
Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Investigation on Wheat Mites
2.2. Specimen Collection
2.3. DNA Extraction and Mitochondrial COI Gene Amplification
2.4. Data Analysis
3. Results
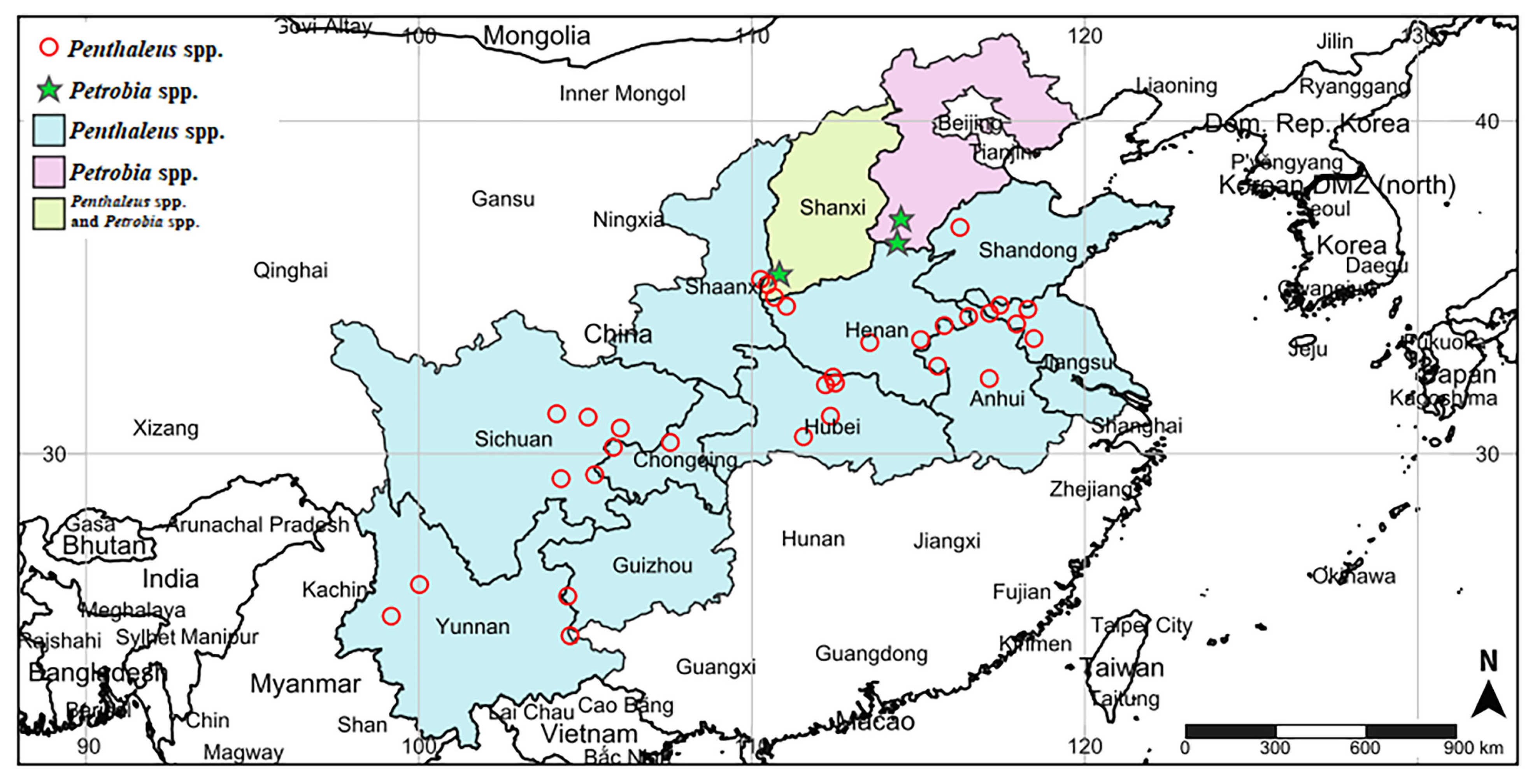
3.1. Distribution Pattern of Wheat Pest Mites
3.2. Population Distribution of Blue Oat Mites
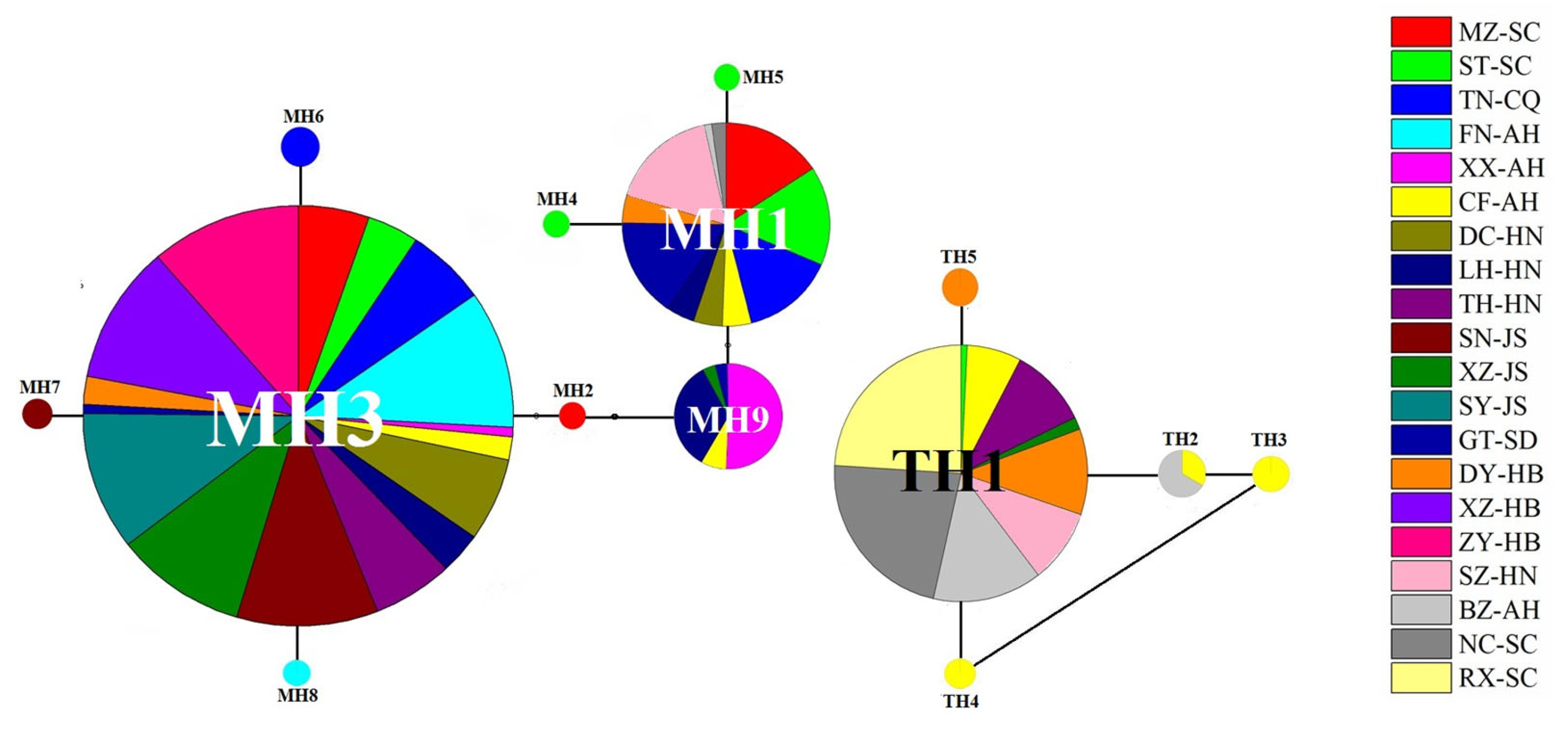
3.3. Haplotype Diversity, Nucleotide Diversity, and Haplotype distribution
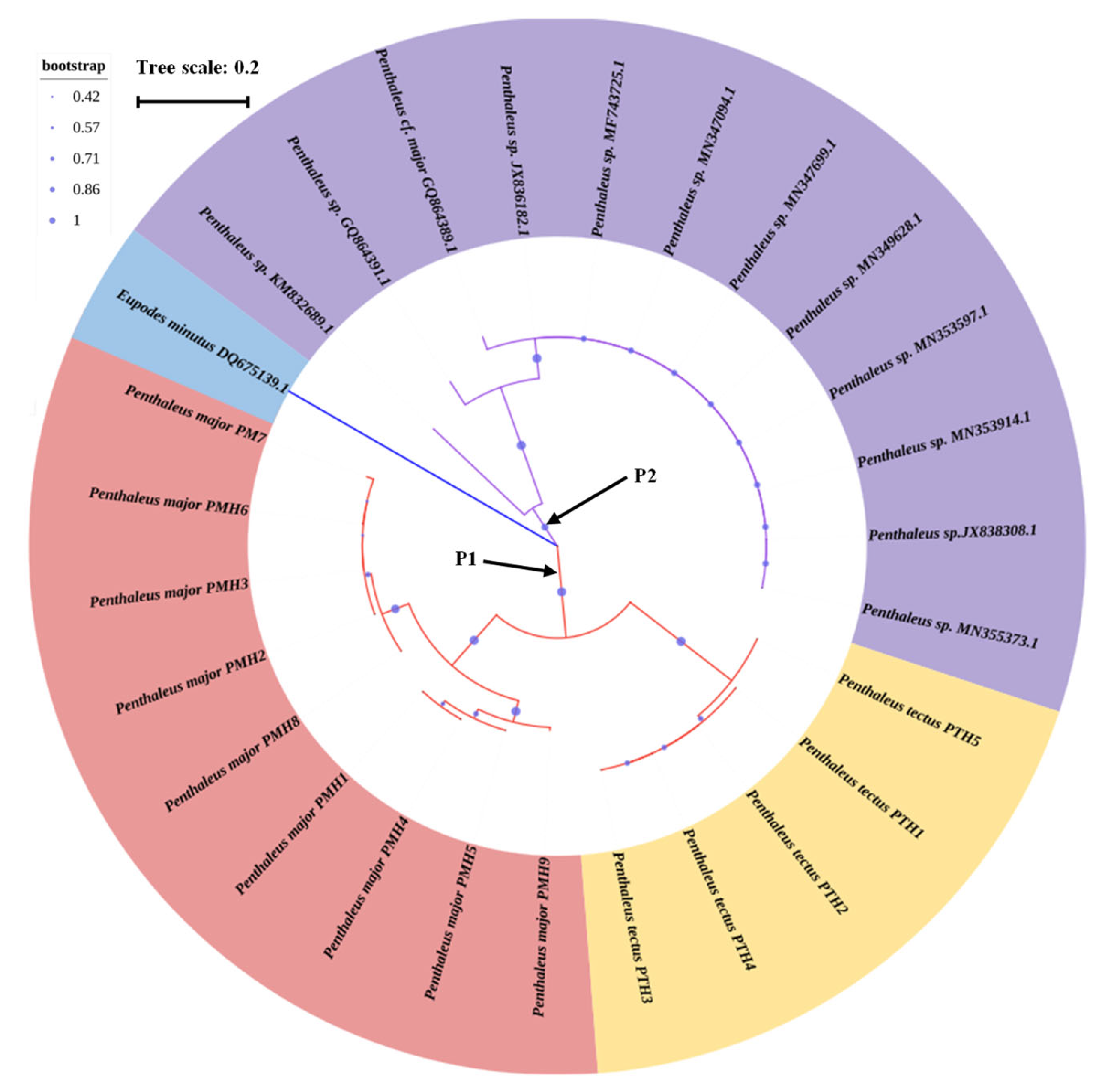
3.4. Phylogenetic Relationship and Haplotype Network
3.5. Population Differentiation and Genetic Structure
3.6. Demographic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chada, H.L. Biology of the winter grain mite and its control in small grains. J. Econ. Entomol. 1956, 49, 515–520. [Google Scholar] [CrossRef]
- Narayan, D.S. Morphological, biological and ecological studies on the winter grain mite, Penthaleus major (Dugés), Penthaleidae; Acarina. Part 1. J. Zool. Soc. India 1962, 14, 45–63. [Google Scholar]
- Wallace, M.M.H.; Mahon, J.A. The distribution of Halotydeus destructor and Penthaleus major (Acari: Eupodidae) in Australia in relation to climate and land use. Aust. J. Zool. 1971, 19, 65–76. [Google Scholar] [CrossRef]
- Robinson, M.T.; Hoffmann, A.A. The pest status and distribution of three cryptic blue oat mite species (Penthaleus spp.) and redlegged earth mite (Halotydeus destructor) in southeastern Australia. Exp. Appl. Acarol. 2001, 25, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Umina, P.A.; Hoffmann, A.A.; Weeks, A.R. Biology, ecology and control of the Penthaleus species complex (Acari: Penthaleidae). Exp. Appl. Acarol. 2004, 34, 211–237. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.A.H.; Clarke, M.B.; Ronning, D.A. Estimating invertebrate pest losses in six major Australian grain crops. Aust. J. Entomol. 2013, 52, 227–241. [Google Scholar] [CrossRef]
- Qin, T.K.; Halliday, R.B. The Australian species of Chromotydaeus Berlese and Penthaleus C. L. Koch (Acarina, Penthaleidae). J. Nat. Hist. 1996, 30, 1833–1848. [Google Scholar] [CrossRef]
- Qin, T.K.; Halliday, R.B. Revision of the Australian and South African species of Halotydeus (Acarina: Penthaleidae). Bull. Entomol. Res. 1996, 86, 441–450. [Google Scholar] [CrossRef]
- Womersley, H. On the name of the “blue oat mite” of Australia. Bull. Entomol. Res. 1935, 26, 163. [Google Scholar] [CrossRef]
