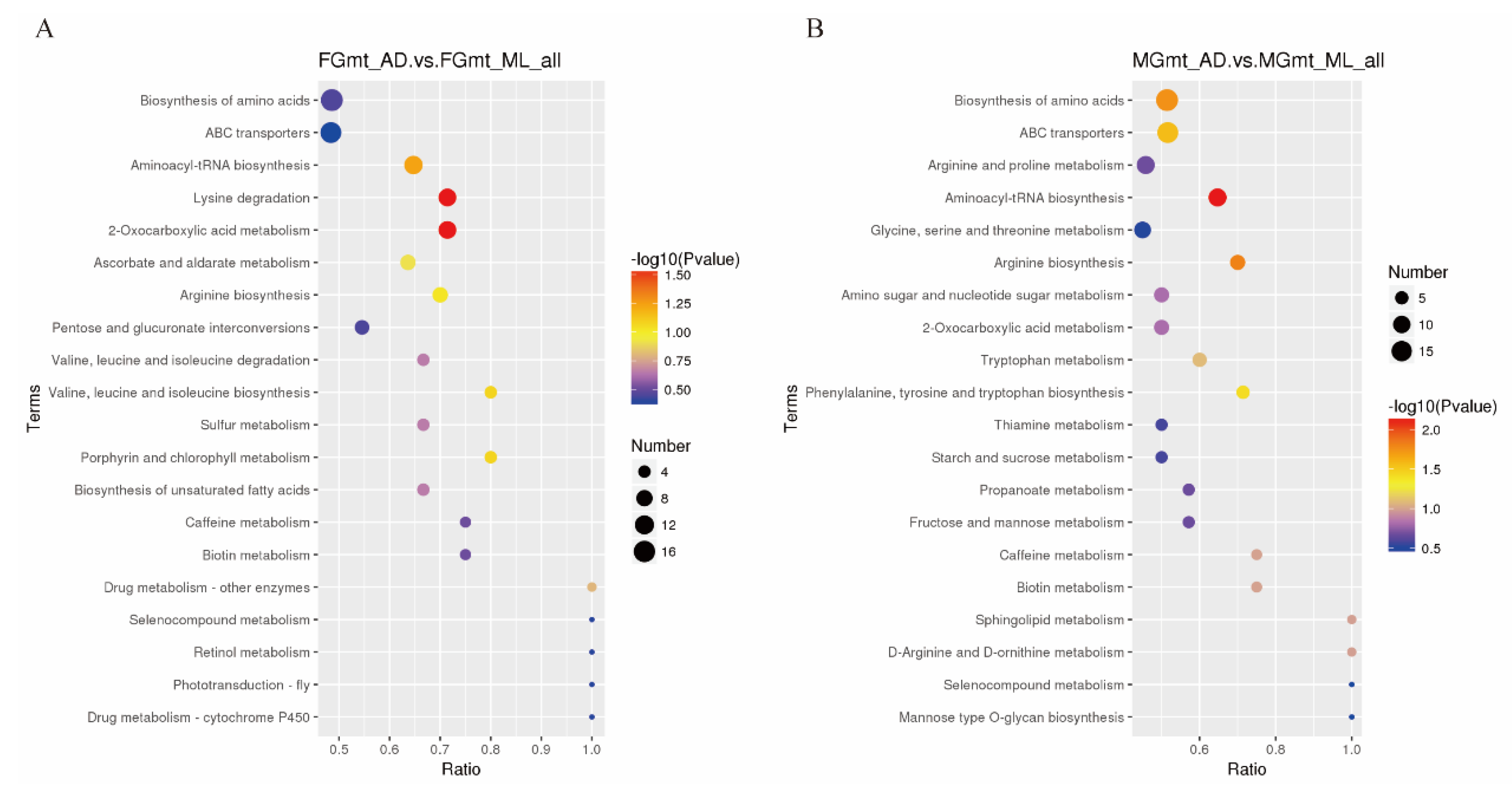
3.3. Differential Metabolite Correlation and Functional Classification
3.4. Proteome and Metabolome Association Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, Q.Y.; Guo, Y.R.; Zhang, Z.; Li, D.; Xuan, Z.L.; Li, Z.; Dai, F.Y.; Li, Y.R.; Cheng, D.J.; Li, R.Q.; et al. Complete Resequencing of 40 Genomes Reveals Domestication Events and Genes in Silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, Y.; Matsuura, K.; Nishida, J.; Hayashiya, K.; Naito, K.I. FOOD SELECTION BY SILKWORM LARVAE. Nature 1962, 194, 754. [Google Scholar] [CrossRef]
- Ito, T.; Horie, Y.; Watanabe, K.; Takamiya, K.; Furuyama, M.; Miyabaya, M.; Yamamoto, K.; Nagashim, M. Rearing of larvae of silkworm, bombyx-mori, entirely on semisynthetic diets. J. Agric. Chem. Soc. Jpn. 1974, 48, 403–407. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Mizuta, Y.; Takamiya, K.; Ueda, S.; Kimura, R.; Higuchi, T.; Takahashi, S. Growth, development, and cocoon production in artificial-diet-rearing of silkworm, bombyx-mori, with or without application of a synthetic juvenile-hormone analog. J. Agric. Chem. Soc. Jpn. 1975, 49, 41–50. [Google Scholar]
- Tsuchida, K.; Yoshitake, N. Relationship between photoperiod and secretion of the diapause hormone during larval stages of the silkworm, bombyx-mori L, reared on an artificial diet. J. Insect Physiol. 1983, 29, 755–759. [Google Scholar] [CrossRef]
- Li, X.Y.; Chiang, Y.L. Some factors influencing the larval growth of bombyx-mori L reared on artificial diets. Acta Entomol. Sin. 1982, 25, 382–389. [Google Scholar]
- Sumida, M.; Yoshio, H.; Tanaka, Y.; Matsubara, F. Developmental-changes in urea concentrations in the hemolymph of daizo(t), an original strain of the silkworm, bombyx-mori reared on an artificial diet and on fresh mulberry leaves. Comp. Biochem. Physiol. A Physiol. 1995, 110, 33–38. [Google Scholar] [CrossRef]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.L.; Zhang, S.X.; Chen, Z.H.; Tao, H.; Li, X.; Qiu, J.F.; Cui, W.Z.; Sima, Y.H.; Cui, W.Z.; Xu, S.Q. Differences in gut microbiota between silkworms (Bombyx mori) reared on fresh mulberry (Morus alba var. multicaulis) leaves or an artificial diet. RSC Adv. 2018, 8, 26200–26212. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Cheng, T.C.; Dang, Y.H.; Peng, Z.W.; Zhao, P.; Liu, S.P.; Jin, S.K.; Lin, P.; Sun, Q.; Xia, Q.Y. Identification of a midgut-specific promoter in the silkworm Bombyx mori. Biochem. Biophys. Res. Commun. 2013, 433, 542–546. [Google Scholar] [CrossRef]
- Cho, J.M.; Yoo, D.; Lee, J.Y.; Oh, M.S.; Ha, K.C.; Baek, H.I.; Lee, S.M.; Lee, J.H.; Yoo, H.J. Supplementation with a Natural Source of Amino Acids, Sil-Q1 (Silk Peptide), Enhances Natural Killer Cell Activity: A Redesigned Clinical Trial with a Reduced Supplementation Dose and Minimized Seasonal Effects in a Larger Population. Nutrients 2021, 13, 2930. [Google Scholar] [CrossRef]
