

Winners and Losers: Cordulegaster Species under the Pressure of Climate Change
 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
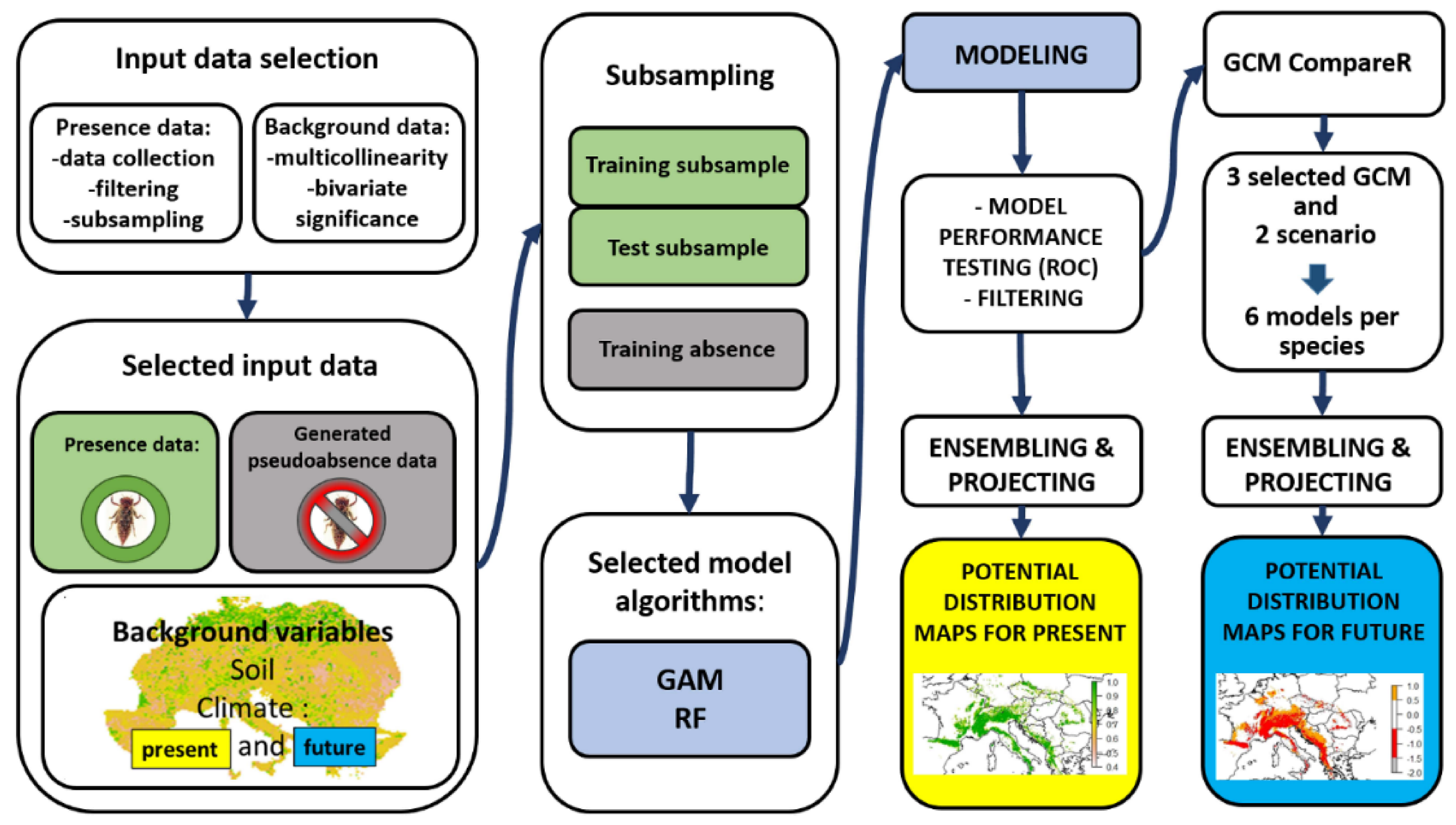
2. Materials and Methods
2.1. Occurrence Data
2.2. Environmental Variables
2.3. Future Climate Variables
2.4. Algorithm Selection
2.5. Modeling
3. Results
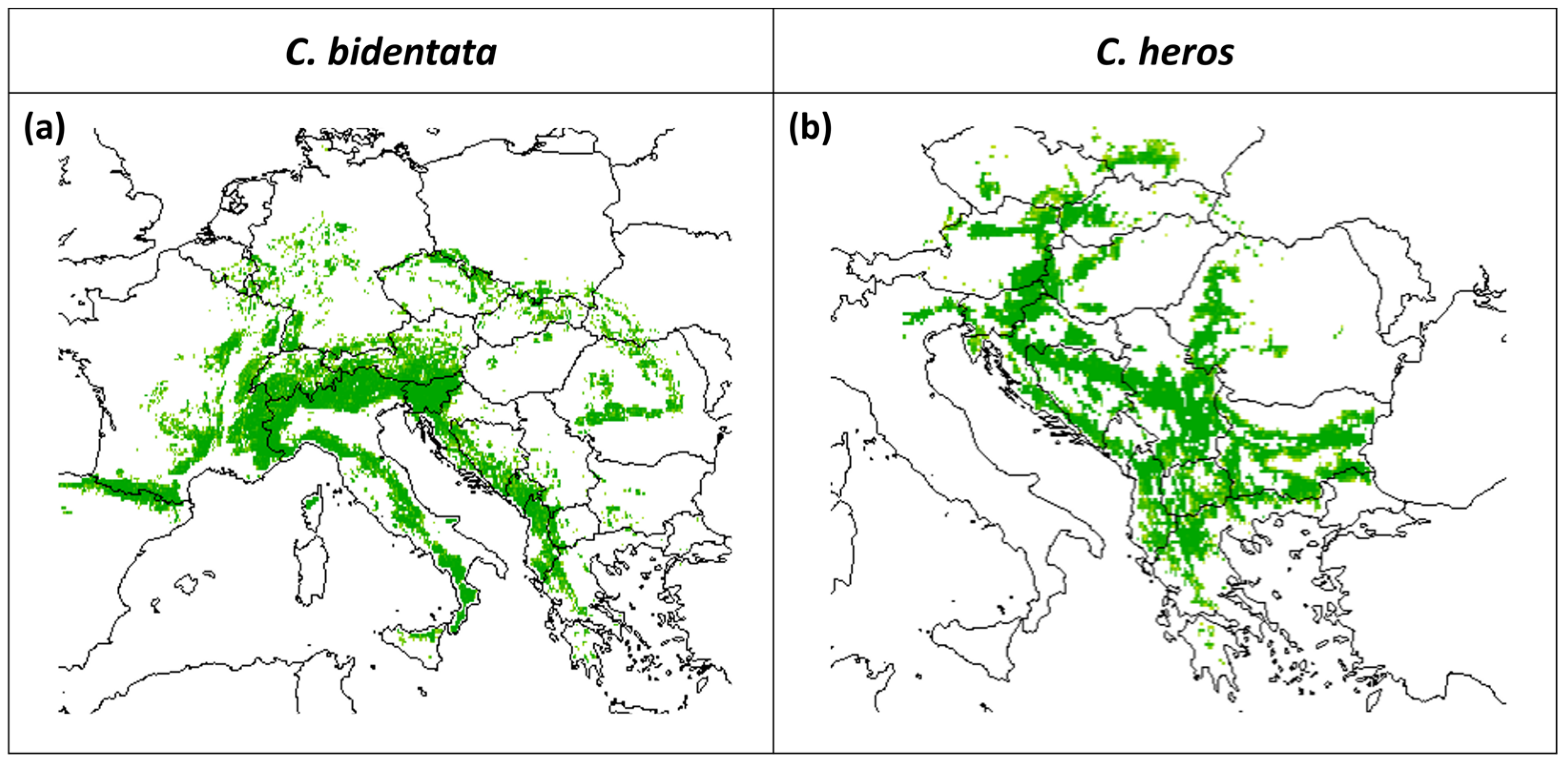
3.1. Present Distribution
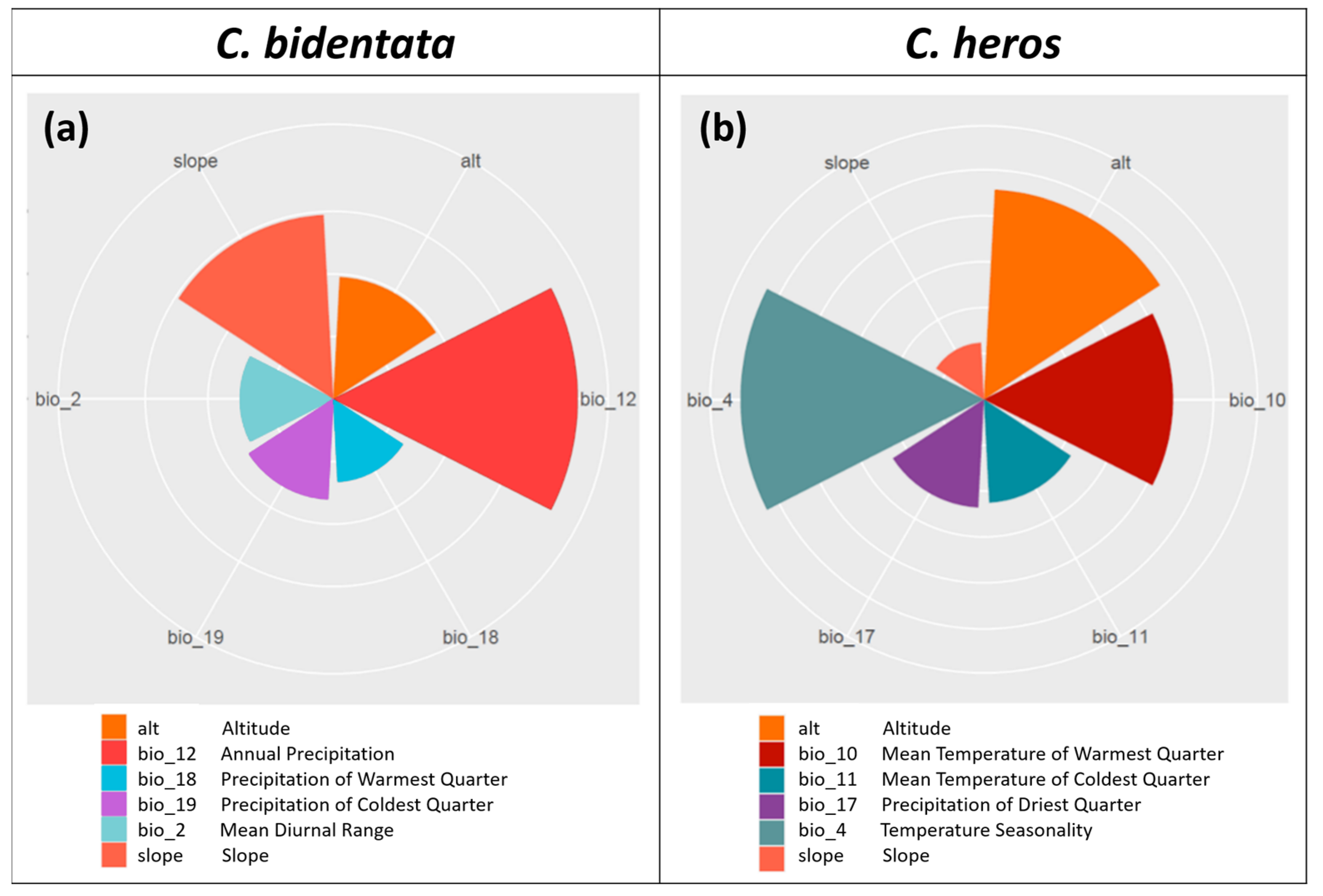
3.2. Ecological Niche Characteristics of the Species
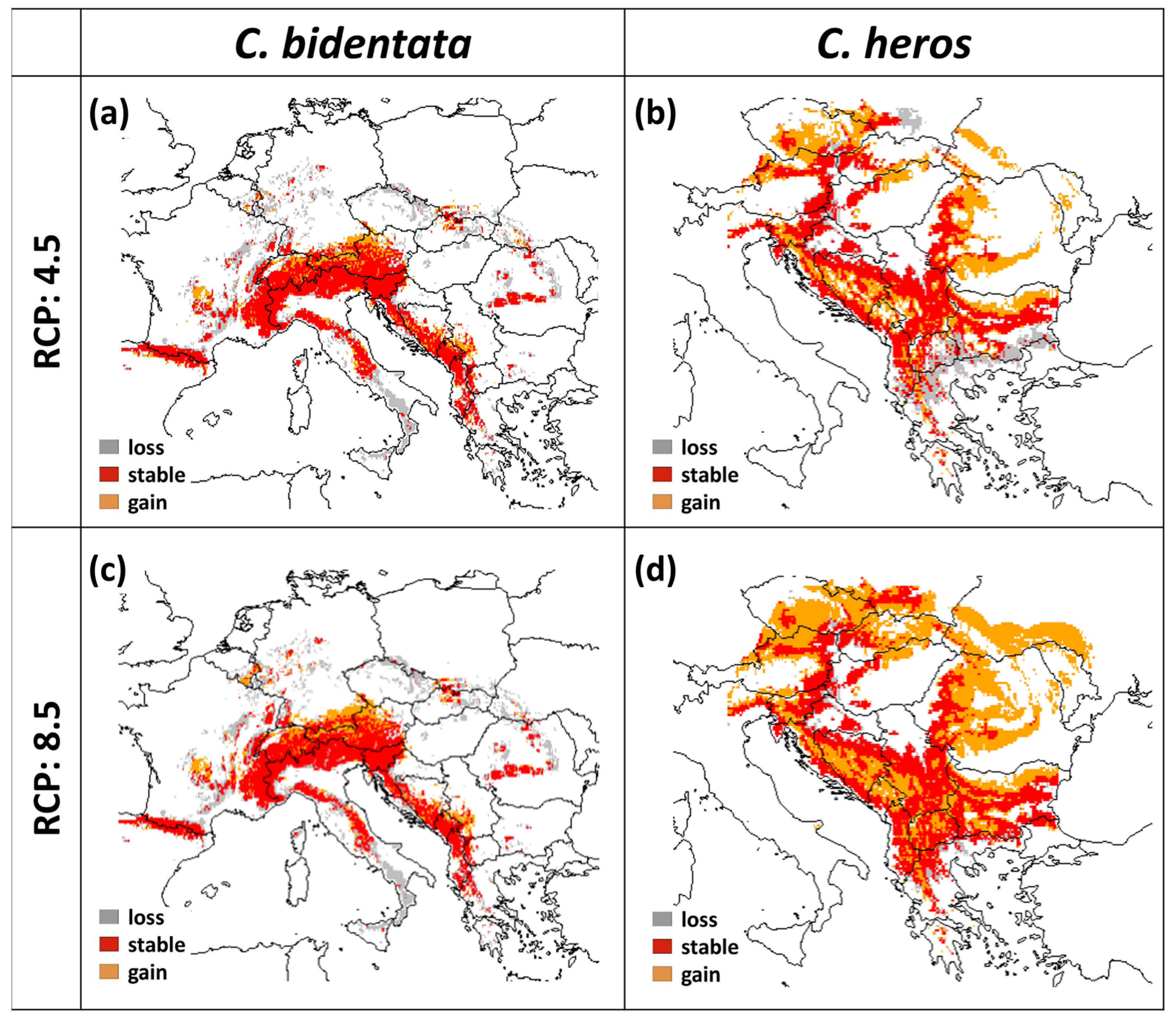
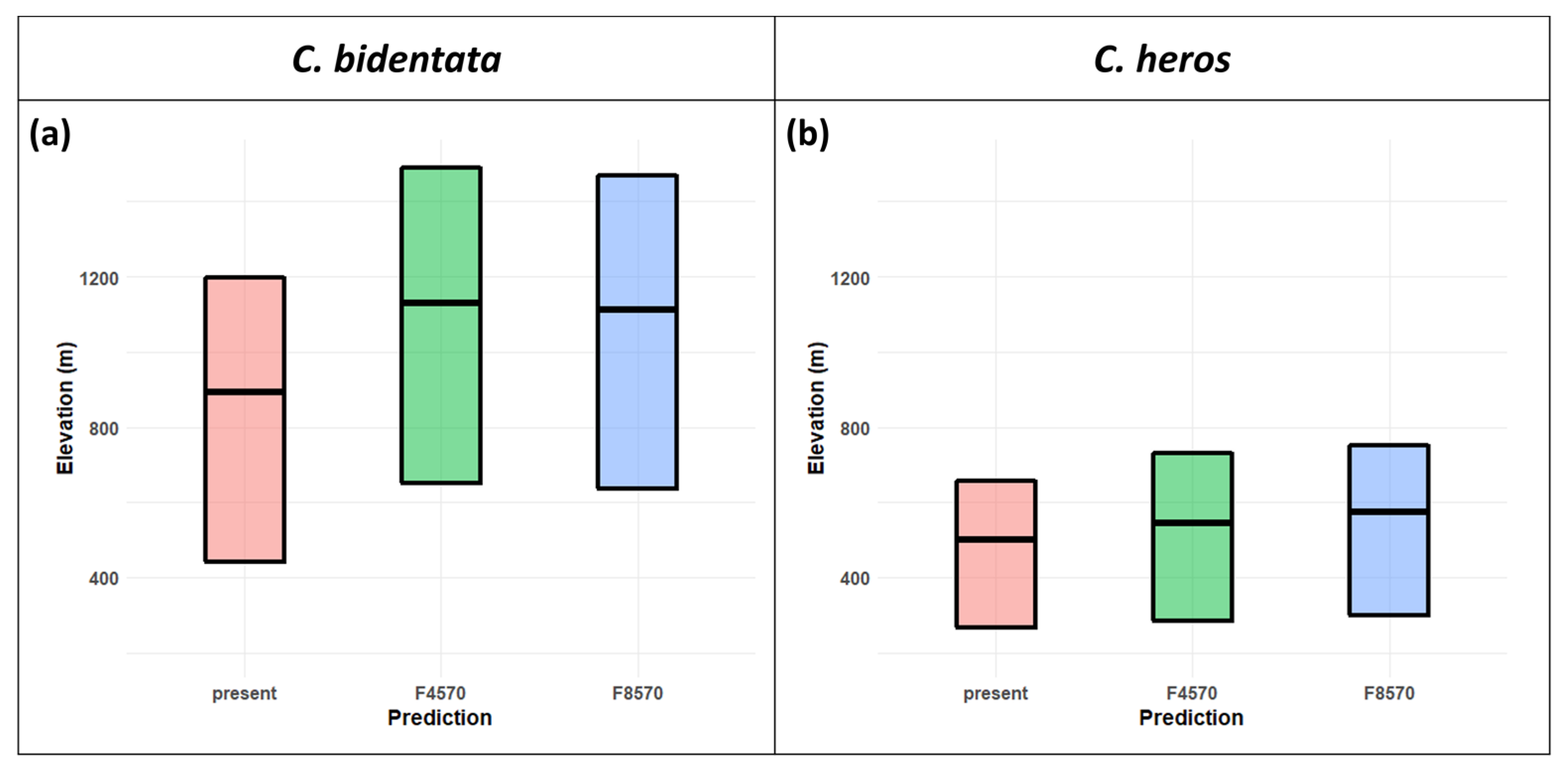
3.3. Future Projections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Malmqvist, B.; Rundle, S. Threats to the running water ecosystems of the world. Environ. Conserv. 2002, 29, 134–153. [Google Scholar] [CrossRef]