- Weeks, A.R.; Fripp, Y.J.; Hoffmann, A.A. Genetic structure of Halotydeus destructor and Penthaleus major populations in Victoria (Acari: Penthaleidae). Exp. Appl. Acarol. 1995, 19, 633–646. [Google Scholar] [CrossRef]
- Weeks, A.R.; Hoffmann, A.A. The biology of Penthaleus species in southeastern Australia. Entomol. Exp. Appl. 1999, 92, 179–189. [Google Scholar] [CrossRef]
- Halliday, R.B. Systematics and biology of Penthaleus tectus sp. n. (Acari: Penthaleidae), a recently discovered pest of grain crops in eastern Australia. Aust. J. Entomol. 2005, 44, 144–149. [Google Scholar] [CrossRef]
- Weeks, A.R.; Hoffmann, A.A. Intense selection of mite clones in a heterogeneous environment. Evolution 1998, 52, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.T.; Weeks, A.R.; Hoffmann, A.A. Geographic patterns of clonal diversity in the earth mite species Penthaleus major with particular emphasis on species margins. Evolution 2002, 56, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.R.; Hoffmann, A.A. Frequency-dependent selection maintains clonal diversity in an asexual organism. Proc. Natl. Acad. Sci. USA 2008, 105, 17872–17877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Allendorf, F.W.; Luikart, G. Conservation and the Genetics of Populations; Wiley: New York, NY, USA, 2009. [Google Scholar]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Vandewoestijne, S.; Schtickzelle, N.; Baguette, M. Positive correlation between genetic diversity and fitness in a large, well-connected metapopulation. BMC Biol. 2008, 6, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoa, N.P.; Hamzah, A.; Nguyen, N.H. Genetic variation and correlated changes in reproductive performance of a red tilapia line selected for improved growth over three generations. Anim. Reprod. Sci. 2017, 184, 94–101. [Google Scholar] [CrossRef]
- Carrazco-Montalvo, A.; Ponce, P.; Villota, S.D.; Quentin, E.; Muñoz-Tobar, S.; Coloma, J.; Cevallos, V. Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador. Insects 2022, 13, 305. [Google Scholar] [CrossRef]
- Pramual, P.; Jomkumsing, P.; Wongpakam, K.; Vaisusuk, K.; Chatan, W.; Gomontean, B. Population genetic structure and population history of the biting midge Culicoides mahasarakhamense (Diptera: Ceratopogonidae). Insects 2022, 13, 724. [Google Scholar] [CrossRef]