- Chen, Q.M.; Liu, X.Y.; Zhao, P.; Sun, Y.H.; Zhao, X.J.; Xiong, Y.; Xu, G.W.; Xia, Q.Y. GC/MS-based metabolomic studies reveal key roles of glycine in regulating silk synthesis in silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2015, 57, 41–50. [Google Scholar] [CrossRef]
- Tang, X.; Liu, H.; Shi, Z.; Chen, Q.; Kang, X.; Wang, Y.; Zhao, P. Enhanced silk yield in transgenic silkworm (Bombyx mori) via ectopic expression of BmGT1-L in the posterior silk gland. Insect Mol. Biol. 2020, 29, 452–465. [Google Scholar] [CrossRef]
- Chen, X.D.; Ye, A.H.; Wu, X.H.; Qu, Z.G.; Xu, S.Q.; Sima, Y.; Wang, Y.J.; He, R.M.; Jin, F.; Zhan, P.F.; et al. Combined analysis of silk synthesis and hemolymph amino acid metabolism reveal key roles for glycine in increasing silkworm silk yields. Int. J. Biol. Macromol. 2022, 209, 1760–1770. [Google Scholar] [CrossRef]
- Ito, T. Application of artificial diets in sericulture. Jarq-Jpn. Agric. Res. Q. 1980, 14, 163–168. [Google Scholar]
- Lamberti, C.; Gai, F.; Cirrincione, S.; Giribaldi, M.; Purrotti, M.; Manfredi, M.; Marengo, E.; Sicuro, B.; Saviane, A.; Cappellozze, S.; et al. Investigation of the protein profile of silkworm (Bombyx mori) pupae reared on a well-calibrated artificial diet compared to mulberry leaf diet. PeerJ 2019, 7, e6723. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.Y.; Wang, G.H.; Dong, Z.M.; Xia, Q.Y.; Zhao, P. Comparative Fecal Metabolomes of Silkworms Being Fed Mulberry Leaf and Artificial Diet. Insects 2020, 11, 851. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Qian, H.Y.; Li, G.; Zhao, G.D.; Liu, M.Z.; Xu, A.Y. Metabolic Characterisation of the Midgut ofBombyx moriVarieties after BmNPV Infection Using GC-MS-Based Metabolite Profiling. Int. J. Mol. Sci. 2020, 21, 4707. [Google Scholar] [CrossRef]
- Dong, H.L.; Zhang, S.X.; Tao, H.; Chen, Z.H.; Li, X.; Qiu, J.F.; Cui, W.Z.; Sima, Y.H.; Cui, W.Z.; Xu, S.Q. Metabolomics differences between silkworms (Bombyx mori) reared on fresh mulberry (Morus) leaves or artificial diets. Sci. Rep. 2017, 7, 10972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, I.B.; Cole, S.P.C.; Kuchler, K.; Higgins, C.F. ABC Proteins: From Bacteria to Man; Elsevier Science & Technology: Jordan Hill, UK, 2002. [Google Scholar]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Dassa, E.; Bouige, P. The ABC of ABCs: A phylogenetic and functional classification of ABC systems in living organisms. Res. Microbiol. 