

Developmental Biology and Identification of a Garden Pest, Otiorhynchus (Podoropelmus) smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae, Entiminae), with Comments on Its Origin and Distribution

 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Materials
2.2. Molecular Study
2.3. Morphology and Larval Instar Identification
3. Results
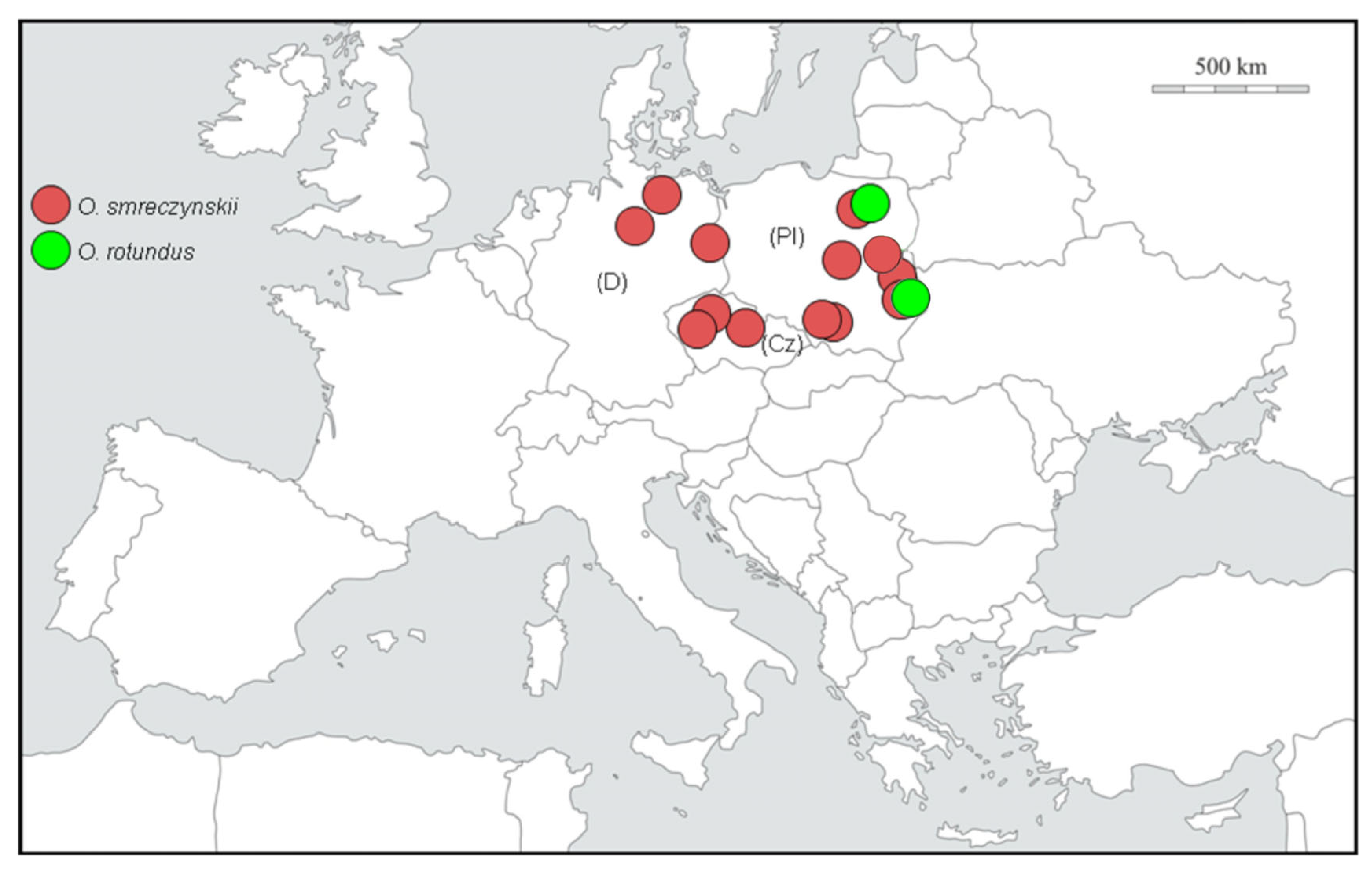
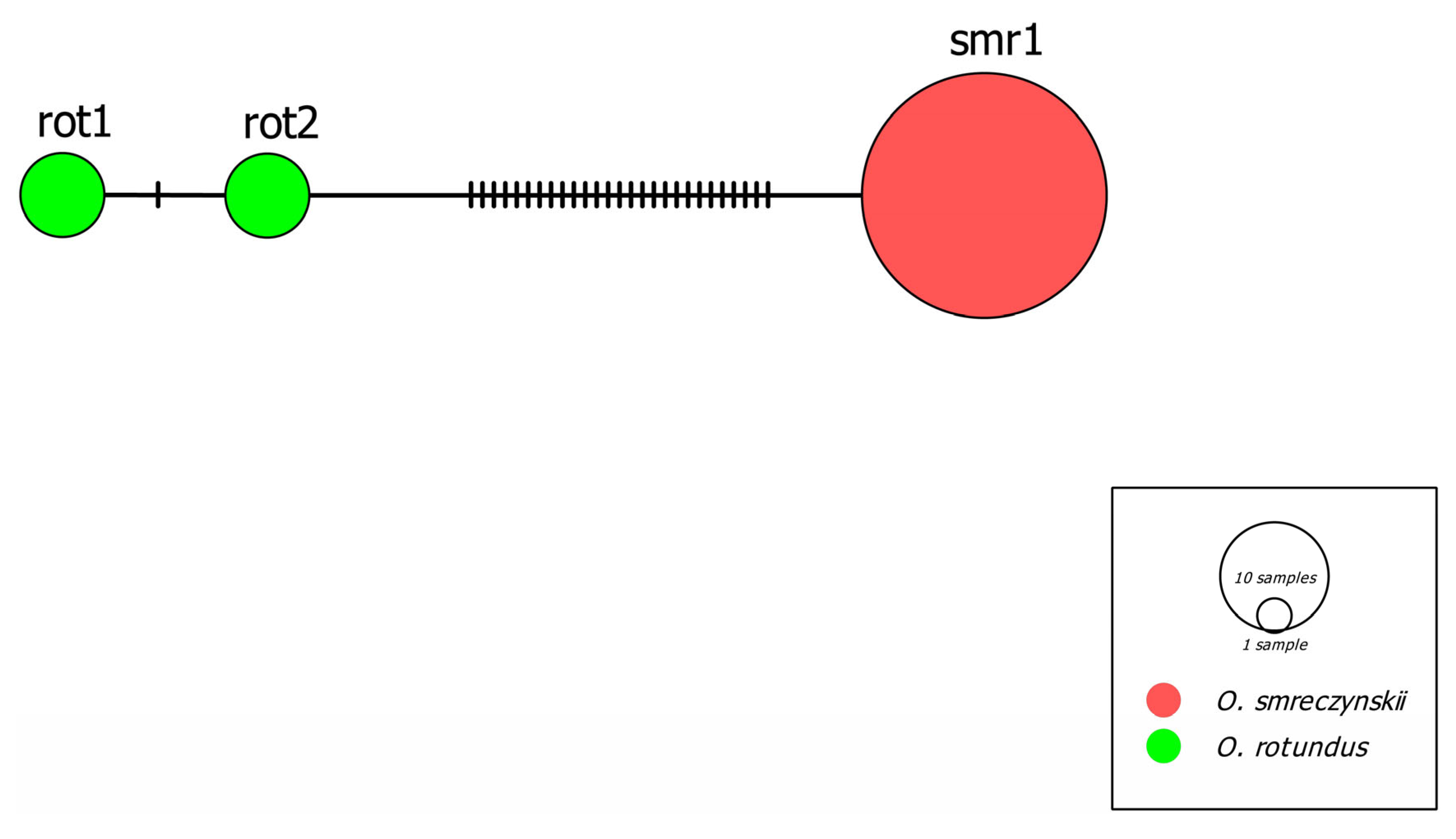
3.1. Molecular Study
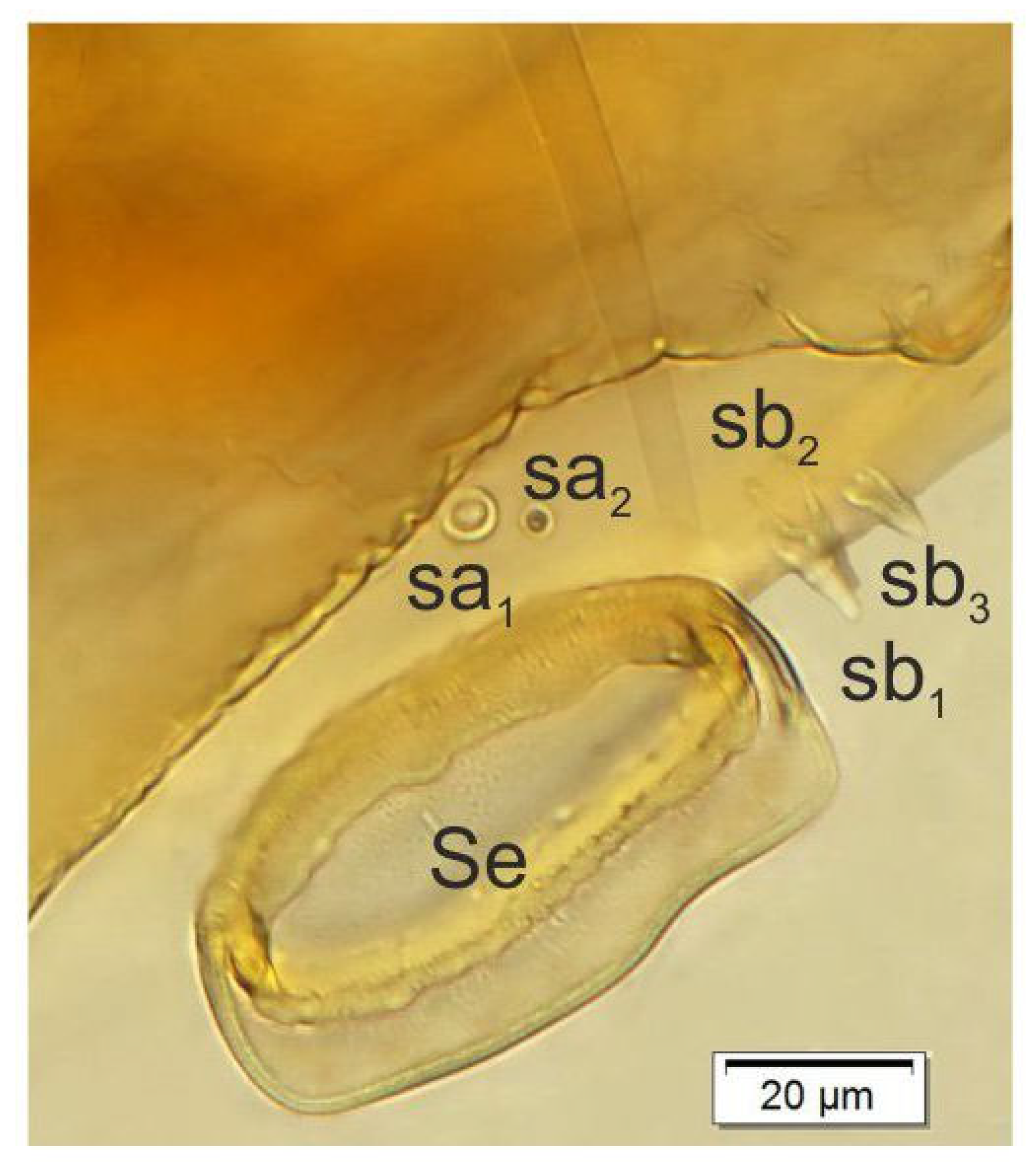
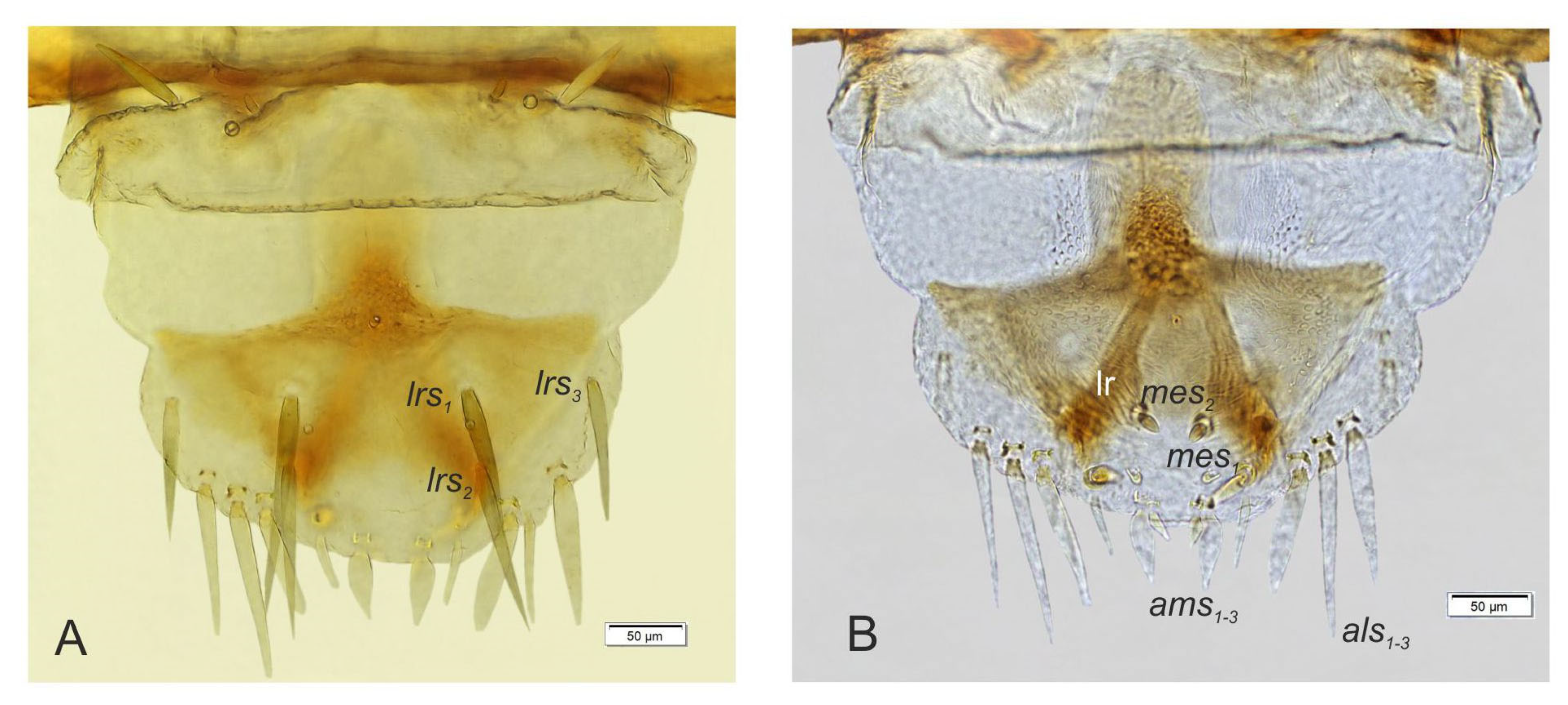
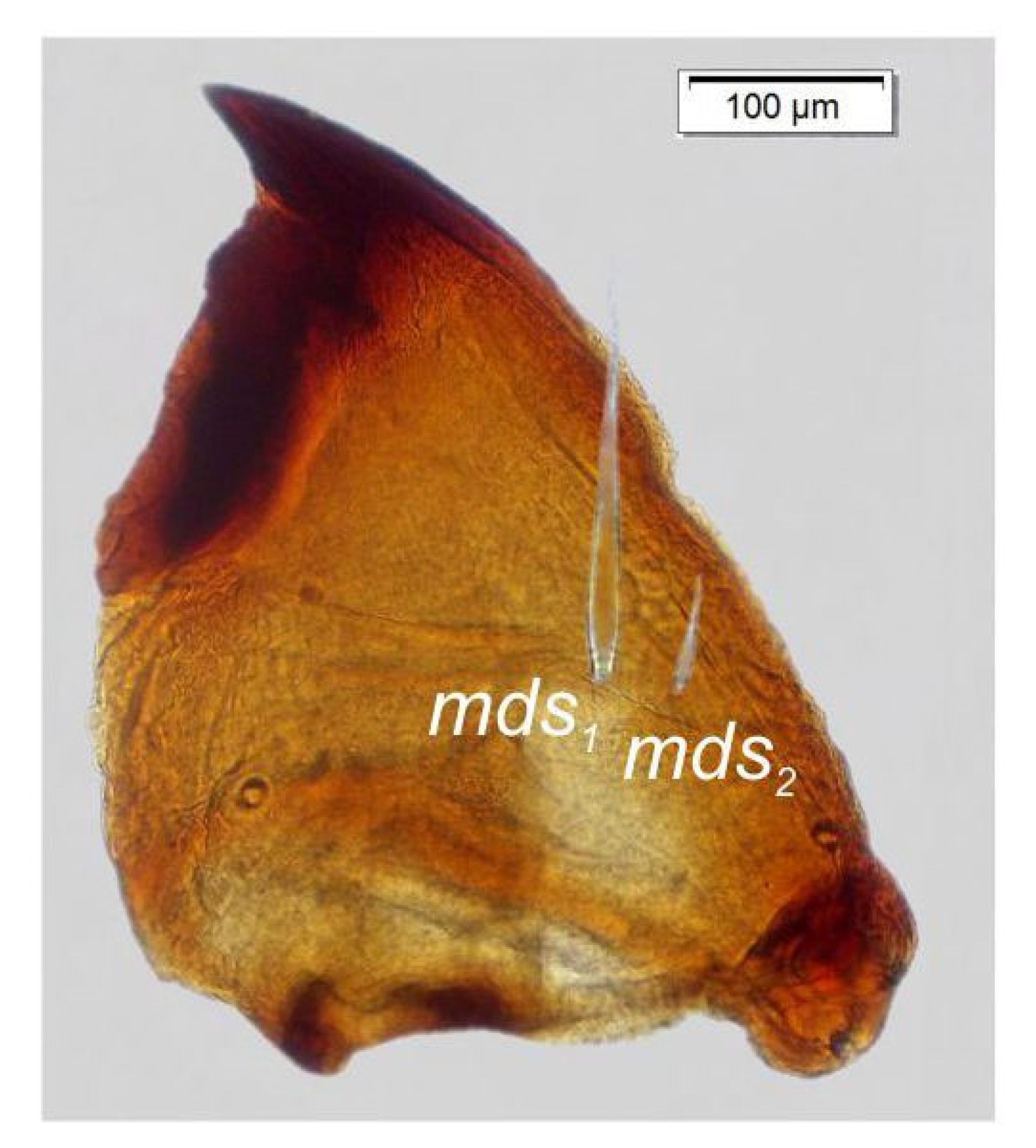
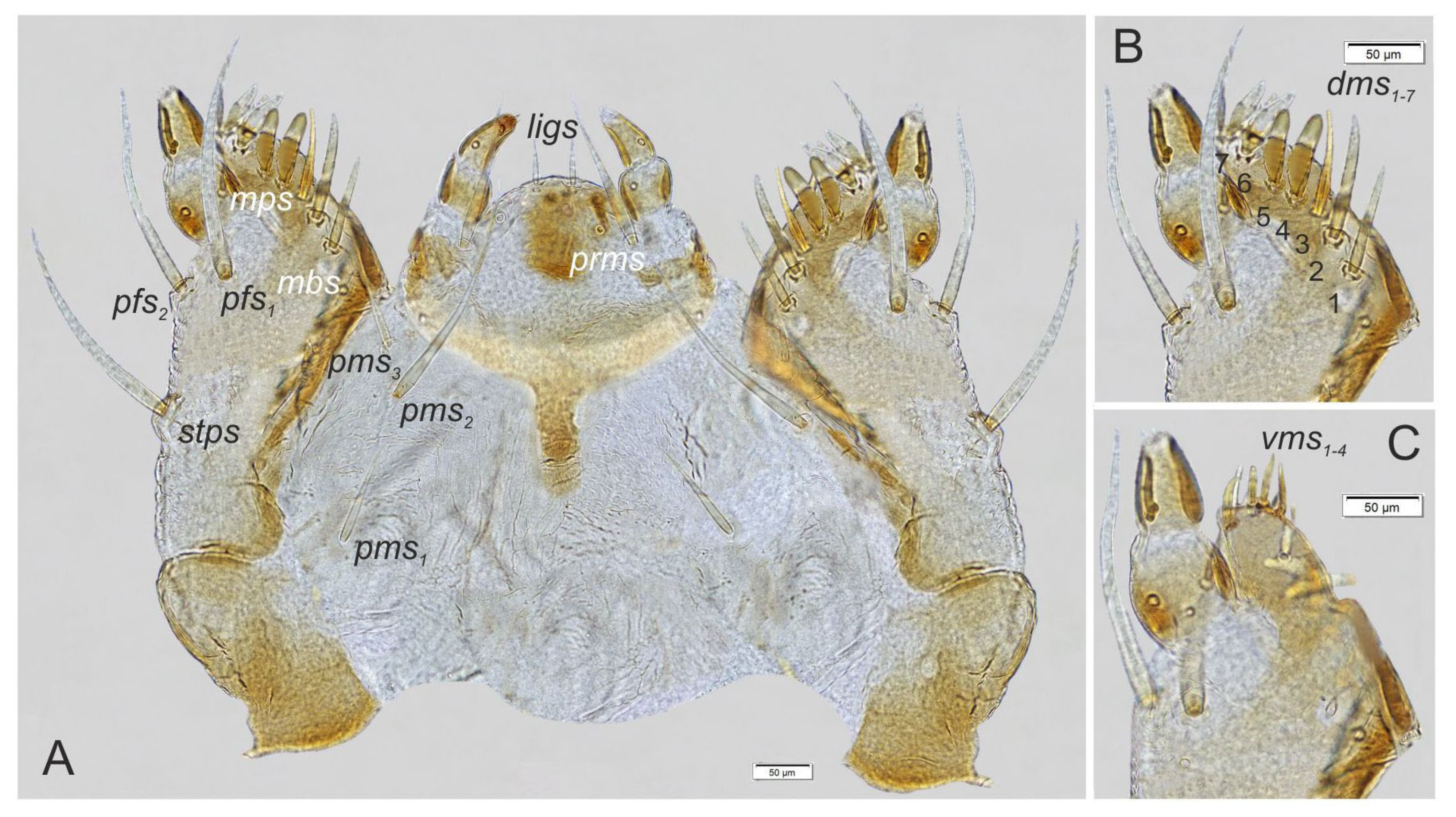
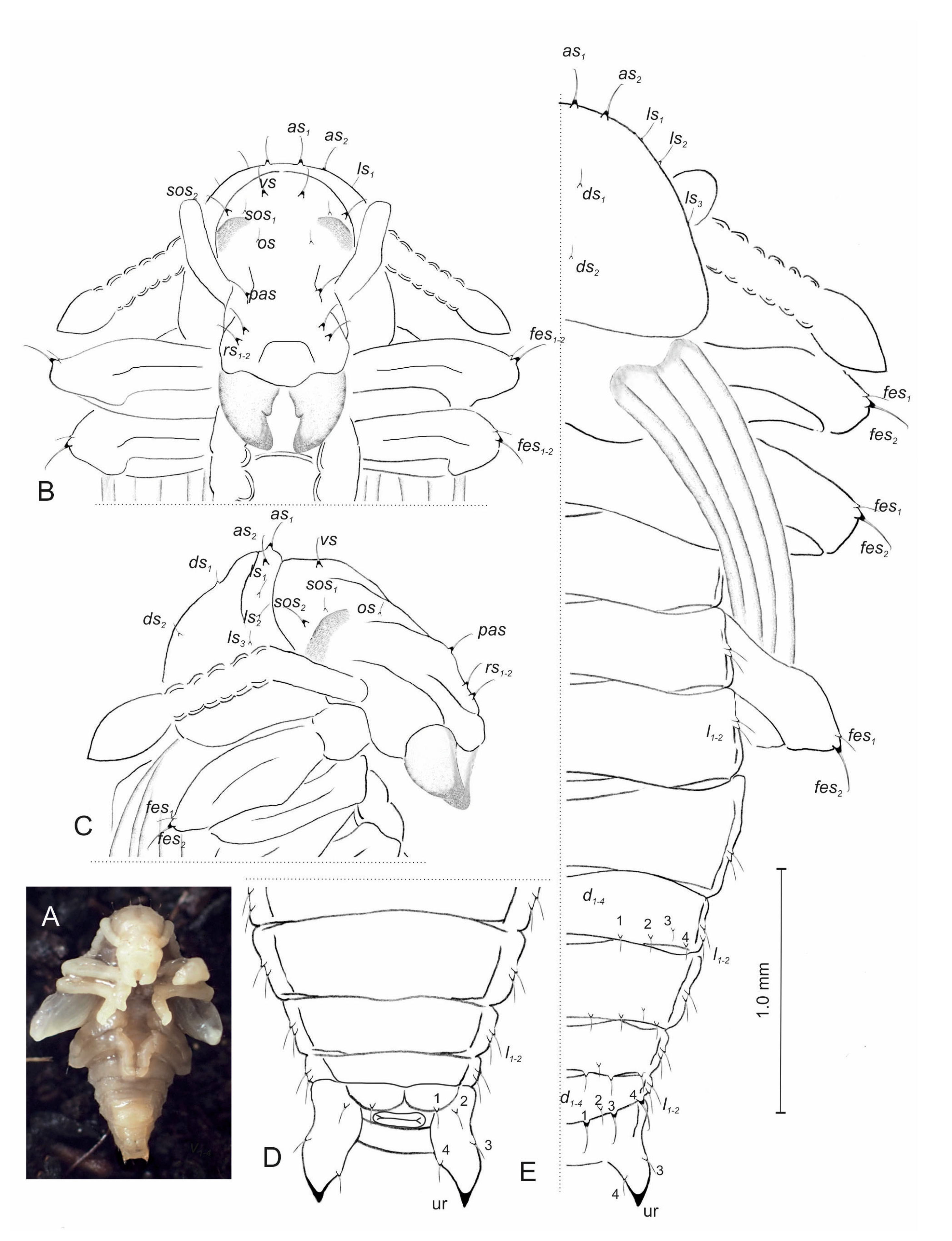
3.2. Description of Immature Stages of Otiorhynchus smreczynskii
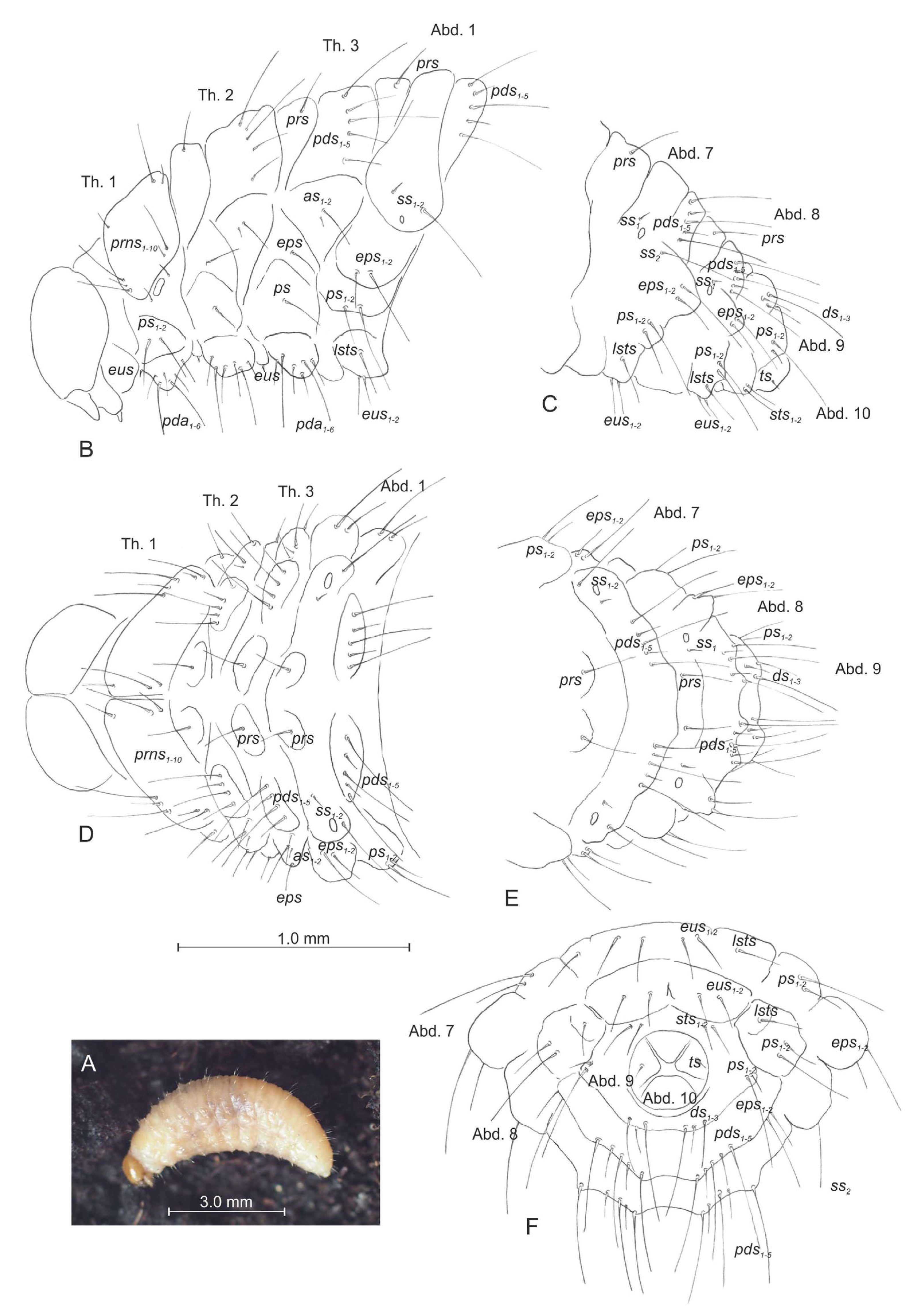
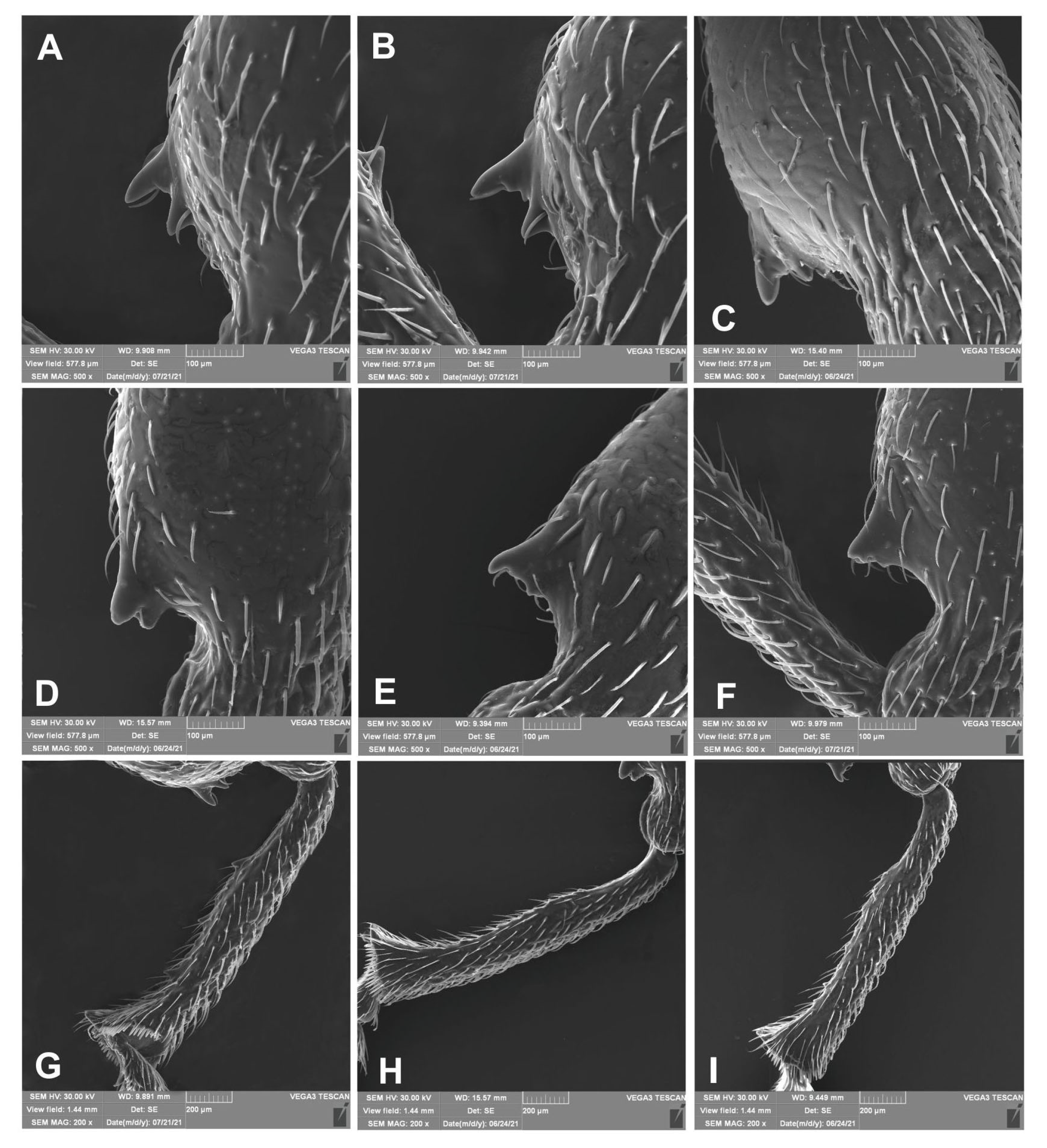
3.2.1. Larva (Figure 4A–F)
- First instar: head width: 0.2504, 0.2671, 0.2702
- Medium instars: head width: 0.7301, 0.7001
