
Automatic Detection of Moths (Lepidoptera) with a Funnel Trap Prototype

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the New Probe (ZooLog VARL)
2.1.1. Modified VARL Trap
2.1.2. Sensor-Ring
2.1.3. Data Communication System
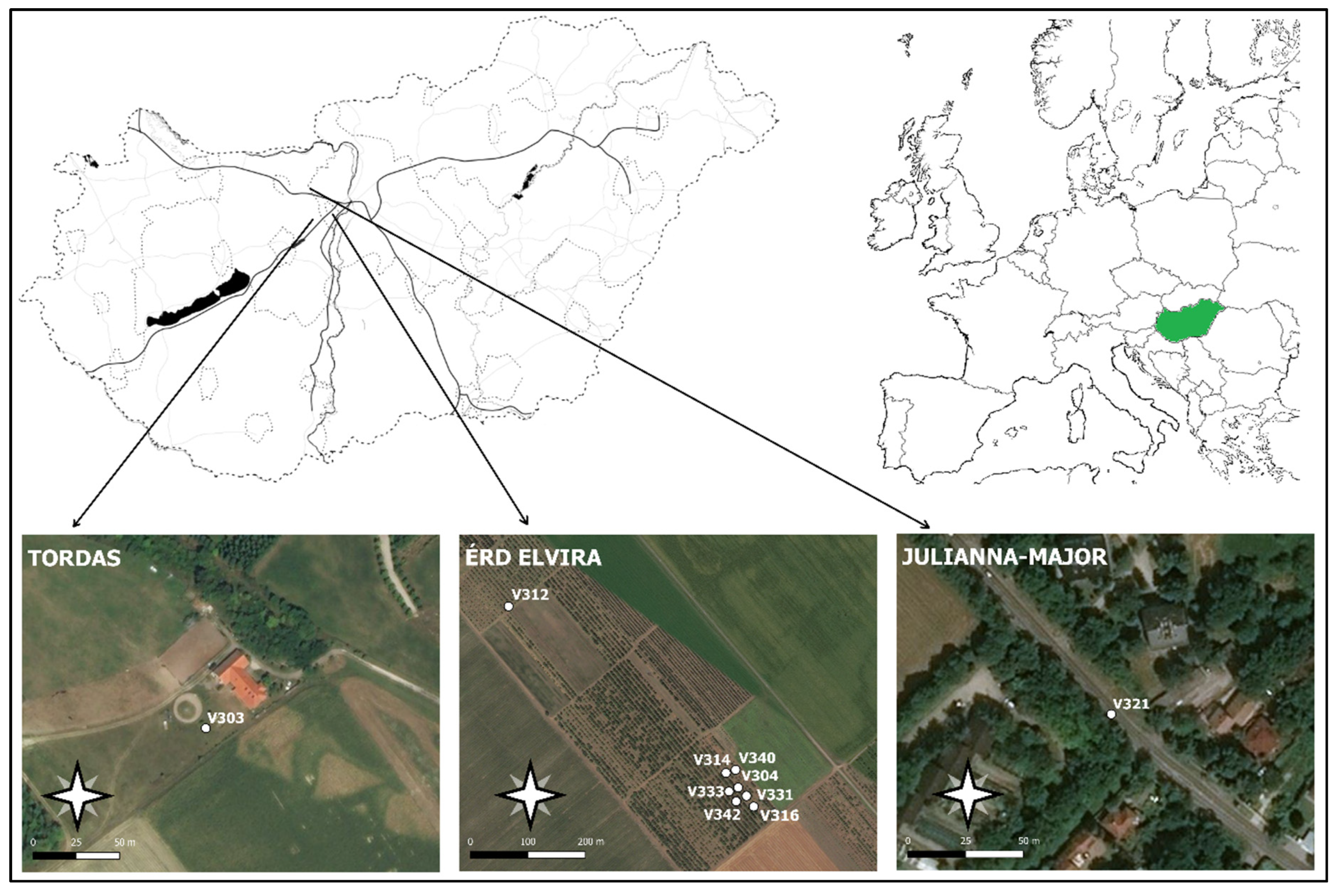
2.2. Field Tests
2.3. Data Analysis
2.3.1. Data Filtering
2.3.2. Statistical Analysis
3. Results
3.1. Data Filtering Using ANN (Artificial Neural Network) Approach
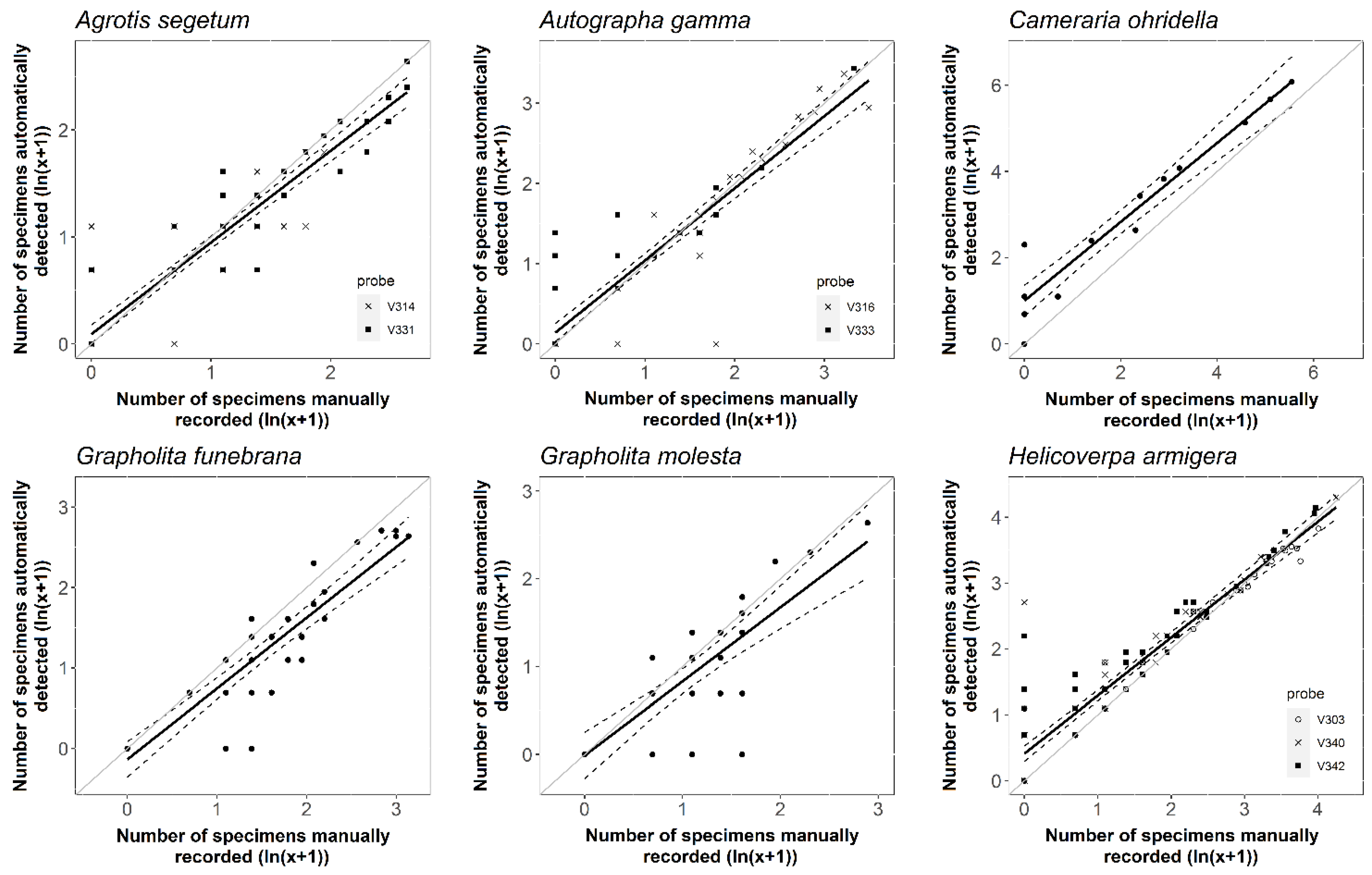
3.2. Detection Accuracy and Performance of the New Probe
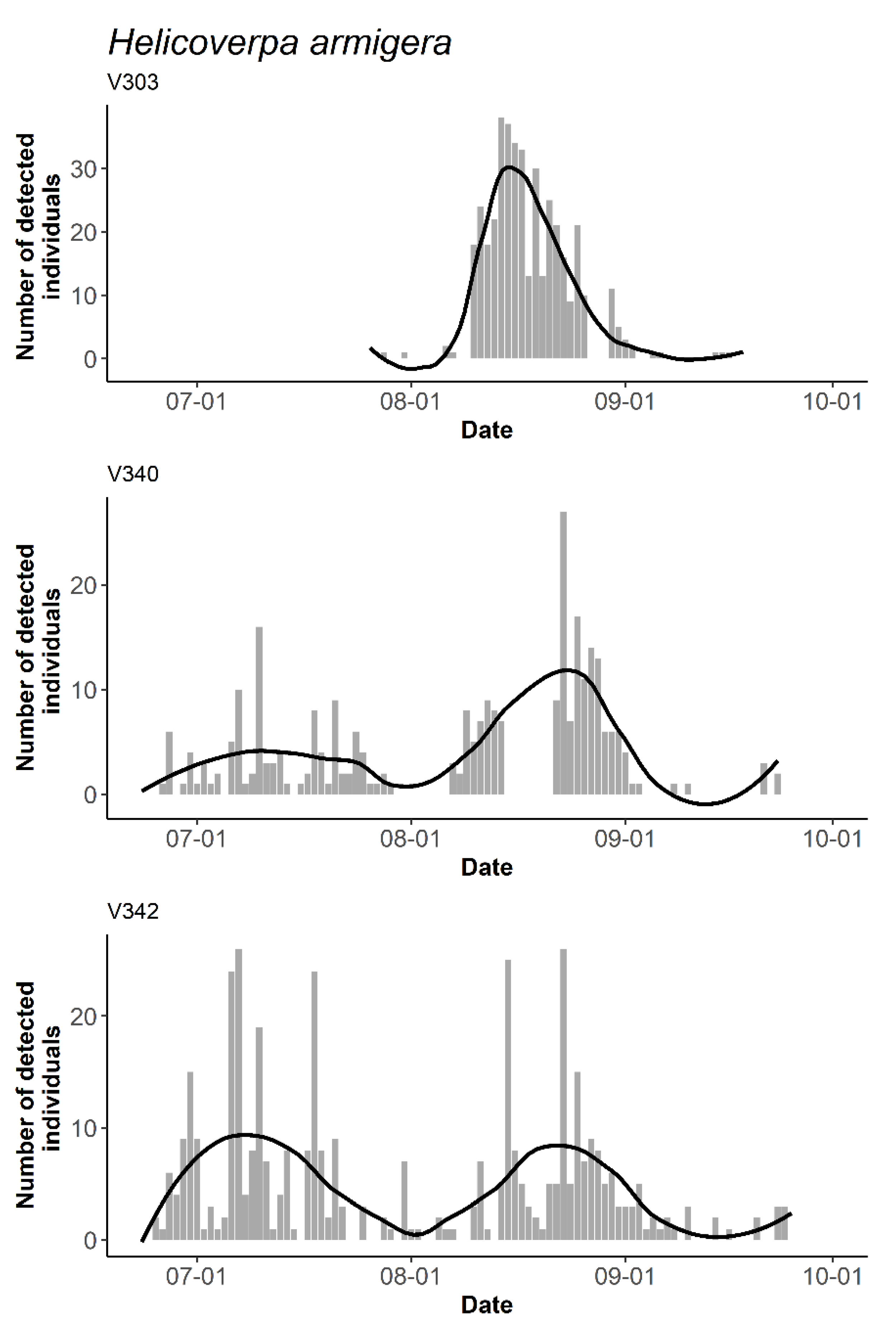
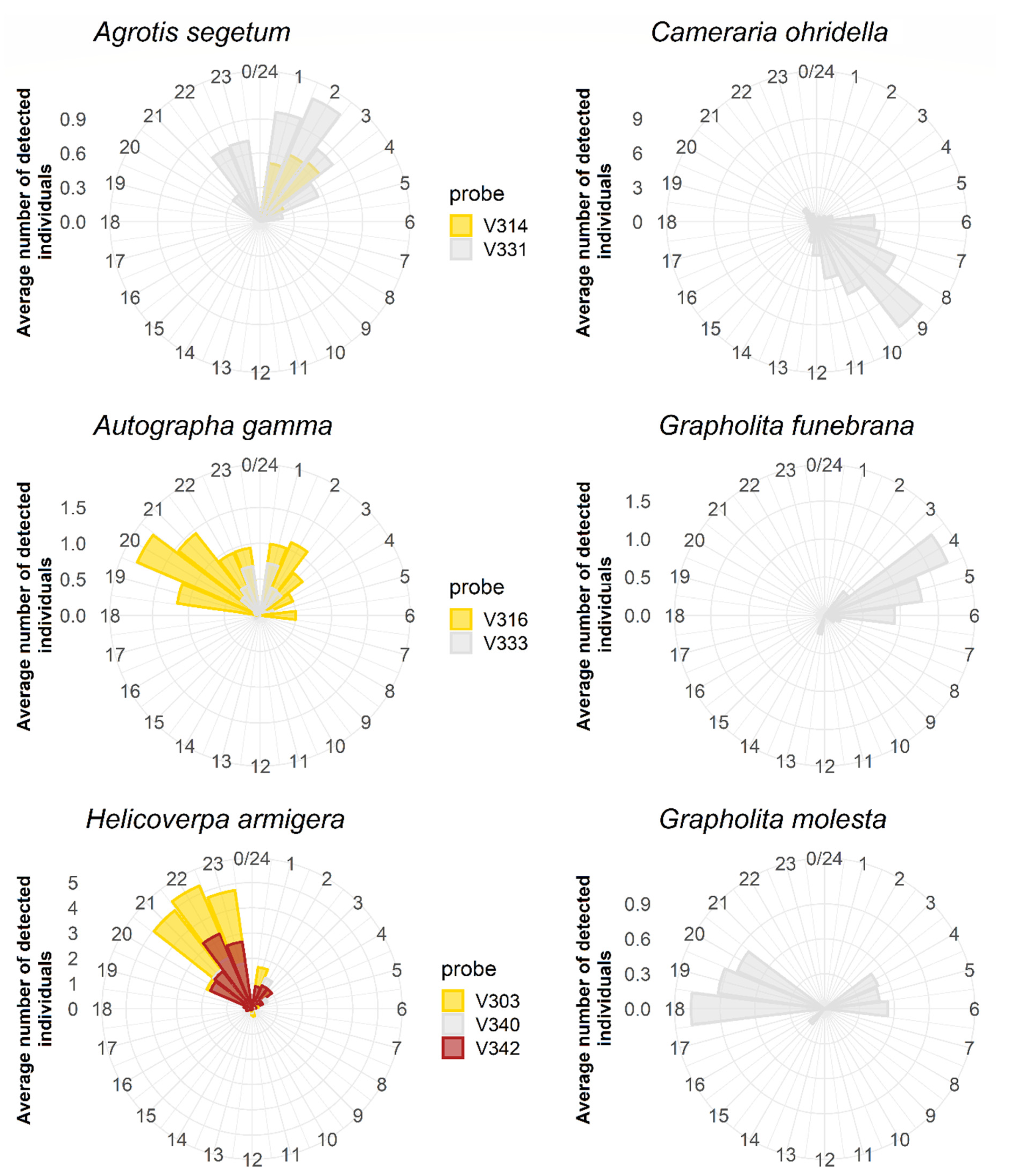
3.3. The Temporal and Daily Activity Patterns of Target Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kogan, M.; Hilton, R.J. Conceptual framework for integrated pest management (IPM) of tree-fruit pests. In Biorational Tree-Fruit Pest Management; CABI: Wallingford, UK, 2009; pp. 1–31. [Google Scholar]
- Preti, M.; Verheggen, F.; Angeli, S. Insect pest monitoring with camera-equipped traps: Strengths and limitations. J. Pest Sci. 2021, 94, 203–217. [Google Scholar] [CrossRef]
- Cardim Ferreira Lima, M.; Damascena de Almeida Leandro, M.E.; Valero, C.; Pereira Coronel, L.C.; Gonçalves Bazzo, C.O. Automatic Detection and Monitoring of Insect Pests—A Review. Agriculture 2020, 10, 161. [Google Scholar] [CrossRef]
- Čirjak, D.; Miklečić, I.; Lemić, D.; Kos, T.; Pajač Živković, I. Automatic pest monitoring systems in apple production under changing climatic conditions. Horticulturae 2022, 8, 520. [Google Scholar] [CrossRef]
- Liu, H.; Lee, S.-H.; Chahl, J.S. A review of recent sensing technologies to detect invertebrates on crops. Precis. Agric. 2017, 18, 635–666. [Google Scholar] [CrossRef]