2001, 152, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Labbe, R.; Caveney, S.; Donly, C. Genetic analysis of the xenobiotic resistance-associated ABC gene subfamilies of the Lepidoptera. Insect Mol. Biol. 2011, 20, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Miyamoto, K.; Yamamoto, K.; Narukawa, J.; Kawai, S.; Sezutsu, H.; Kobayashi, I.; Uchino, K.; Tamura, T.; Mita, K.; et al. Single amino acid mutation in an ATP-binding cassette transporter gene causes resistance to Bt toxin Cry1Ab in the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2012, 109, E1591–E1598. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.D.; Cheng, T.C.; Wang, G.H.; Duan, J.; Niu, W.H.; Xia, Q.Y. Genome-wide analysis of the ATP-binding cassette (ABC) transporter gene family in the silkworm, Bombyx mori. Mol. Biol. Rep. 2012, 39, 7281–7291. [Google Scholar] [CrossRef]
- Wang, X.; Yi, X.L.; Hou, C.X.; Wang, X.Y.; Sun, X.; Zhang, Z.J.; Qin, S.; Li, M.W. Map-based cloning and functional analysis revealed ABCC2 is responsible for Cry1Ac toxin resistance in Bombyx mori. Arch. Insect Biochem. Physiol. 2022, 110, e21886. [Google Scholar] [CrossRef]
- Liu, S.M.; Zhou, S.; Tian, L.; Guo, E.N.; Luan, Y.X.; Zhang, J.Z.; Li, S. Genome-wide identification and characterization of ATP-binding cassette transporters in the silkworm, Bombyx mori. BMC Genom. 2011, 12, 491. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.J.; Qi, J.P.; Shen, G.W.; Qin, D.Y.; Wu, J.X.; Song, Y.W.; Cao, Y.; Zhao, P.; Xia, Q.Y. Effects of Microbial Transfer during Food-Gut-Feces Circulation on the Health of Bombyx mori. Microbiol. Spectr. 2022, 10, e02357-22. [Google Scholar] [CrossRef]
- Tao, S.; Wang, J.; Liu, M.; Sun, F.; Li, B.; Ye, C. Haemolymph metabolomic differences in silkworms (Bombyx mori L.) under mulberry leaf and two artificial diet rearing methods. Arch. Insect Biochem. Physiol. 2022, 109, e21851. [Google Scholar] [CrossRef]
- Qin, L.G.; Shi, H.F.; Xia, H.C.; Chen, L.; Yao, Q.; Chen, K.P. Comparative Proteomic Analysis of Midgut Proteins From Male and Female Bombyx mori (Lepidoptera: Bombycidae). J. Insect Sci. 2014, 14, 226. [Google Scholar] [CrossRef] [Green Version]
- Kamtongdee, C.; Sumriddetchkajorn, S.; Sa-Ngiamsak, C. Feasibility Study of silkworm pupa sex identification with pattern matching. Comput. Electron. Agric. 2013, 95, 31–37. [Google Scholar] [CrossRef]
- Zhu, Z.Q.; Yuan, H.F.; Song, C.F.; Li, X.Y.; Fang, D.W.; Guo, Z.Y.; Zhu, X.Y.; Liu, W.; Yan, G.D. High-speed sex identification and sorting of living silkworm pupae using near-infrared spectroscopy combined with chemometrics. Sens. Actuators B Chem. 2018, 268, 299–309. [Google Scholar] [CrossRef]
- Zhu, M.; Hu, X.L.; Liang, Z.; Jiang, M.S.; Xue, R.Y.; Gong, Y.C.; Zhang, X.; Cao, G.L.; Gong, C.L. Functional characterization of BmOVOs in silkworm, Bombyx mori. BMC Genom. 2019, 20, 342. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.Z.; Confalonieri, F.; Jacquet, M.; Perasso, R.; Li, Z.G.; Janin, J. Silk fibroin: Structural implications of a remarkable amino acid sequence. Proteins Struct. Funct. Genet. 2001, 44, 119–122. [Google Scholar] [CrossRef]