- Mature larva: head width: 0.9601, 1.0506, 1.134, 1.1361
- Body length: 6.0–6.5 mm
- Body width in the widest area (third abdominal segment): 1.98–2.20 mm
- Head width of mature larvae: 0.96–1.13 mm

3.2.2. Larval Instar Identification
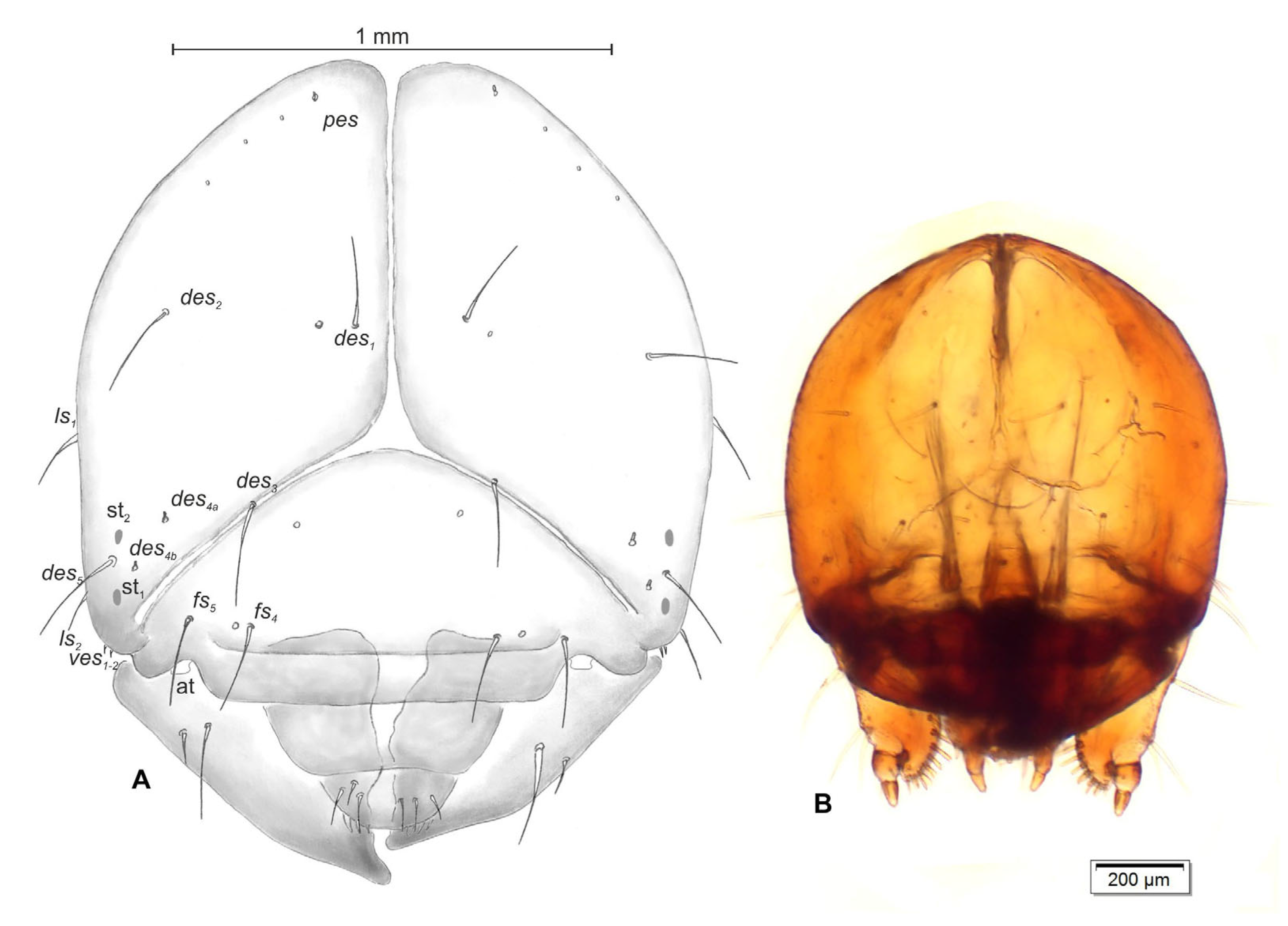
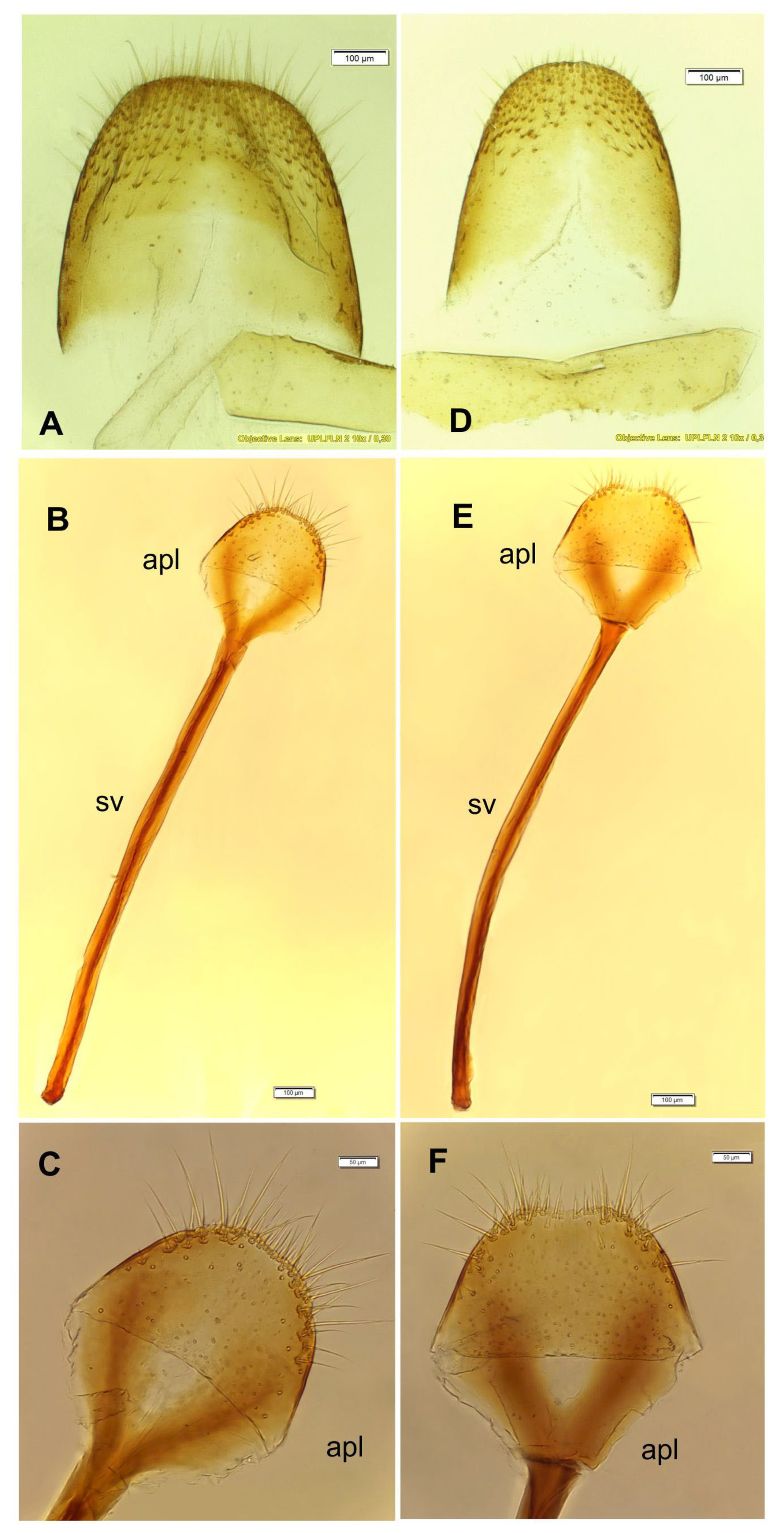
3.2.3. Pupa (Figure 10A–D)

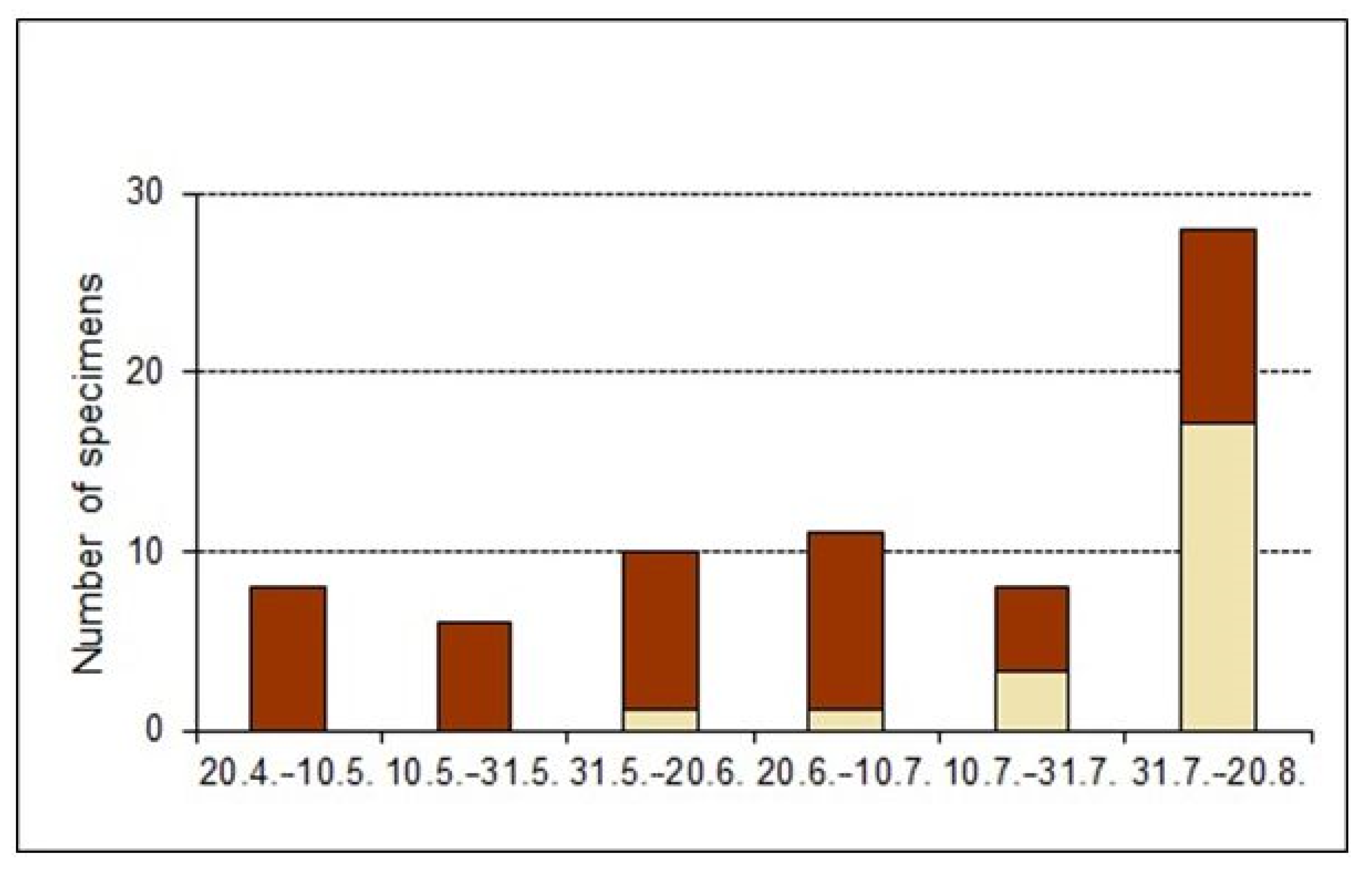
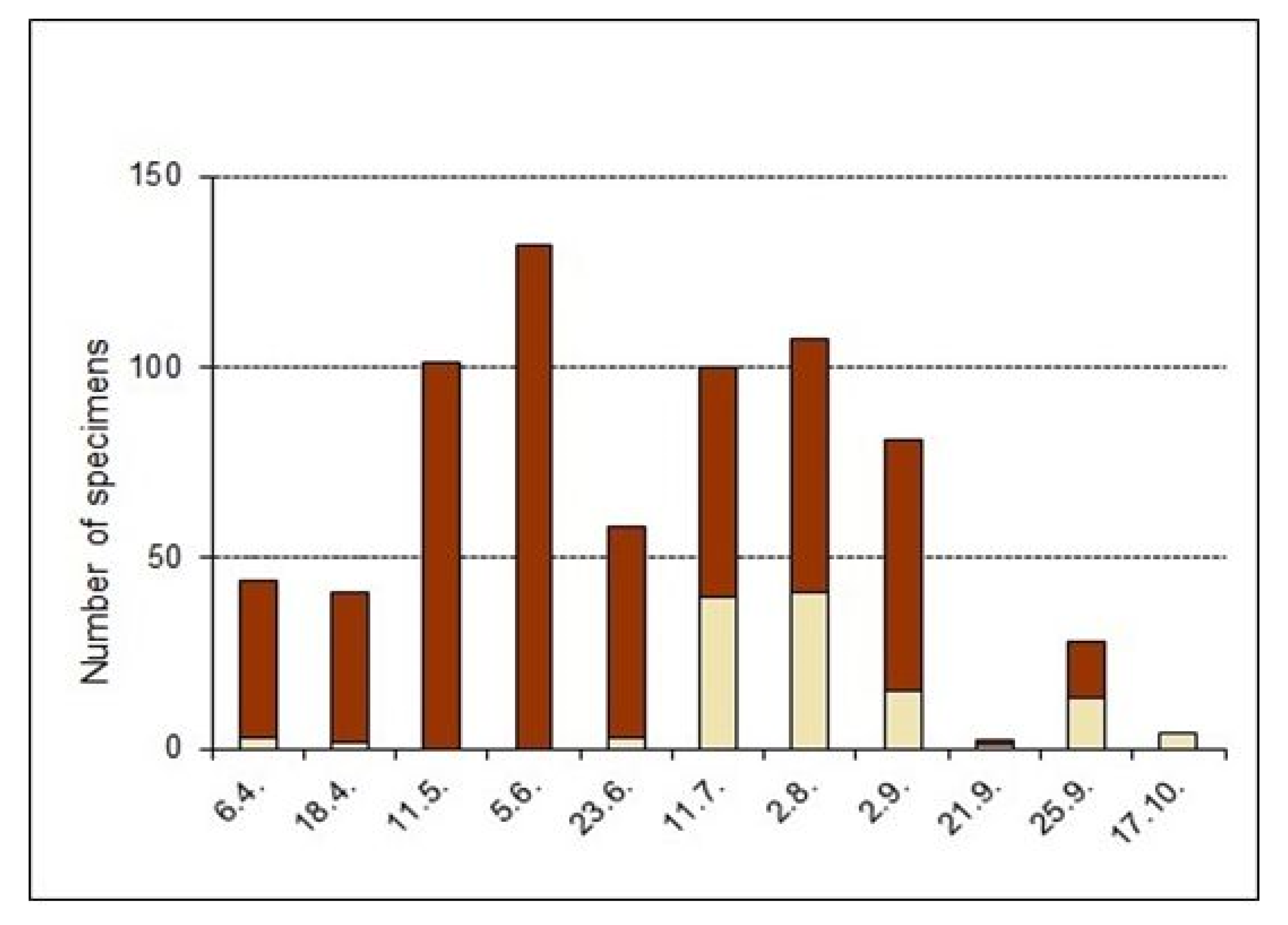
3.3. Biology: Development and Phenology
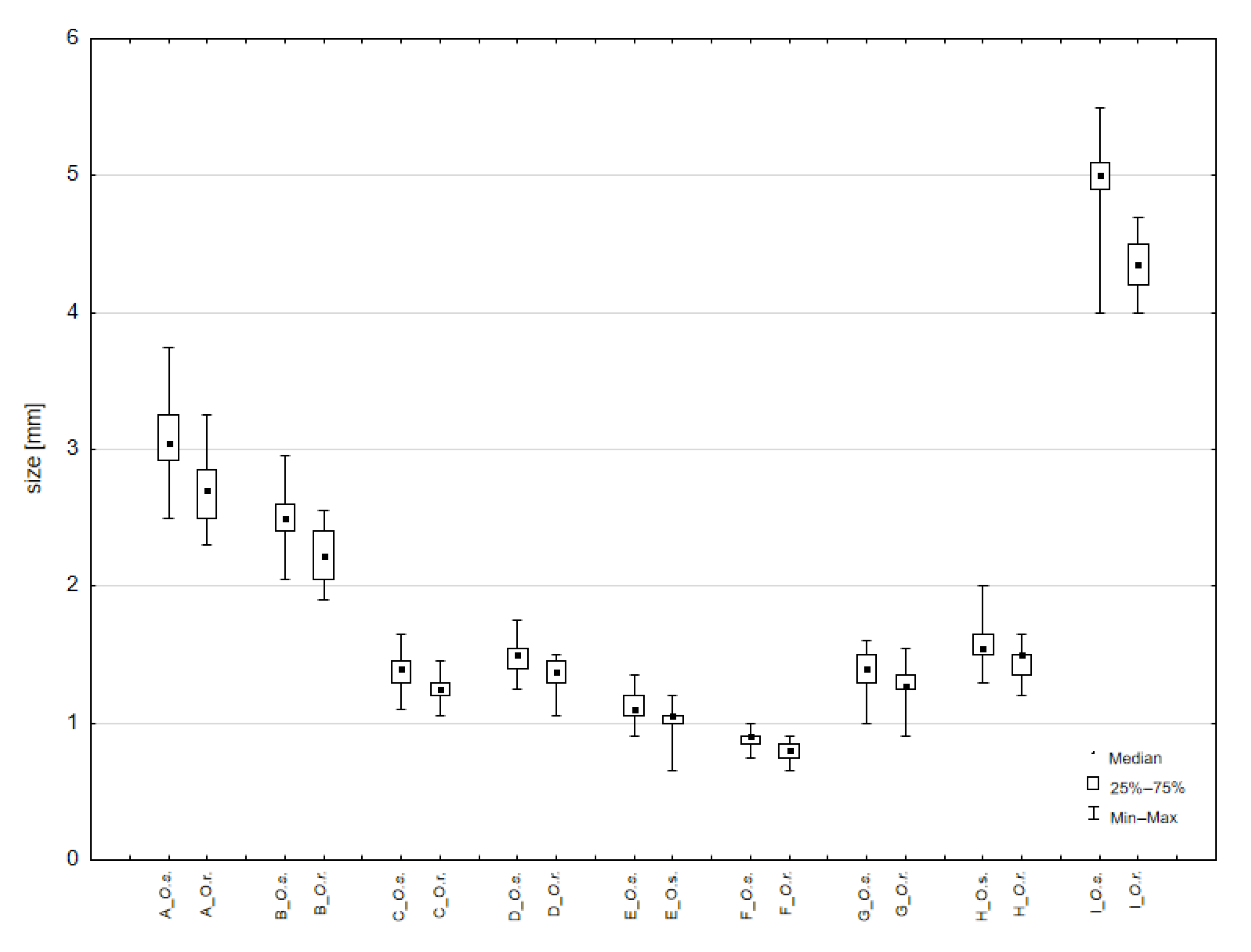
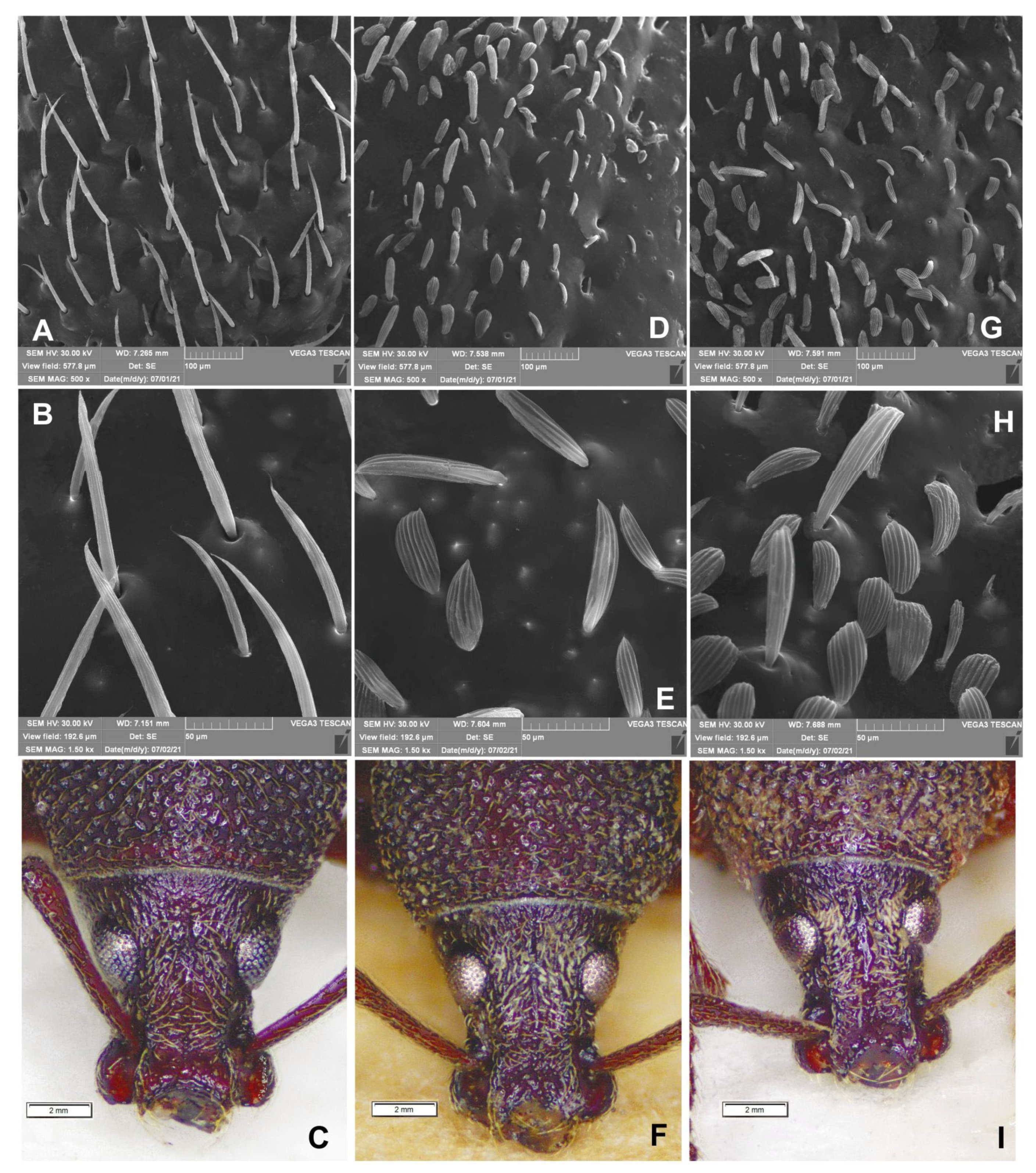
3.4. Comparison of External Morphology in Adults of Otiorhynchus smreczynskii and O. rotundus
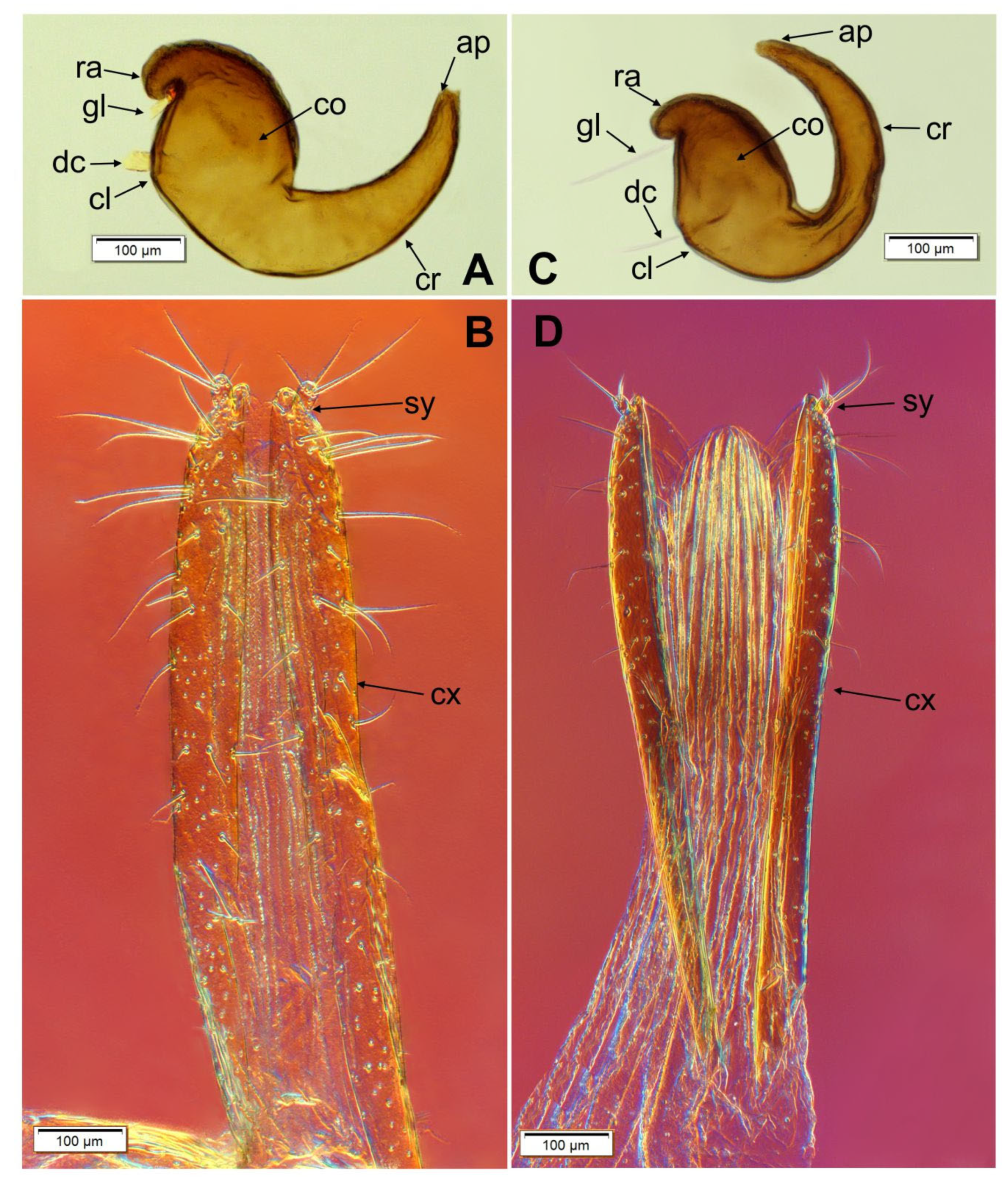
3.5. Description of Female Genitalia of Otiorhynchus smreczynskii and O. rotundus
4. Discussion
4.1. Morphology and Species Identification
4.2. Biology
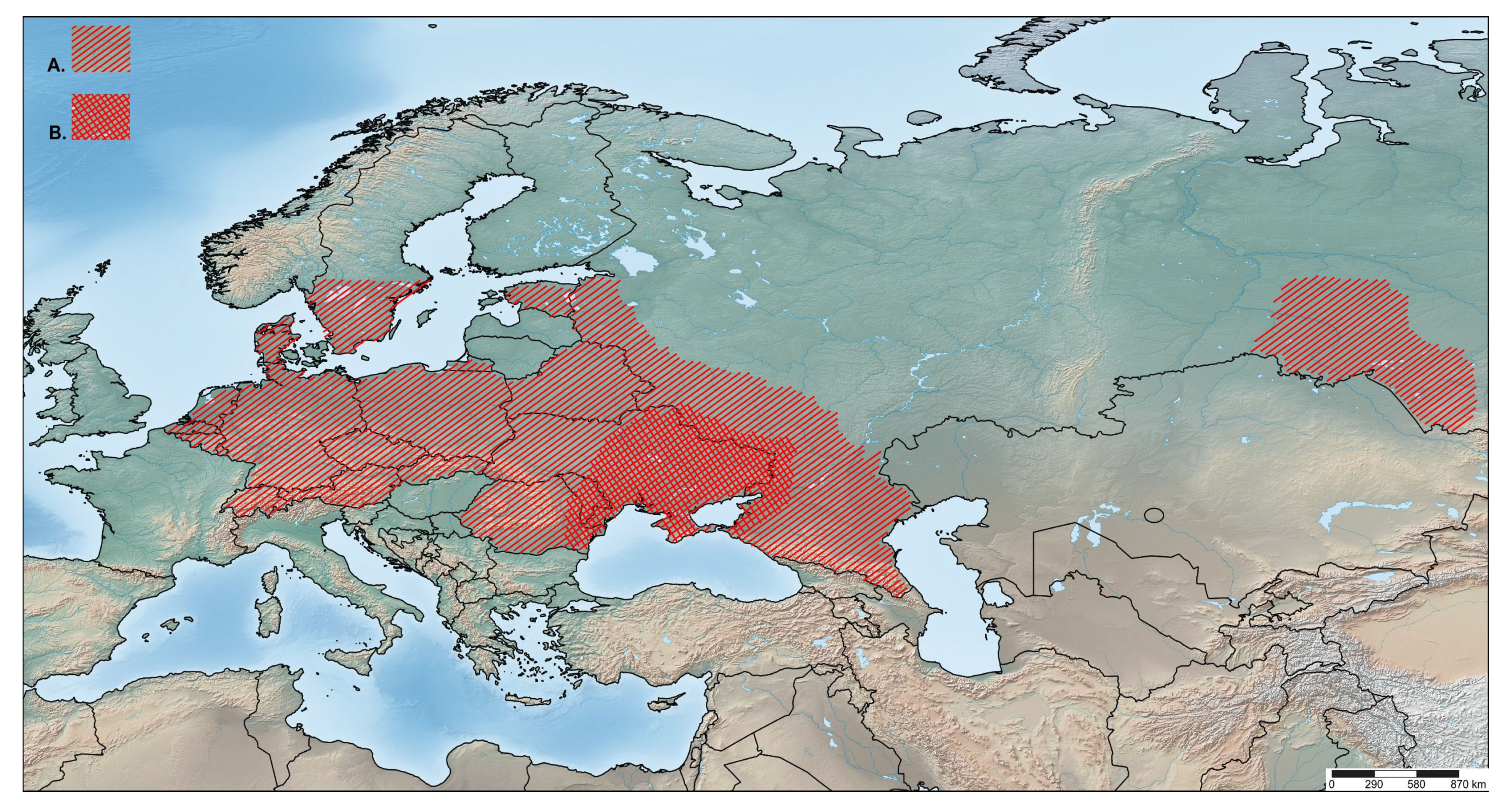