- Vasconcelos, D.; Nunes, N.; Ribeiro, M.; Prandi, C.; Rogers, A. LOCOMOBIS: A low-cost acoustic-based sensing system to monitor and classify mosquitoes. In Proceedings of the 2019 16th IEEE Annual Consumer Communications & Networking Conference (CCNC), Las Vegas, NV, USA, 11–14 January 2019; pp. 1–6. [Google Scholar]
- Mankin, R.; Machan, R.; Jones, R. Field testing of a prototype acoustic device for detection of Mediterranean fruit flies flying into a trap. In Proceedings of the 7th International Symposium Fruit Flies of Economic Importance, Salvador, Brazil, 10–15 September 2006; pp. 165–169. [Google Scholar]
- Potamitis, I.; Rigakis, I.; Vidakis, N.; Petousis, M.; Weber, M. Affordable bimodal optical sensors to spread the use of automated insect monitoring. J. Sens. 2018, 2018, 3949415. [Google Scholar] [CrossRef] [Green Version]
- Frewin, A.; Lopez, B.; Cox, A.; Hoffman, E.; Hazell, J. Comparison of two traps for monitoring California red scale (Hemiptera: Diaspididae). Fla. Entomol. 2019, 102, 586–591. [Google Scholar] [CrossRef]
- Rustia, D.J.A.; Lin, C.E.; Chung, J.-Y.; Zhuang, Y.-J.; Hsu, J.-C.; Lin, T.-T. Application of an image and environmental sensor network for automated greenhouse insect pest monitoring. J. Asia-Pac. Entomol. 2020, 23, 17–28. [Google Scholar] [CrossRef]
- Zhong, Y.; Gao, J.; Lei, Q.; Zhou, Y. A vision-based counting and recognition system for flying insects in intelligent agriculture. Sensors 2018, 18, 1489. [Google Scholar] [CrossRef] [Green Version]