- Wang, J.Q. Acute symptoms identification methods and prevention methods of mite pests on wheat. Hortic. Seed 2017, 4, 53–55. [Google Scholar]
- He, X.Y. Occurrence and control technology of wheat main pests in Taihe county in 2020. Anhui Agric. Sci. Bull. 2021, 5, 115–117. [Google Scholar] [CrossRef]
- Hill, M.P.; Hoffmann, A.A.; Mccoll, S.A.; Umina, P.A. Distribution of cryptic blue oat mite species in Australia: Current and future climate conditions. Agric. For. Entomol. 2012, 14, 127–137. [Google Scholar] [CrossRef]
- Umina, P.A.; Hoffmann, A.A. Tolerance of cryptic species of blue oat mites (Penthaleus spp.) and the redlegged earth mite (Halotydeus destructor) to pesticides. Aust. J. Exp. Agric. 1999, 39, 621–628. [Google Scholar] [CrossRef]
- Wang, J. The life tables of laboratory population of Frankliniella occidentalis and Thrips tabaci on purple cabbage. Acta. Phytophyl. Sin. 2011, 27, 300–302. [Google Scholar]
- Li, X.; Zhang, Z.; Zhang, J.; Huang, J.; Wang, L.; Li, Y.; Hafeez, M.; Lu, Y. Population genetic diversity and structure of Thrips tabaci (Thysanoptera: Thripidae) on Allium hosts in China, inferred from mitochondrial COI gene sequences. J. Econ. Entomol. 2020, 113, 1426–1435. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Sudhir, K.; Glen, S.; Koichiro, T. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H. Arlequin suite ver3.5: A new series of programs to perform population genetics analyses under linux and windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slatkin, M.; Hudson, R.R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 1991, 129, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Goodall-Copestake, W.; Tarling, G.; Murphy, E. On the comparison of population-level estimates of haplotype and nucleotide diversity: A case study using the gene cox1 in animals. Heredity 2012, 109, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.T.; Hoffmann, A.A. Additional tests on the effects of pesticides on cryptic species of blue oat mite (Penthaleus spp.) and the redlegged earth mite (Halotydeus destructor). Aust. J. Exp. Agric. 2000, 40, 671–678. [Google Scholar] [CrossRef]
- Fox, J.A.; Dybdahl, M.F.; Jokela, J.; Lively, C.M. Genetic structure of coexisting sexual and clonal subpopulations in a freshwater snail (Potamopyrgus antipodarum). Evolution 1996, 50, 1541–1548. [Google Scholar]