- Palmer, M.A.; Lettenmaier, D.P.; Poff, N.L.; Postel, S.L.; Richter, B.; Warner, R. Climate change and river ecosystems: Protection and adaptation options. Environ. Manag. 2009, 44, 1053–1068. [Google Scholar] [CrossRef] [PubMed]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- Thorp, J.H.; Cothran, M.L. Regulation of Freshwater Community Structure at Multiple Intensities of Dragonfly Predation. Ecology 1984, 65, 1546–1555. [Google Scholar] [CrossRef] [Green Version]
- Van Buskirk, J. Interactive effects of dragonfly predation in experimental pond communities. Ecology 1988, 69, 857–867. [Google Scholar] [CrossRef]
- Worthen, W.; Fravel, R.; Horne, C. Downstream Changes in Odonate (Insecta: Odonata) Communities along a Suburban to Urban Gradient: Untangling Natural and Anthropogenic Effects. Insects 2021, 12, 201. [Google Scholar] [CrossRef]
- Brown, K.S., Jr. Conservation of neotropical environments: Insects as indicators. In The Conservation of Insects and Their Habitats; Collins, N.M., Thomas, J.A., Eds.; Academic Press: London, UK, 1991; pp. 349–404. [Google Scholar]
- Samways, M.J.; Steytler, N.S. Dragonfly (Odonata) distribution patterns in urban and forest landscapes, and recommendations for riparian management. Biol. Conserv. 1996, 78, 279–288. [Google Scholar] [CrossRef]