- Roosjen, P.P.; Kellenberger, B.; Kooistra, L.; Green, D.R.; Fahrentrapp, J. Deep learning for automated detection of Drosophila suzukii: Potential for UAV-based monitoring. Pest Manag. Sci. 2020, 76, 2994–3002. [Google Scholar] [CrossRef]
- Jiang, J.A.; Tseng, C.L.; Lu, F.M.; Yang, E.C.; Wu, Z.S.; Chen, C.P.; Lin, S.H.; Lin, K.C.; Liao, C.S. A GSM-based remote wireless automatic monitoring system for field information: A case study for ecological monitoring of the oriental fruit fly, Bactrocera dorsalis (Hendel). Comput. Electron. Agric. 2008, 62, 243–259. [Google Scholar] [CrossRef]
- Holguin, G.A.; Lehman, B.L.; Hull, L.A.; Jones, V.P.; Park, J. Electronic traps for automated monitoring of insect populations. IFAC Proc. Vol. 2010, 43, 49–54. [Google Scholar] [CrossRef]
- Rigakis, I.; Potamitis, I.; Tatlas, N.-A.; Livadaras, I.; Ntalampiras, S. A multispectral backscattered light recorder of insects’ wingbeats. Electronics 2019, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Jung, S.; Kim, Y.; Lee, Y. Real-time monitoring of oriental fruit moth, Grapholita molesta, populations using a remote sensing pheromone trap in apple orchards. J. Asia-Pac. Entomol. 2011, 14, 259–262. [Google Scholar] [CrossRef]
- Dombos, M.; Kosztolányi, A.; Szlávecz, K.; Gedeon, C.; Flórián, N.; Groó, Z.; Dudás, P.; Bánszegi, O. EDAPHOLOG monitoring system: Automatic, real-time detection of soil microarthropods. Methods Ecol. Evol. 2017, 8, 313–321. [Google Scholar] [CrossRef]
- Gedeon, C.I.; Flórián, N.; Liszli, P.; Hambek-Oláh, B.; Bánszegi, O.; Schellenberger, J.; Dombos, M. An Opto-electronic sensor for detecting soil microarthropods and estimating their size in field conditions. Sensors 2017, 17, 1757. [Google Scholar] [CrossRef] [Green Version]