- Herbert, P.D.N.; Crease, T.J. Clonal coexistence in Daphnia pulex (Leydig): Another planktonic paradox. Science 1980, 207, 1363–1365. [Google Scholar] [CrossRef]
- Turner, B.J.; Elder, J.F.J.; Laughlin, T.F.; Davis, W.P.; Taylor, D.S. Extreme clonal diversity and divergence in populations of a selfing hermaphroditic fish. Proc. Natl. Acad. Sci. USA 1992, 89, 10643–10647. [Google Scholar] [CrossRef] [Green Version]
- Vrijenhoek, R.C. Factors affecting clonal diversity and coexistence. Amer. Zool. 1979, 19, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Levene, H. Genetic equilibrium when more than one ecological niche is available. Am. Nat. 1953, 87, 331–333. [Google Scholar] [CrossRef]
- Strobeck, C. Sufficient conditions for polymorphism with N niches and M mating groups. Am. Nat. 1974, 108, 152–156. [Google Scholar] [CrossRef]
- Cao, L.J.; Gao, Y.F.; Gong, Y.J.; Chen, J.C.; Chen, M.; Hoffmann, A.; Wei, S.J. Population analysis reveals genetic structure of an invasive agricultural Thrips pest related to invasion of greenhouses and suitable climatic space. Evol. Appl. 2019, 12, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Cai, W.Z.; Luo, J.Y.; Zhang, S.; Li, W.D.; Wang, C.Y.; Lv, L.M.; Cui, J.J. Population genetic structure and expansion patterns of the cotton pest Adelphocoris fasciaticollis. J. Pest. Sci. 2018, 91, 539–550. [Google Scholar] [CrossRef]
- Zamoum, T.; Simon, J.C.; Crochard, D.; Ballanger, Y.; Lapchin, L.; Vanlerberghe-Masutti, F.; Guillemaud, T. Does insecticide resistance alone account for the low genetic variability of asexually reproducing populations of the peach-potato aphid Myzus persicae? Heredity 2005, 94, 630–639. [Google Scholar] [CrossRef]
- RidsdillSmith, T.J.; Annells, A.J. Seasonal occurrence and abundance of redlegged earth mite Halotydeus destructor (Acari: Penthaleidae) in annual pastures of southwestern Australia. Bull. Entomol. Res. 1997, 87, 413–423. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Pop. ID | Population | Longitude (°E) | Latitude (°N) | Number of P. major | Number of P. tectus |
|---|---|---|---|---|---|---|
| SW (SC, CQ) | RX-SC | Sichuan, Rongxian | 104°27′ | 29°25′ | 31 | 0 |
| MZ-SC | Sichuan, Mianzhu | 104°14′ | 31°20′ | 30 | 0 | |
| ST-SC | Sichuan, Santai | 105°08′ | 31°10′ | 27 | 1 | |
| NC-SC | Sichuan, Nanchong | 106°05′ | 30°77′ | 2 | 29 | |