4.3. Origin and Distribution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cmoluch, Z. Otiorhynchus (Tournieria) smreczynskii sp.n. (Coleoptera Curculionidae). Bull. L’académie Pol. Sci. 1968, 16, 25–27. [Google Scholar]
- Reitter, E. Übersicht der Untergattungen und Artengruppen des Genus Otiorrhynchus Germ. Wien. Entomol. Ztg. 1912, 31, 45–67. [Google Scholar]
- Reitter, E. Curculionidae, 21. Teil: Die gezähnten Arten der Gattung Otiorrhynchus Germ.: Dorymerus und Tournieria. Verh. Nat. Forsch. Ver. Brünn 1914, 52, 129–251. [Google Scholar]
- Löbl, I.; Smetana, A. Catalogue of Palaearctic Coleoptera. In Curculionoidea II; Brill: Leiden, The Netherlands; Boston, MA, USA, 2013; Volume 8, 700p. [Google Scholar]
- Alonso-Zarazaga MABarrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.H.C.; Machado, A.; et al. Cooperative catalogue of Palaearctic Coleoptera Curculionoidea; Monografías Electrónicas SEA; Sociedad Entomológica Aragonesa S.E.A.: Zaragoza, Spain, 2017; Volume 8, 729p. [Google Scholar]