- Xing, R.; Chen, X.D.; Zhou, Y.F.; Zhang, J.; Su, Y.Y.; Qiu, J.F.; Sima, Y.H.; Zhang, K.Q.; He, Y.; Xu, S.Q. Targeting and retention enhancement of quantum dots decorated with amino acids in an invertebrate model organism. Sci. Rep. 2016, 6, 19802. [Google Scholar] [CrossRef] [Green Version]
- Grzeskowiak, J.; Lochynska, M. Possibilities and Conditions of Developing Mulberry Silkworm Rearing in Poland, Especially for Purposes Related to the Production of Natural Cosmetics Based on Silk Proteins Obtained from Silkworm Cocoons. J. Nat. Fibers 2022, 19, 14318–14327. [Google Scholar] [CrossRef]
- Xia, Q.Y.; Li, S.; Feng, Q.L. Advances in Silkworm Studies Accelerated by the Genome Sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef]
- Hu, J.S.; Tian, J.H.; Li, F.C.; Xue, B.; Hu, J.H.; Cheng, X.Y.; Li, J.X.; Shen, W.D.; Li, B. Clone and functional analysis of Seryl-tRNA synthetase and Tyrosyl-tRNA synthetase from silkworm, Bombyx mori. Sci. Rep. 2017, 7, 41563. [Google Scholar] [CrossRef] [Green Version]
- Engsontia, P.; Sangket, U.; Chotigeat, W.; Satasook, C. Molecular Evolution of the Odorant and Gustatory Receptor Genes in Lepidopteran Insects: Implications for Their Adaptation and Speciation. J. Mol. Evol. 2014, 79, 21–39. [Google Scholar] [CrossRef]
- Guo, H.; Cheng, T.; Chen, Z.; Jiang, L.; Guo, Y.; Liu, J.; Li, S.; Taniai, K.; Asaoka, K.; Kadono-Okuda, K. Expression map of a complete set of gustatory receptor genes in chemosensory organs of Bombyx mori. Insect Biochem. Mol. Biol. 2017, 82, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R. New polar constituents of the pupae of the silkworm Bombyx mori L. I. Isolation and identification of methionine sulfoxide, methionine sulfone, and gamma-cyclic di-L-glutamate. Biosci. Biotechnol. Biochem. 2007, 71, 3055–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Hanashima, T.; Yamada, H. New Polar Constituents in the Pupae of the Silkworm Bombyx mori L. (II): Developmental Changes of the Constituents. Biosci. Biotechnol. Biochem. 2010, 74, 2133–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Kimura, M.; Kuwano, E.; Suzuki, K. Isolation of l(+)-methionine sulfoxide related to diapause induction in the silkworm, bombyx-mori. Insect Biochem. Mol. Biol. 1995, 25, 975–980. [Google Scholar] [CrossRef]
- Haghighi, A.Z.; Maples, K.R. On the mechanism of the inhibition of glutamine synthetase and creatine phosphokinase by methionine sulfoxide. J. Neurosci. Res. 1996, 43, 107–111. [Google Scholar] [CrossRef]