| TN-CQ | Chongqing, Tongnan | 105°84′ | 30°19′ | 32 | 0 | |
| YV (HB) | DY-HB | Hubei, Dangyang | 111°55′ | 30°51′ | 10 | 16 |
| XZ-HB | Hubei, Xiangzhou | 112°21′ | 32°07′ | 30 | 0 | |
| ZY-HB | Hubei, Zaoyang | 112°51′ | 32°12′ | 32 | 0 | |
| ZX-HB | Hubei, Zhongxiang | 112°36′ | 31°13′ | 4 | 0 | |
| YH (JS, AH, HN, SD, SX) | SN-JS | Jiangsu, Suining | 117°95′ | 33°90′ | 31 | 0 |
| XZ-JS | Jiangsu, Xuzhou | 117°14′ | 34°22′ | 28 | 2 | |
| SY-JS | Jiangsu, Siyang | 118°47′ | 33°46′ | 30 | 0 | |
| DH-JS | Jiangsu, Donghai | 118°29′ | 34°34′ | 3 | 1 | |
| FN-AH | Anhui, Funan | 115°58′ | 32°63′ | 31 | 0 | |
| BZ-AH | Anhui, Bozhou | 115°78′ | 33°85′ | 1 | 20 | |
| CF-AH | Anhui, Changfeng | 117°13′ | 32°26′ | 13 | 13 | |
| XX-AH | Anhui, Xiaoxian | 116°51′ | 34°12′ | 29 | 0 | |
| DC-HN | Henan, Dancheng | 115°07′ | 33°43′ | 22 | 0 | |
| SZ-HN | Henan, Shanzhou | 111°04′ | 34°43′ | 17 | 12 | |
| TH-HN | Henan, Tanghe | 112°44′ | 32°30′ | 17 | 13 | |
| LH-HN | Henan, Luohe | 113°54′ | 33°34′ | 31 | 0 | |
| GT-SD | Shandong, Gaotang | 116°25′ | 36°80′ | 18 | 0 | |
| RC-SX | Shanxi, Ruicheng | 110°68′ | 34°70′ | 0 | 1 |
| Species | Length of Sequences (bp) | C | V | PI | S | A + T (%) |
|---|---|---|---|---|---|---|
| P. major | 657–658 | 636 | 22 | 19 | 3 | 70.30 |
| P. tectus | 657–658 | 654 | 4 | 4 | None | 69.90 |
| Species | Pop.ID | H | Haplotype (No. of Specimens) | Hd | Pi | K |
|---|---|---|---|---|---|---|
| P. major | MZ-SC | 3 | MH1 (14), MH2 (1), MH3 (15) | 0.549 | 0.014 | 9.274 |
| ST-SC | 4 | MH1 (14), MH3 (11), MH4 (1), MH5 (1) | 0.584 | 0.014 | 9.111 | |
| NC-SC | 1 | MH1 (2) | 0 | 0 | 0 | |
| TN-CQ | 3 | MH1 (13), MH3 (17), MH6 (2) | 0.567 | 0.014 | 9.085 | |
| DY-HB | 2 | MH1 (4), MH3 (6) | 0.513 | 0.014 | 9.231 | |
| XZ-HB | 1 | MH3 (30) | 0 | 0 | 0 | |
| ZY-HB | 1 | MH3 (32) | 0 | 0 | 0 | |
| ZX-HB | 1 | MH1 (4) | 0 | 0 | 0 | |
| SN-JS | 2 | MH3 (30), MH7 (1) | 0.065 | 0 | 0.065 | |
| XZ-JS | 1 | MH3 (28) | 0 | 0 | 0 | |
| SY-JS | 1 | MH3 (30) | 0 | 0 | 0 | |
| DH-JS | 1 | MH3 (3) | 0 | 0 | 0 | |
| FN-AH | 2 | MH3 (30), MH8 (1) | 0.065 | 0 | 0.065 | |
| BZ-AH | 1 | MH1 (1) | 0 | 0 | 0 | |
| CF-AH | 3 | MH1 (4), MH3 (5), MH9 (4) | 0.708 | 0.014 | 8.917 | |
| XX-AH | 2 | MH3 (2), MH9 (27) | 0.114 | 0.003 | 1.825 | |
| DC-HN | 2 | MH1 (4), MH3 (18) | 0.280 | 0.008 | 5.040 | |
| SZ-HN | 2 | MH1 (15), MH9 (2) | 0.221 | 0 | 0.441 | |