- Smreczyński, S. Ryjkowce (Curculionidae): Podrodzina Curculionidae i uzupełnienie do zeszytów 98a–e. In Klucze do Oznaczania Owadów Polski; Państwowe Wydawnictwo Naukowe: Warszawa, Poland,, 1976; 115p. [Google Scholar]
- Dieckmann, L. Beiträge zur Insektenfauna der DDR: Coleoptera—Curculionidae (Brachycerinae, Otiorhynchinae, Brachyderinae). Beiträge Entomol. 1980, 30, 145–310. [Google Scholar]
- Korotyaev, B.A.; Kataev, B.M.; Kovalev, A.V. On records of three additional species of the weevil genus Otiorhynchus Germ. (Coleoptera, Curculionidae: Entiminae) on lilacs (Syringa L.) in St. Petersburg. Entomol. Obozr. 2018, 97, 93–101. (In Russian) [Google Scholar] [CrossRef]
- Cmoluch, Z. Taxonomischer Wert des Spiculum ventrale von Weibchen der Arten in der Untergattung Tournieria Stierl. (Otiorhynchus Germ., Curculionidae, Coleoptera). Ann. UMCS Sec. C 1969, XXIV, 259–267. [Google Scholar]
- Frieser, R. 7. Unterfamilie: Otiorhynchinae. In Die Käfer Mitteleuropas; Freude, H., Harde, K.W., Lohse, G.A., Eds.; Bruchidae, Anthribidae, Scolytidae, Platypodidae & Curculionidae; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 1981; Volume 10, pp. 184–240. [Google Scholar]
- Stüben, P.E.; Schütte, A.; Bayer, C.; Astrin, J.J. The Molecular Weevil Identification project (Coleoptera: Curculionoidea), Part II. Towards an integrative taxonomy. In SNUDEBILLER: Studies on Taxonomy, Biology and Ecology of Curculionoidea; Curculio–Institute: Mönchengladbach, Germany, 2015; No. 237; 294p. [Google Scholar]
- Sprick, P.; Stüben, P.E.; Bayer, C. Digital-Weevil-Determination. Digitaler Bildbestimmungsschlüssel für bodenbewohnende Rüsselkäfer (Curculionidae: Entiminae) aus Baumschulen, Hopfengärten, Staudengärtnereien, Garten- und Parkanlagen. In Rüsselkäfer in Anthropogenen Lebensräumen; Sprick, P., Stüben, P.E., Eds.; Curculio–Institute: Mönchengladbach, Germany, 2012. [Google Scholar]