- Simonovic, A.D.; Anderson, M.D. Analysis of methionine oxides and nitrogen-transporting amino acids in chilled and acclimated maize seedlings. Amino Acids 2007, 33, 607–613. [Google Scholar] [CrossRef]
- Miyashita, A.; Takahashi, S.; Ishii, K.; Sekimizu, K.; Kaito, C. Primed Immune Responses Triggered by Ingested Bacteria Lead to Systemic Infection Tolerance in Silkworms. PLoS ONE 2015, 10, e0130486. [Google Scholar] [CrossRef] [Green Version]
- Nishida, S.; Ono, Y.; Sekimizu, K. Lactic acid bacteria activating innate immunity improve survival in bacterial infection model of silkworm. Drug Discov. 2016, 10, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Nishida, S.; Ishii, M.; Nishiyama, Y.; Abe, S.; Ono, Y.; Sekimizu, K. Lactobacillus paraplantarum 11-1 Isolated from Rice Bran Pickles Activated Innate Immunity and Improved Survival in a Silkworm Bacterial Infection Model. Front. Microbiol. 2017, 8, 436. [Google Scholar] [CrossRef] [Green Version]
- Rahul, K.; Anil, P.; Pooja, M.; Mihir, R.; Vankadara, S. Efficacy of phototrophic bacterial feed supplementation on economic traits and disease resistance in mulberry silkworm, Bombyx mori (Lepidoptera: Bombycidae). Rev. Soc. Entomol. Argent. 2022, 81, 28–40. [Google Scholar] [CrossRef]
- Weissbach, H.; Etienne, F.; Hoshi, T.; Heinemann, S.H.; Lowther, W.T.; Matthews, B.; St John, G.; Nathan, C.; Brot, N. Peptide methionine sulfoxide reductase: Structure, mechanism of action, and biological function. Arch. Biochem. Biophys. 2002, 397, 172–178. [Google Scholar] [CrossRef]
- Santana, T.P.; Gasparino, E.; de Sousa, F.C.B.; Khatlab, A.S.; Zancanela, V.; Brito, C.O.; Barbosa, L.T.; Fernandes, R.P.M.; Del Vesco, A.P. Effects of free and dipeptide forms of methionine supplementation on oxidative metabolism of broilers under high temperature. Animal 2021, 15, 100173. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compared Samples | Number of Total Identify | Number of Total Significant | Number of Significant Up | Number of Significant Down |
|---|---|---|---|---|
| MGmt_AD. vs. MGmt_ML | 758 | 280 | 183 | 97 |
| FGmt_AD. vs. FGmt_ML | 758 | 318 | 245 | 73 |
| FGmt_AD. vs. MGmt_AD | 758 | 139 | 90 | 49 |
| FGmt_ML. vs. MGmt_ML | 758 | 95 | 65 | 30 |
| Metabolites | Class | FC (Female) | FC (Male) |
|---|---|---|---|
| Methionine sulfoxide | Amino acids and their derivatives | 11.18 | 59.43 |
| Nε-(1-carboxymethyl)-L-lysine | Amino acids and their derivatives | 27.35 | 38.38 |
| Lysophosphatidylcholine 16:1 | Phospholipid | 8.22 | 12.20 |
| 2,6-Diaminooimelic acid | Amino acids and their derivatives | 10.66 | 11.34 |
| Imatinib | Other | 7.70 | 11.32 |
| Sachydrine | Amino acids and their derivatives | 8.24 | 9.11 |
| Lysophosphatidylcholine 15:1 | Phospholipid | 7.90 | 6.70 |
| 18-Hydroxycorticosterone | Steroids and their derivatives | 0.06 | 0.09 |