| TH-HN | 1 | MH3 (17) | 0 | 0 | 0 | |
| LH-HN | 3 | MH1 (4), MH3 (9), MH9 (18) | 0.581 | 0.011 | 7.277 | |
| GT-SD | 3 | MH1 (14), MH3 (2), MH9 (2) | 0.288 | 0.007 | 4.275 | |
| P. tectus | RX-SC | 1 | TH1 (31) | 0 | 0 | 0 |
| ST-SC | 1 | TH1 (1) | 0 | 0 | 0 | |
| NC-SC | 1 | TH1 (29) | 0 | 0 | 0 | |
| DY-HB | 2 | TH1 (14), TH5 (2) | 0.199 | 0.000 | 0.199 | |
| XZ-JS | 1 | TH1 (2) | 0 | 0 | 0 | |
| DH-JS | 1 | TH1 (1) | 0 | 0 | 0 | |
| BZ-AH | 2 | TH1 (18), TH2 (2) | 0.138 | 0.000 | 0.138 | |
| CF-AH | 4 | TH1 (9), TH2 (1), TH3 (2), TH4 (1) | 0.467 | 0.001 | 0.686 | |
| SZ-HN | 1 | TH1 (12) | 0 | 0 | 0 | |
| TH-HN | 1 | TH1 (13) | 0 | 0 | 0 | |
| RC-SX | 1 | TH1 (1) | 0 | 0 | 0 |
| Locations | MZ- SC | ST- SC | NC- SC | TN- CQ | DY- HB | XZ- HB | ZY- HB | ZX- HB | SN- JS | XZ- JS | SY- JS | DH-JS | FN- AH | BZ- AH | CF- AH | XX- AH | DC- HN | SZ- HN | TH- HN | LH- HN | GT- SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MZ-SC | inf | 0.84 | inf | inf | 0.30 | 0.29 | 0.75 | 0.30 | 0.31 | 0.30 | 0.91 | 0.30 | 9.01 | inf | 0.41 | 1.62 | 0.31 | 0.40 | 2.80 | 0.64 | |
| ST-SC | −0.01 | 3.32 | 7.33 | 83.08 | 0.18 | 0.17 | 0.40 | 0.17 | 0.18 | 0.18 | 0.45 | 0.17 | inf | inf | 0.75 | 0.70 | 0.51 | 0.23 | inf | 1.36 | |
| NC-SC | 0.23 | 0.07 | 0.55 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | inf | 10.62 | inf | inf | inf | inf | inf | inf | |
| TN-CQ | −0.02 | 0.03 | 0.31 | inf | 0.41 | 0.40 | 1.15 | 0.41 | 0.43 | 0.41 | 1.50 | 0.41 | 1.36 | 7.56 | 0.31 | 3.13 | 0.25 | 0.55 | 1.50 | 0.46 | |
| DY-HB | −0.06 | 0.00 | 0.31 | −0.07 | 0.21 | 0.20 | 1.11 | 0.21 | 0.22 | 0.21 | 1.67 | 0.21 | 2.00 | inf | 0.22 | 4.75 | 0.14 | 0.33 | 1.70 | 0.38 | |
| XZ-HB | 0.45 | 0.59 | 1.00 | 0.38 | 0.55 | inf | inf | inf | inf | inf | inf | inf | 0.00 | 0.10 | 0.02 | 1.17 | 0.00 | inf | 0.11 | 0.02 | |
| ZY-HB | 0.46 | 0.60 | 1.00 | 0.38 | 0.56 | 0.00 | inf | 249.8 | inf | inf | inf | 249.8 | 0.00 | 0.10 | 0.02 | 1.12 | 0.00 | inf | 0.11 | 0.02 | |
| ZX-HB | 0.25 | 0.39 | 1.00 | 0.18 | 0.18 | 0.00 | 0.00 | inf | inf | inf | inf | inf | 0.00 | 0.33 | 0.04 | inf | 0.00 | inf | 0.23 | 0.06 | |
| SN-JS | 0.46 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | −0.14 | inf | inf | inf | inf | 0.00 | 0.10 | 0.02 | 1.16 | 0.00 | inf | 0.11 | 0.02 | |
| XZ-JS | 0.44 | 0.58 | 1.00 | 0.37 | 0.53 | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | inf | 0.00 | 0.11 | 0.02 | 1.24 | 0.00 | inf | 0.11 | 0.03 | |