- Sprick, P. Bodenrüssler als Schädlinge im Garten– und Hopfenbau: Erfassung des Artenspektrums und Untersuchungen zu Bekämpfungsmöglichkeiten als Grundlage zur Entwicklung gezielter Bekämpfungsstrategien: Abschlussbericht. In Rüsselkäfer in Anthropogenen Lebensräumen; Sprick, P., Stüben, P.E., Eds.; Curculio–Institute: Mönchengladbach, Germany, 2012; 66p. [Google Scholar]
- Germain, J.F.; Chatot, C.; Meusnier, I.; Artige, E.; Rasplus, J.Y.; Cruaud, A. Molecular identification of Epitrix potato flea beetles (Coleoptera: Chrysomelidae) in Europe and North America. Bull. Entomol. Res. 2013, 103, 354–362. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2011, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data: III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- May, B.M. An introduction to the immature stages of Australian Curculionoidea. In Australian Weevils; Brentidae, Eurhynchidae, Apionidae and a chapter on immature stages by Brenda May; Zimmerman, E.C., Ed.; CSIRO: Melbourne, Australia, 1994; Volume II, 755p. [Google Scholar]
- Hille Ris Lambers, D. On mounting aphids and other soft–skinned insects. Entomol. Ber. 1950, 13, 55–58. [Google Scholar]
- Marvaldi, A.E. Higher level phylogeny of Curculionidae (Coleoptera: Curculionoidea) based mainly on larval characters, with special reference to broad–nosed weevils. Cladistics 1997, 13, 285–312. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Antennae and Sensilla. In Comprehensive Insect Physiology, Chemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 1–69. [Google Scholar]
- Howden, A.T. Structures related to oviposition in Curculionoidea. Mem. Entomol. Soc. Wash. 1995, 14, 53–102. [Google Scholar]
- Velázquez de Castro, A.J. Estudio Morfológico y Taxonómico del Género Sitona Germar, 1817 (Coleoptera, Curculionidae). Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 1997. Available online: http://eprints.ucm.es/4193/ (accessed on 25 March 2023).