- Balla, E.; Flórián, N.; Gergócs, V.; Gránicz, L.; Tóth, F.; Németh, T.; Dombos, M. An Opto-electronic Sensor-ring to Detect Arthropods of Significantly Different Body Sizes. Sensors 2020, 20, 982. [Google Scholar] [CrossRef] [Green Version]
- Flórián, N.; Gránicz, L.; Gergócs, V.; Tóth, F.; Dombos, M. Detecting Soil Microarthropods with a Camera-Supported Trap. Insects 2020, 11, 244. [Google Scholar] [CrossRef]
- Tóth, Z.; Tóth, M.; Jósvai, J.K.; Tóth, F.; Flórián, N.; Gergócs, V.; Dombos, M. Automatic Field Detection of Western Corn Rootworm (Diabrotica virgifera virgifera; Coleoptera: Chrysomelidae) with a New Probe. Insects 2020, 11, 486. [Google Scholar] [CrossRef]
- Tóth, M.; Imrei, Z.; Szôcs, G. Non-sticky, non-saturable, high capacity new pheromone traps for Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) and Helicoverpa (Heliothis) armigera (Lepidoptera: Noctuidae). Integr. Term. Kert. Szántóf. Kult 2000, 21, 44–49. [Google Scholar]
- Tóth, M.; Imrei, Z.; Szöcs, G.; Ujváry, I.; Kárpáti, Z.; Jermi, T. Development of new and more efficient funnel trap designs for pest moths of larger size (Lepidoptera, Noctuidae, Geometridae) and yellowjackets (Vespa spp., Hymenoptera, Vespidae). In Proceedings of the Abstract Conference 47th Plant Protection Days, Budapest, Hungary, 27–28 February 2001; p. 68. (In Hungarian). [Google Scholar]
- Jiang, J.A.; Lin, T.S.; Yang, E.C.; Tseng, C.L.; Chen, C.P.; Yen, C.W.; Zheng, X.Y.; Liu, C.Y.; Liu, R.H.; Chen, Y.F. Application of a web-based remote agro-ecological monitoring system for observing spatial distribution and dynamics of Bactrocera dorsalis in fruit orchards. Precis. Agric. 2013, 14, 323–342. [Google Scholar] [CrossRef]