- Watson, J.A.L.; Arthington, A.H.; Conrick, D.L. Effect of sewage effluent on dragonflies of Bulimba Creek, Brisbane. Aust. J. Mar. Freshw. Res. 1982, 33, 517–528. [Google Scholar] [CrossRef]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; De Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Boudot, J.P.; Kalkman, V.J. (Eds.) Atlas of the European Dragonflies and Damselflies; KNNV Publishing: Leiden, The Netherlands, 2015. [Google Scholar]
- Balázs, A.; Fric, Z.F.; Holuša, O. Flying activity and population dynamics of Cordulegaster heros Theischinger, 1979 (Insecta: Odonata: Cordulegastridae) in Slovakia. Int. J. Odonatol. 2020, 23, 155–163. [Google Scholar] [CrossRef]
- Boda, R.; Bereczki, C.; Pernecker, B.; Mauchart, P.; Csabai, Z. Life history and multiscale habitat preferences of the red-listed Balkan Goldenring, Cordulegaster heros Theischinger, 1979 (Insecta, Odonata), in South-Hungarian headwaters: Does the species have mesohabitat-mediated microdistribution? Hydrobiologia 2015, 760, 121–132. [Google Scholar] [CrossRef]
- Fekete, J.; Ézsöl, T.; Kloknicer, T.; Katona, G. Further data on Cordulegaster bidentata Selys, 1873 from the territory of Bükk National Park. Folia Hist.-Nat. Musei Matra. 2018, 42, 11–14. Available online: http://real.mtak.hu/id/eprint/142017 (accessed on 28 March 2023).
- IUCN (2021): International Union for Conservation of Nature Annual Report; IUCN: Gland, Switzerland, 2022.