| SY-JS | 0.45 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | 0.00 | 0.10 | 0.02 | 1.17 | 0.00 | inf | 0.11 | 0.02 | |
| DH-JS | 0.22 | 0.36 | 1.00 | 0.14 | 0.13 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.00 | inf | 0.00 | 0.38 | 0.04 | inf | 0.00 | inf | 0.23 | 0.06 | |
| FN-AH | 0.46 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | −0.14 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.10 | 0.02 | 1.16 | 0.00 | inf | 0.11 | 0.02 | |
| BZ-AH | 0.03 | −0.23 | 0.00 | 0.16 | 0.11 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | inf | inf | 0.15 | inf | 0.00 | inf | inf | |
| CF-AH | −0.01 | −0.06 | 0.02 | 0.03 | 0.00 | 0.71 | 0.72 | 0.43 | 0.71 | 0.70 | 0.71 | 0.40 | 0.71 | −0.33 | 0.72 | 0.59 | 0.41 | 0.15 | inf | 1.59 | |
| XX-AH | 0.38 | 0.25 | −0.30 | 0.45 | 0.54 | 0.93 | 0.93 | 0.88 | 0.93 | 0.93 | 0.93 | 0.87 | 0.93 | −0.93 | 0.26 | 0.09 | 27.53 | 0.02 | 1.80 | inf | |
| DC-HN | 0.13 | 0.26 | 0.66 | 0.07 | 0.05 | 0.17 | 0.18 | −0.03 | 0.18 | 0.17 | 0.18 | −0.08 | 0.18 | 0.62 | 0.30 | 0.73 | 0.07 | 1.85 | 0.36 | 0.14 | |
| SZ-HN | 0.44 | 0.33 | 0.00 | 0.50 | 0.64 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 0.00 | 0.38 | 0.01 | 0.79 | 0.00 | 0.97 | 4.8 | |
| TH-HN | 0.38 | 0.52 | 1.00 | 0.31 | 0.43 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.00 | 0.00 | −0.02 | 1.00 | 0.62 | 0.91 | 0.12 | 1.00 | 0.14 | 0.03 | |
| LH-HN | 0.08 | 0.00 | −0.07 | 0.14 | 0.13 | 0.70 | 0.70 | 0.54 | 0.70 | 0.69 | 0.70 | 0.52 | 0.70 | −0.47 | −0.04 | 0.12 | 0.41 | 0.21 | 0.64 | 5.18 | |
| GT-SD | 0.28 | 0.16 | −0.27 | 0.35 | 0.40 | 0.91 | 0.91 | 0.81 | 0.91 | 0.91 | 0.91 | 0.80 | 0.91 | −0.88 | 0.14 | −0.04 | 0.65 | 0.05 | 0.88 | 0.05 |
| Locations | RX-SC | ST-SC | NC-SC | XZ-JS | BZ-AH | CF-AH | SZ-HN | TH-HN | RC-SX | DY-HB |
|---|---|---|---|---|---|---|---|---|---|---|
| RX-SC | inf | inf | inf | 2.63 | 0.58 | inf | inf | inf | 1.63 | |
| ST-SC | 0.00 | inf | inf | inf | inf | inf | inf | inf | inf | |
| NC-SC | 0.00 | 0.00 | inf | 2.83 | 0.62 | inf | inf | inf | 1.74 | |
| XZ-JS | 0.00 | 0.00 | 0.00 | inf | inf | inf | inf | inf | inf | |
| DH-JS | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | inf | inf | inf | |
| BZ-AH | 0.09 | −0.89 | 0.08 | −0.28 | 2.35 | 15.00 | 11.04 | inf | 3.76 | |