- Shieh, J.C.; Wang, J.Y.; Lin, T.S.; Lin, C.H.; Yang, E.C.; Tsai, Y.J.; Tsai, H.T.; Chiou, M.T.; Lu, F.M.; Jiang, J.A. A GSM-based field monitoring system for Spodoptera litura (Fabricius). Eng. Agric. Environ. Food 2011, 4, 77–82. [Google Scholar] [CrossRef]
- Löfstedt, C.; Van Der Pers, J.N.; Lofqvist, J.; Lanne, B.S.; Appelgren, M.; Bergström, G.; Thelin, B. Sex pheromone components of the turnip moth, Agrotis segetum. J. Chem. Ecol. 1982, 8, 1305–1321. [Google Scholar] [CrossRef]
- Mazor, M.; Dunkelblum, E. Circadian rhythms of sexual behavior and pheromone titers of two closely related moth species Autographa gamma and Cornutiplusia circumflexa. J. Chem. Ecol. 2005, 31, 2153–2168. [Google Scholar] [CrossRef] [PubMed]
- Francke, W.; Franke, S.; Bergmann, J.; Tolasch, T.; Subchev, M.; Mircheva, A.; Toshova, T.; Svatoš, A.; Kalinová, B.; Kárpáti, Z. Female sex pheromone of Cameraria ohridella Desch. and Dim.(Lepidoptera: Gracillariidae): Structure confirmation, synthesis and biological activity of (8E, 10Z)-8, 10-tetradecadienal and some analogues. Z. Für Nat. C 2002, 57, 739–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jermy, T.; Balázs, K. A Növényvédelmi Állattan Kézikönyve; Akadémiai Kiadó: Budapest, Hungary, 1993; Volume 4/A, pp. 373–380. [Google Scholar]
- Han, K.S.; Jung, J.K.; Choi, K.H.; Lee, S.W.; Boo, K.S. Sex pheromone composition and male trapping of the oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae) in Korea. J. Asia-Pac. Entomol. 2001, 4, 31–35. [Google Scholar] [CrossRef]