- Lang, C.; Müller, H.; Waringer, J.A. Larval habitats and longitudinal distribution patterns of Cordulegaster heros Theischinger and C. bidentata Sélys in an Austrian forest stream (Anisoptera: Cordulegastridae). Odonatologica 2001, 30, 395–409. [Google Scholar]
- Froufe, E.; Ferreira, S.; Boudot, J.P.; Alves, P.C.; Harris, D.J. Molecular phylogeny of the Western Palaearctic Cordulegaster taxa (Odonata: Anisoptera: Cordulegastridae). Biol. J. Linn. Soc. 2014, 111, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Bánkuti, K.; Kovács, T. Adatok az Északborsodi-hegyvidék Odonata faunájához. Folia Hist.-Nat. Musei Matra. 1994, 19, 51–58. Available online: http://real.mtak.hu/141193/1/51-58.pdf (accessed on 28 March 2023).
- De Knijf, G.; Vanappelghem, C.; Demolder, H. Odonata from Montenegro, with notes on taxonomy, regional diversity and conservation. Odonatologica 2013, 42, 1–29. Available online: https://purews.inbo.be/ws/files/289719/DeKnijf_etal_2013_Odonatologica.pdf (accessed on 28 March 2023).
- Boudot, J.-P. IUCN Red List of Threatened Species (IUCN SSC Dragonfly Specialist Group) Cordulegaster heros: 2018. [CrossRef]
- Csordás, L.; Ferincz, Á.; Lőkkös, A.; Rozner, G.Y. New data on the distribution of Large Golden Ringed Dragonfly (Cordulegaster heros Theischinger, 1979) (Odonata) in Zselic hills. Nat. Som. 2009, 15, 53–56. [Google Scholar] [CrossRef]
- Rozner Gy Ferincz, Á.; Miókovics, E. Adatok a (Cordulegaster bidentata Sélys, 1843) és a kétcsíkos hegyiszitakötő (Cordulegaster heros Theischinger, 1979) elterjedéséhez a Bakonyban. Természetvédelmi Közlemények 2012, 18, 447–455. [Google Scholar]
- Bahlo, K. Rheobionte Libellen (Odonata) in Fliessgewässern der Lüneburger Heide (Niedersachsen). Braunschw. Nat. Schr. 1989, 3, 407–418. [Google Scholar]
- Collins, S.D.; McIntyre, N.E. Modeling the distribution of odonates: A review. Freshw. Sci. 2015, 34, 1144–1158. [Google Scholar] [CrossRef]
- Gibbs, K.E.; Bradeen, B.; Boland, D. Spatial and Temporal Segregation Among Six Species of Coexisting Ophiogomphus (Odonata: Gomphidae) in the Aroostook River, Maine. Northeast. Nat. 2004, 11, 295–312. [Google Scholar] [CrossRef]
- Corser, J.D. Status and Ecology of a Rare Gomphid Dragonfly at the Northern Extent of its Range. Northeast. Nat. 2010, 17, 341–345. [Google Scholar] [CrossRef]
- De Almeida, M.C.; Cortes, L.G.; De Marco, P., Jr. New records and a niche model for the distribution of two Neotropical damselflies: Schistolobos boliviensis and Tuberculobasis inversa (Odonata: Coenagrionidae). Insect Conserv. Divers. 2010, 3, 252–256. [Google Scholar] [CrossRef]
- Domisch, S.; Jähnig, S.C.; Haase, P. Climate-change winners and losers: Stream macroinvertebrates of a submontane region in Central Europe. Freshw. Biol. 2011, 56, 2009–2020. [Google Scholar] [CrossRef]
- Domisch, S.; Araújo, M.B.; Bonada, N.; Pauls, S.U.; Jähnig, S.C.; Haase, P. Modelling distribution in European stream macroinvertebrates under future climates. Glob. Chang. Biol. 2013, 19, 752–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch, J.M.; Samways, M.J.; Hill, T.R.; Piper, S.E.; Taylor, S. Application of predictive distribution modelling to invertebrates: Odonata in South Africa. Biodivers. Conserv. 2006, 15, 4239–4251. [Google Scholar] [CrossRef]
- Howard, T.G. Salmon River Watershed Inventory and Landscape Analysis; New York Natural Heritage Program: Albany, NY, USA, 2006. [Google Scholar]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation during the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- De Knijf, G.; Flenker, U.; Vanappelghem, C.; Manci, C.O.; Kalkman, V.J.; Demolder, H. The status of two boreo-alpine species,Somatochlora alpestrisandS. arctica, in Romania and their vulnerability to the impact of climate change (Odonata: Corduliidae). Int. J. Odonatol. 2011, 14, 111–126. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.-C. Latitudinal and Elevational Range Shifts under Contemporary Climate Change. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 599–611. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Kaschner, K.; Tittensor, D.P.; Ready, J.; Gerrodette, T.; Worm, B. Current and Future Patterns of Global Marine Mammal Biodiversity. PLoS ONE 2011, 6, e19653. [Google Scholar] [CrossRef]