| 20-Carboxy-Leukotriene B4 | Fatty acyl | 0.13 | 0.15 |
| Carnitine-C5 | Carnitines | 0.17 | 0.15 |
| 2-Methylbutyroylcarnitine | Fatty acyl | 0.16 | 0.16 |
| Map Title | Map ID | Differential Proteins p Value | Differential Metabolites p Value |
|---|---|---|---|
| Pyruvate metabolism | map00620 | 1.00 | 0.75 |
| Drug metabolism–cytochrome P450 | map00982 | 0.22 | 0.40 |
| Pyrimidine metabolism | map00240 | 0.15 | 0.82 |
| Tryptophan metabolism | map00380 | 0.40 | 0.53 |
| Valine, leucine, and isoleucine degradation | map00280 | 0.28 | 0.22 |
| Glycerolipid metabolism | map00561 | 0.50 | 1.00 |
| Lysine degradation | map00310 | 0.34 | 0.03 |
| Ascorbate and aldarate metabolism | map00053 | 0.40 | 0.12 |
| Histidine metabolism | map00340 | 0.34 | 0.78 |
| Glycine, serine, and threonine metabolism | map00260 | 1.00 | 0.47 |
| Cysteine and methionine metabolism | map00270 | 0.49 | 0.47 |
| Map Title | Map ID | Differential Proteins p Value | Differential Metabolites p Value |
|---|---|---|---|
| mTOR signaling pathway | map04150 | 0.24 | 0.54 |
| Glycerolipid metabolism | map00561 | 0.40 | 0.54 |
| Glutathione metabolism | map00480 | 0.16 | 1.00 |
| Caffeine metabolism | map00232 | 0.24 | 0.10 |
| Arginine biosynthesis | map00220 | 1.00 | 0.02 |
| Inositol phosphate metabolism | map00562 | 0.42 | 0.66 |
| Phenylalanine metabolism | map00360 | 0.24 | 1.00 |
| Drug metabolism–other enzymes | map00983 | 0.24 | 0.54 |
| Biosynthesis of amino acids | map01230 | 0.33 | 0.02 |
| Thiamine metabolism | map00730 | 0.42 | 0.28 |
| Sphingolipid metabolism | map00600 | 0.06 | 0.10 |
| Metabolic pathways | map01100 | 0.63 | 0.83 |
| Galactose metabolism | map00052 | 0.15 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Deng, J.; Deng, X.; Liu, L.; Zha, X. Metabonomic Analysis of Silkworm Midgut Reveals Differences between the Physiological Effects of an Artificial and Mulberry Leaf Diet. Insects 2023, 14, 347. https://doi.org/10.3390/insects14040347
Li J, Deng J, Deng X, Liu L, Zha X. Metabonomic Analysis of Silkworm Midgut Reveals Differences between the Physiological Effects of an Artificial and Mulberry Leaf Diet. Insects. 2023; 14(4):347. https://doi.org/10.3390/insects14040347
Chicago/Turabian StyleLi, Juan, Jing Deng, Xuan Deng, Lianlian Liu, and Xingfu Zha. 2023. "Metabonomic Analysis of Silkworm Midgut Reveals Differences between the Physiological Effects of an Artificial and Mulberry Leaf Diet" Insects 14, no. 4: 347. https://doi.org/10.3390/insects14040347
APA StyleLi, J., Deng, J., Deng, X., Liu, L., & Zha, X. (2023). Metabonomic Analysis of Silkworm Midgut Reveals Differences between the Physiological Effects of an Artificial and Mulberry Leaf Diet. Insects, 14(4), 347. https://doi.org/10.3390/insects14040347