- Kou, R.; Chow, Y.-S. Calling behavior of the cotton bollworm, Heliothis armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1987, 80, 490–493. [Google Scholar] [CrossRef]
- Tóth, M. The pheromones and their practical application (in Hungarian). In Integrated Pest Management of Pests; Jenser, G., Ed.; Mezőgazda Kiadó: Budapest, Hungary, 2003; pp. 21–50. [Google Scholar]
- Abadi, M.; Agarwal, A.; Barham, P.; Brevdo, E.; Chen, Z.; Citro, C.; Corrado, G.S.; Davis, A.; Dean, J.; Devin, M. Tensorflow: Large-scale machine learning on heterogeneous distributed systems. arXiv 2016, arXiv:1603.04467. [Google Scholar]
- Github. The TensorBoard Repository on GitHub. 2016. Available online: http://github.com/tensorflow/tensorboard (accessed on 15 June 2017).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. J. Math. Methods Biosci. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Getting Started with ggplot2. In ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; pp. 11–31. [Google Scholar]
- Grolemund, G.; Wickham, H. Dates and times made easy with lubridate. J. Stat. Softw. 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Preti, M.; Favaro, R.; Knight, A.L.; Angeli, S. Remote monitoring of Cydia pomonella adults among an assemblage of nontargets in sex pheromone-kairomone-baited smart traps. Pest Manag. Sci. 2021, 77, 4084–4090. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Szarukán, I.; Szalárdi, T.; Szanyi, S.; Jósvai, J.K.; Tóth, M. Addition of 4-oxoisophorone improves performance of bisexual lure for Autographa gamma (L.) (Lepidoptera: Noctuidae). J. Appl. Entomol. 2022, 146, 328–334. [Google Scholar] [CrossRef]