- Mizsei, E.; Üveges, B.; Vági, B.; Szabolcs, M.; Lengyel, S.; Pfliegler, W.P.; Nagy, Z.T.; Tóth, J.P. Species distribution modelling leads to the discovery of new populations of one of the least known European snakes, Vipera ursinii graeca, in Albania. Amphib. -Reptil. 2016, 37, 55–68. Available online: https://brill.com/view/journals/amre/37/1/article-p55_6.xml (accessed on 28 March 2023). [CrossRef]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CABI Rev. 2019, 1–3. [Google Scholar] [CrossRef]
- Bennie, J.; Wilson, R.; Maclean, I.; Suggitt, A. Seeing the woods for the trees—When is microclimate important in species distribution models? Glob. Chang. Biol. 2014, 20, 2699–2700. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.R.; Porter, W.P. NicheMapR—An R package for biophysical modelling: The microclimate model. Ecography 2017, 40, 664–674. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological niches and geographic distributions (MPB-49). In Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; Mcdonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Lannuzel, G.; Balmot, J.; Dubos, N.; Thibault, M.; Fogliani, B. High-resolution topographic variables accurately predict the distribution of rare plant species for conservation area selection in a narrow-endemism hotspot in New Caledonia. Biodivers. Conserv. 2021, 30, 963–990. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Bystriakova, N.; Peregrym, M.; Erkens, R.H.; Bezsmertna, O.; Schneider, H. Sampling bias in geographic and environmental space and its effect on the predictive power of species distribution models. Syst. Biodivers. 2012, 10, 305–315. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- WorldClim, Maps, Graphs, Tables, and DATA of the Globla Climate. Available online: http://www.worldclim.org/ (accessed on 16 July 2020).
- Hijmans, R.J. (2020) raster: Geographic Data Analysis and Modeling. R package version 2.8–19. 2019. Available online: https://cran.r-project.org/package=raster (accessed on 22 April 2019).
- Bivand, R.; Rundel, C.; Pebesma, E.; Stuetz, R.; Hufthammer, K.O.; Bivand, M.R. Package ‘rgeos’. The Comprehensive R Archive Network (CRAN). 2017.
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography: Moving beyond static species distribution models. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martínez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Barbosa, A.M. fuzzySim: Applying fuzzy logic to binary similarity indices in ecology. Methods Ecol. Evol. 2015, 6, 853–858. [Google Scholar] [CrossRef]
- Fajardo, J.; Corcoran, D.; Roehrdanz, P.R.; Hannah, L.; Marquet, P.A. GCM compare R: A web application to assess differences and assist in the selection of general circulation models for climate change research. Methods Ecol. Evol. 2020, 11, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package ‘biomod2′. Species distribution modeling within an ensemble forecasting framework. Ecography 2016, 32, 369–373. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Ridgeway, G. Generalized Boosted Models: A Guide to the Gbm Package. 2012. Available online: http://gradientboostedmodels.googlecode.com/git/gbm/inst/doc/gbm.pdf (accessed on 12 April 2018).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Lewin-Koh, J.N.; Bivand, R. “Package ‘Maptools’: Tools for Reading and Handling Spatial Objects,” R Package Version 1.1-6. 2022. Available online: https://cran.r-project.org/web/packages/maptools/index.html (accessed on 2 December 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 15 February 2019).