| CF-AH | 0.30 | −0.67 | 0.29 | −0.16 | 0.10 | 1.35 | 1.25 | inf | 1.3 | |
| SZ-HN | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.16 | inf | inf | 5.79 | |
| TH-HN | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.17 | 0.00 | inf | 4.92 | |
| RC-SX | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | −0.89 | −0.67 | 0.00 | inf | |
| DY-HB | 0.13 | −0.87 | 0.13 | −0.27 | 0.06 | 0.16 | 0.04 | 0.05 | -0.87 |
| Locations | Southwestern China (SW) | Middle and Lower Yangtze Valleys (YV) | Yellow and Huai Valleys (YH) |
|---|---|---|---|
| Southwestern China (SW) | 0.43819 | 1.08303 | |
| Middle and Lower Yangtze Valleys (YV) | 0.36327 | 3.81602 | |
| Yellow and Huai Valleys (YH) | 0.18754 | 0.06149 |
| Locations | Southwestern China (SW) | Middle and Lower Yangtze Valleys (YV) | Yellow and Huai Valleys (YH) |
|---|---|---|---|
| Southwestern China (SW) | 4.23242 | 0.84639 | |
| Middle and Lower Yangtze Valleys (YV) | 0.05577 | 4.07018 | |
| Yellow and Huai Valleys (YH) | 0.22802 | 0.05787 |
| Species | Source of Variation | df | Sum of Squares | % of Variation | Fixation Indices | p-Value |
|---|---|---|---|---|---|---|
| P. major | Among regions | 2 | 152.02 | 2.05 | FCT = 0.02 | p = 0.332 |
| Among locations within regions | 18 | 859.49 | 55.74 | FSC = 0.57 | p < 0.001 | |
| Within locations | 417 | 733.64 | 42.20 | FST = 0.58 | p < 0.001 | |
| P. tectus | Among regions | 2 | 0.53 | 2.35 | FCT = 0.02 | p = 0.206 |
| Among locations within regions | 8 | 1.04 | 9.21 | FSC = 0.09 | p = 0.239 | |
| Within locations | 128 | 8.17 | 88.43 | FST = 0.12 | p = 0.070 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, W.; Qin, Y.; Wang, M.; Li, Y.; Liu, H. Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China. Insects 2023, 14, 377. https://doi.org/10.3390/insects14040377
Wang X, Wang W, Qin Y, Wang M, Li Y, Liu H. Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China. Insects. 2023; 14(4):377. https://doi.org/10.3390/insects14040377
Chicago/Turabian StyleWang, Xian, Wenjie Wang, Yang Qin, Mian Wang, Yaying Li, and Huai Liu. 2023. "Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China" Insects 14, no. 4: 377. https://doi.org/10.3390/insects14040377
APA StyleWang, X., Wang, W., Qin, Y., Wang, M., Li, Y., & Liu, H. (2023). Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China. Insects, 14(4), 377. https://doi.org/10.3390/insects14040377