- Gaiger, F.; Vanin, S.A. On the utility of female genitalia in the systematics of Curculionidae: Examples from Entimini and Hylobiina (Coleoptera: Curculionidae). Ann. Société Entomol. Fr. 2008, 44, 47–57. [Google Scholar] [CrossRef]
- Leibee, G.L.; Pass, B.C.; Yeargan, K.V. Instar determination of Clover Root Curculio, Sitona hispidulus (Coleoptera: Curculionidae). J. Kans. Entomol. Soc. 1980, 53, 473–475. [Google Scholar]
- Sprick, P.; Gosik, R. Biology and morphology of the mature larva of Mitoplinthus caliginosus caliginosus (Curculionidae, Molytinae). In SNUDEBILLER: Studies on Taxonomy, Biology and Ecology of Curculionoidea; Curculio–Institute: Mönchengladbach, Germany, 2014; No. 229; 10p. [Google Scholar]
- Gosik, R.; Sprick, P.; Morris, M.G. Descriptions of immature stages of four species of the genera Graptus, Peritelus, Philopedon, and Tanymecus and larval instar determination in Tanymecus (Coleoptera, Curculionidae, Entiminae). ZooKeys 2019, 813, 111–150. [Google Scholar] [CrossRef] [PubMed]
- von Lengerken, H. Lebensweise und Entwicklung des Fliederschädlings Otiorrhynchus rotundatus Siebold. Z. Angew. Entomol. 1919, 5, 67–83. [Google Scholar] [CrossRef]
- von Lengerken, H. Neues über die Lebensweise von Otiorrhynchus rotundatus Siebold. Z. Angew. Entomol. 1919, 5, 319–321. [Google Scholar]
- Scherf, H. Die Entwicklungsstadien der mitteleuropäischen Curculioniden (Morphologie, Bionomie, Ökologie). In Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main; Waldemar Kramer: Frankfurt am Main, Germany, 1964; Volume 506, pp. 1–336. [Google Scholar]
- Gosik, R.; Sprick, P.; Skuhrovec, J.; Deruś, M.; Hommes, M. Morphology and identification of the mature larvae of several species of the genus Otiorhynchus (Coleoptera, Curculionidae, Entiminae) from Central Europe with an update of the life history traits. Zootaxa 2016, 4108, 1–67. [Google Scholar] [CrossRef]
- Kangas, E.; Rutanen, I. Identification of females of the Finnish species of Altica Müller (Coleoptera, Chrysomelidae). Entomol. Fenn. 1993, 4, 115–128. [Google Scholar] [CrossRef] [Green Version]
- von Lengerken, H. Beitrag zur Lebensgewohnheit von Otiorrhynchus rotundatus Siebold. Z. Wiss. Insektenbiologie 1913, 9, 7–12. [Google Scholar]
- Richter, H. Der Fliederblattrüßler wandert nach Westen. Z. Pflanzenkrankh. Pflanzenschutz 1961, 68, 565–569. [Google Scholar]
- Germann, C. Otiorhynchus smreczynskii Cmoluch, 1968—Nun auch in der Schweiz (Coleoptera, Curculionidae, Entiminae). Mitt. Entomol. Ges. Basel 2006, 56, 122–126. [Google Scholar]
- Balalaikins, M.; Bukejs, A. Otiorhynchus smreczynskii (Coleoptera: Curculionidae)—A new to Estonia and Lithuania weevil species with notes on its occurrence and bionomy in the Eastern Baltic region. Acta Zool. Litu. 2011, 4, 263–267. [Google Scholar] [CrossRef]
- Heijerman, T.; Burgers, J. Otiorhynchus smreczynskii, weer een nieuwe snuitkeverexoot aangetroffen in Nederland (Coleoptera: Curculionidae). Entomol. Ber. 2015, 75, 137–141. [Google Scholar]
- Korotyaev, B.A.; Andreeva, S.V. On establishing of a weevil Otiorhynchus smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae) in St. Petersburg (Russia). Entomol. Rev. 2016, 96, 134–136. [Google Scholar] [CrossRef]
- Burkhardt, F. Zur Verbreitung und Lebensweise von Otiorrhynchus rotundatus Siebold. Z. Angew. Entomol. 1919, 5, 295–300. [Google Scholar] [CrossRef]
- Schulze, J. Otiorrhynchus rotundatus Sieb. Kleine Mitteilungen 1658. Entomol. Blätter Biol. Syst. Käfer 1959, 59, 60. [Google Scholar]
- Sprick, P. Untersuchungen über Otiorhynchus smreczynskii Cmoluch 1968, einen wenig bekannten Dickmaulrüßler, im Stadtgebiet von Hannover. Anz. Schädlingskunde Pflanzenschutz Umweltschutz 1989, 62, 47–50. [Google Scholar] [CrossRef]
- Cmoluch, Z.; Czarniawski, W. Otiorhynchus smreczynskii Cmoluch 1968, i gatunki ryjkowcowatych (Coleoptera, Attelabidae, Apionidae, Curculionidae) zbiorowisk synantropijnych Lublina. Folia Soc. Sci. Lub. 1992, 32, 63–69. [Google Scholar]
- Fägerström, C.; Kärnestam, E.; Anderson, R. New and expected weevils (Coleoptera: Otiorhynchini) on ornamental shrubs in Sweden. Entomol. Tidskr. 2010, 131, 37–48. [Google Scholar]
- Kolov, S.V.; Korotyaev, B.A. On establishing of two weevils (Coleoptera, Curculionidae: Entiminae) damaging fruit and berry crops in Southeastern Kazakhstan. Entomol. Rev. 2017, 97, 404–405. [Google Scholar] [CrossRef]
- Sautkin, F.V.; Meleshko, J.Y. First Records of the weevil Otiorhynchus smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae: Entiminae) in the Republic of Belarus. Entomol. Rev. 2016, 96, 866–872. [Google Scholar] [CrossRef]
- Sprick, P.; Stüben, P.E. Rüsselkäfer in anthropogenen Lebensräumen; Curculio–Institute: Mönchengladbach, Germany, 2012; 170 p. [Google Scholar]
- Kizub, I.V.; Leshchenko, M.V. Contribution to the knowledge of the genus Otiorhynchus Germar, 1822 (Coleoptera: Curculionidae) fauna of Ukraine: Part 3. Munis Entomol. Zool. 2020, 15, 397–406. [Google Scholar]
- Schuh, R.; Link, A.; Holzer, E. Bemerkenswerte Käferfunde aus Österreich (XVI) (Coleoptera). Koleopterol. Rundsch. 2009, 79, 321–326. [Google Scholar]
- Alekseev, V.I. Checklist of Curculionoidea (Insecta: Coleoptera) of the Kaliningrad region (Russia). Zool. Ecol. 2016, 26, 191–226. [Google Scholar] [CrossRef]
- Von Siebold, C.T.E. Beiträge zur Fauna der wirbellosen Thiere der Provinz Preussen. (Zehnter Beitrag). Die preußischen Käfer. (Fortsetzung und Schluß von S. 219). Neue Preußische Prov.Blätter 1847, 3, 419–451. [Google Scholar]
- Yunakov, N.N.; Nazarenko, V.; Filimonov, R.; Volovnik, S. A survey of the weevils of Ukraine (Coleoptera: Curculionoidea). Zootaxa 2018, 4404, 494. [Google Scholar] [CrossRef]
- Legalov, A.A. Annotated checklist of species of superfamily Curculionoidea (Coleoptera) from Asian part of the Russia. Amurian J. Zool. 2010, 2, 93–132. [Google Scholar] [CrossRef]
- Telnov, D. Check-List of Latvian Beetles (Insecta: Coleoptera) In Compendium of Latvian Coleoptera; Telnov, D., Ed.; Entomological Society of Latvia: Riga, Latvia, 2004; Volume 1, pp. 1–140. [Google Scholar]
- White, M.J.D. Animal Cytology and Evolution, 3rd ed.; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Lynch, M. The genetic structure of a cyclical parthenogen. Evolution 1984, 38, 186–203. [Google Scholar] [CrossRef]
- Stenberg, P.; Lundmark, M. Distribution, mechanisms and evolutionary significance of clonality and polyploidy in weevils. Agric. For. Entomol. 2004, 6, 259–266. [Google Scholar] [CrossRef]
- Stenberg, P.; Terhivuo, J.; Lokki, J.; Saura, A. Clone diversity in the polyploid weevil Otiorhynchus scaber. Hereditas 2000, 132, 137–142. [Google Scholar] [CrossRef]
- Mazur, M. Ryjkowce (Coleoptera: Nemonychidae, Attelabidae, Apionidae, Curculionidae) siedlisk kserotermicznych Polski II. Szczepanowice koło Miechowa. Wiad. Entomol. 2003, 22, 143–150. [Google Scholar]
- Cmoluch, Z. Badania nad fauną ryjkowców (Coleoptera, Curculionidae) roślinnych zespołów kserotermicznych południowo-wschodniej części Wyżyny Lubelskiej. Ann. UMCS Sec. C 1963, XVII, 1–75. [Google Scholar]
- Apfel, W.; Kopetz, A.; Weigel, A. Checkliste der Rüsselkäfer (Curculionoidea) Thüringens: 2. Fassung, Stand: November 2018. Checkl. Thüringer Insekten Spinn. 2018, 24, 9–40. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Group | Haplotypes | d1 (S.E) | d2 (S.E) | P (S.E.) | |
|---|---|---|---|---|---|
| 1 | 2 | ||||
| 1. O. rotundus | rot1-rot2 | 0.029 (0.005) | 0.002 (0.001) | - | (0.008) |
| 1. O. smreczynskii | smr1 | n/c (n/c) | 0.043 | - | |
| Instar | Specimens | Source | |
|---|---|---|---|
| Egg | 0.435 × 0.571 | 11 | own data |
| L1 | 0.258 | 7 | own data |
| Unknown instar | 0.715 | 2 | own data |
| Mature larva | 1.076 | 12 | own data |
| Pupa | 0.985 | 6 | own data |
| Adult | 1.015 | 24 | own data |
| Feature | Species | ||||
|---|---|---|---|---|---|
| O. smreczynskii | O. rotundus | ||||
| Holotype | Others | Females | Males | ||
| a | length of elytra | 3.35 | 2.50–3.75 | 2.30–3.25 | 2.30–2.75 |
| b | width of elytra | 2.80 | 2.05–2.95 | 2.00–2.55 | 1.90–2.25 |
| c | length of pronotum | 1.40 | 1.10–1.65 | 1.15–1.45 | 1.05–1.40 |
| d | width of pronotum | 1.70 | 1.25–1.75 | 1.15–1.50 | 1.05–1.45 |
| e | length of head and rostrum | 1.25 | 0.90–1.35 | 0.65–1.20 | 0.85–1.20 |
| f | width of head | 0.95 | 0.75–1.00 | 0.70–0.90 | 0.65–0.85 |
| g | length of tibia | 1.50 | 1.00–1.60 | 1.00–1.55 | 0.90–1.35 |
| h | length of funiculus and clava | 1.60 | 1.30–2.00 | 1.20–1.65 | 1.20–1.50 |
| i | length of body | 5.50 | 4.00–5.50 * | 4.10–4.70 | 4.00–4.50 |
| Case | Z | p |
|---|---|---|
| abdomen length (A) | 5.4 | <0.05 |
| abdomen width (B) | 5.64 | <0.05 |
| thorax length (C) | 4.51 | <0.05 |
| thorax width (D) | 3.94 | <0.05 |
| head length (E) | 4.05 | <0.05 |
| head width (F) | 5.07 | <0.05 |
| fore tibia length (G) | 3.14 | <0.05 |
| antenna length (H) | 4.39 | <0.05 |
| length of the body (I) | 6.72 | <0.05 |
| A/B | 1.49 | n.s. |
| C/D | 1.25 | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gosik, R.; Sprick, P.; Wrzesień, M.; Dzyr, A.; Krstić, O.; Toševski, I. Developmental Biology and Identification of a Garden Pest, Otiorhynchus (Podoropelmus) smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae, Entiminae), with Comments on Its Origin and Distribution. Insects 2023, 14, 360. https://doi.org/10.3390/insects14040360
Gosik R, Sprick P, Wrzesień M, Dzyr A, Krstić O, Toševski I. Developmental Biology and Identification of a Garden Pest, Otiorhynchus (Podoropelmus) smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae, Entiminae), with Comments on Its Origin and Distribution. Insects. 2023; 14(4):360. https://doi.org/10.3390/insects14040360
Chicago/Turabian StyleGosik, Rafał, Peter Sprick, Małgorzata Wrzesień, Agata Dzyr, Oliver Krstić, and Ivo Toševski. 2023. "Developmental Biology and Identification of a Garden Pest, Otiorhynchus (Podoropelmus) smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae, Entiminae), with Comments on Its Origin and Distribution" Insects 14, no. 4: 360. https://doi.org/10.3390/insects14040360
APA StyleGosik, R., Sprick, P., Wrzesień, M., Dzyr, A., Krstić, O., & Toševski, I. (2023). Developmental Biology and Identification of a Garden Pest, Otiorhynchus (Podoropelmus) smreczynskii Cmoluch, 1968 (Coleoptera, Curculionidae, Entiminae), with Comments on Its Origin and Distribution. Insects, 14(4), 360. https://doi.org/10.3390/insects14040360