- Allan, J.D.; Palmer, M.; Poff, N.L. Climate change and freshwater ecosystems. In Climate Change and Biodiversity; Yale University Press: New Haven, CT, USA, 2005; pp. 274–290. [Google Scholar]
- Xenopoulos, M.A.; Lodge, D.M.; Alcamo, J.; Marker, M.; Schulze, K.; Van Vuuren, D.P. Scenarios of freshwater fish extinctions from climate change and water withdrawal. Glob. Chang. Biol. 2005, 11, 1557–1564. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.B.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the Use of Species Distribution Models in Conservation Planning and Management under Climate Change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Danyik, T.; Kovács, T.; Olajos, P. Magyarország Szitakötőinek Kézikönyve; Magyar Természettudományi Múzeum: Herman Ottó Intézet: Hungary, Budapest, 2018; 290p. [Google Scholar]
- Hynes, H.B.N. The Ecology of Running Waters; Liverpool University Press: Liverpool, UK, 1970; Volume 555. [Google Scholar]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; De Knijf, G.; Suhling, F.; Termaat, T. Diversity and conservation of European dragonflies and damselflies (Odonata). Hydrobiologia 2018, 811, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Boudot, J.-P. IUCN Red List of Threatened Species (IUCN SSC Dragonfly Specialist Group) Cordulegaster bidentata: 2018. [CrossRef]
- Grewe, Y.; Hof, C.; Dehling, D.M.; Brandl, R.; Brändle, M. Recent range shifts of European dragonflies provide support for an inverse relationship between habitat predictability and dispersal. Glob. Ecol. Biogeogr. 2012, 22, 403–409. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A northward shift of range margins in British Odonata. Glob. Chang. Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Fox, R.; Thomas, C.D. The distributions of a wide range of taxonomic groups are expanding polewards. Glob. Chang. Biol. 2006, 12, 450–455. [Google Scholar] [CrossRef]
- Pernecker, B.; Mauchart, P.; Csabai, Z. What to do if streams go dry? Behaviour of Balkan Goldenring (Cordulegaster heros, Odonata) larvae in a simulated drought experiment in SW Hungary. Ecol. Entomol. 2020, 45, 1457–1465. [Google Scholar] [CrossRef]
- Boudot, J.P. Les Cordulegaster du Paléarctique occidental: Identification et répartition (Odonata, Anisoptera, Cordulegastridae). Martinia 2001, 17, 3–34. [Google Scholar]
- Schneider, T.; Vierstraete, A.; Müller, O.; van Pelt, G.J.; Caspers, M.; Ikemeyer, D.; Snegovaya, N.; Dumont, H.J. Taxonomic revision of eastern part of Western Palaearctic Cordulegaster using molecular phylogeny and morphology, with the description of two new species (Odonata: Anisoptera: Cordulegastridae). Diversity 2021, 13, 667. [Google Scholar] [CrossRef]
- Vilenica, M.; Kulijer, D.; Gligorovic, B.; Gligorovic, A.; De Knijf, G. Distribution, habitat requirements and vulnerability of Caliaeschna microstigma (Odonata: Aeshnidae) at the north-western edge of the species’range. Odonatologica 2021, 50, 203–225. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. bidentata | C. heros | ||
|---|---|---|---|
| Variable | Scores | Variable | Scores |
| Annual Precipitation (bio_12) | 0.390 | Temperature Seasonality (bio_4) | 0.265 |
| Slope | 0.295 | Altitude | 0.228 |
| Altitude | 0.195 | Mean Temperature of Warmest Quarter (bio_10) | 0.206 |
| Precipitation of Coldest Quarter (bio_19) | 0.161 | Precipitation of Driest Quarter; (bio_17) | 0.118 |
| Mean Diurnal Range (bio_2) | 0.149 | Mean Temperature of Coldest Quarter (bio_11) | 0.113 |
| Precipitation of Warmest Quarter (bio_18) | 0.133 | Slope | 0.062 |
| C. bidentata | C. heros | |||
|---|---|---|---|---|
| RCP45 | RCP85 | RCP45 | RCP85 | |
| Loss (Cell Number) | 2841 | 3613 | 706 | 392 |
| Stable (Cell Number) | 5264 | 5252 | 9917 | 8066 |
| Gain (Cell Number) | 816 | 1073 | 2375 | 4969 |
| Percentage of Loss | 35.1 | 40.8 | 15.6 | 10.39 |
| Percentage of Gain | 10.1 | 12.1 | 52.6 | 131.7 |
| Species Range Change | −25 | −28.7 | 37 | 121.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fekete, J.; De Knijf, G.; Dinis, M.; Padisák, J.; Boda, P.; Mizsei, E.; Várbíró, G. Winners and Losers: Cordulegaster Species under the Pressure of Climate Change. Insects 2023, 14, 348. https://doi.org/10.3390/insects14040348
Fekete J, De Knijf G, Dinis M, Padisák J, Boda P, Mizsei E, Várbíró G. Winners and Losers: Cordulegaster Species under the Pressure of Climate Change. Insects. 2023; 14(4):348. https://doi.org/10.3390/insects14040348
Chicago/Turabian StyleFekete, Judit, Geert De Knijf, Marco Dinis, Judit Padisák, Pál Boda, Edvárd Mizsei, and Gábor Várbíró. 2023. "Winners and Losers: Cordulegaster Species under the Pressure of Climate Change" Insects 14, no. 4: 348. https://doi.org/10.3390/insects14040348
APA StyleFekete, J., De Knijf, G., Dinis, M., Padisák, J., Boda, P., Mizsei, E., & Várbíró, G. (2023). Winners and Losers: Cordulegaster Species under the Pressure of Climate Change. Insects, 14(4), 348. https://doi.org/10.3390/insects14040348