- Dunkelblum, E.; Bestmann, H.J.; Knauf, W.; Vostrowsky, O. Sex attractant blends for field trapping of Agrotis segetum males (Lepidoptera: Noctuidae) in Israel and Germany. Z. Für Nat. C 1985, 40, 272–277. [Google Scholar] [CrossRef]
- Dömötör, I.; Kiss, J.; Szőcs, G. First results on synchrony between seasonal pattern of pheromone trap captures of cotton bollworm, Helicoverpa armigera and appearance of freshly emerged larvae on developing cobs of corn hybrids. J. Pest Sci. 2007, 80, 183–189. [Google Scholar] [CrossRef]
- Pérez Aparicio, A.; Llorens Calveras, J.; Rosell Polo, J.R.; Martí, J.; Gemeno Marín, C. A cheap electronic sensor automated trap for monitoring the flight activity period of moths. Eur. J. Entomol. 2021, 118, 315–321. [Google Scholar] [CrossRef]
- Sziráki, G. Examinations on tortricid moths trapped by synthetic attractants (Lepidoptera). Folia Entomol. Hung. 1978, 31, 259–264. [Google Scholar]
- Tòth, M.; Sziràki, G.; Szöcs, G.; Sàringer, E. A pheromone inhibitor for male Grapholitha funebrana Tr. and its use for increasing the specificity of the lure for G. molesta Busck (Lepidoptera: Tortricidae). Agric. Ecosyst. Environ. 1991, 35, 65–72. [Google Scholar] [CrossRef]
- Sziraki, G. Specificity of sexual attractant traps for signalization of oriental fruit moth (Grapholita molesta Busck). Acta Phytopathol. Acad. Sci. Hung. 1978, 13, 205–212. [Google Scholar]
- Gentry, C.; Beroza, M.; Blythe, J. Pecan bud moth: Captures in Georgia in traps baited with the pheromone of the oriental fruit moth. Environ. Entomol. 1975, 4, 227–228. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex pheromone aerosol devices for mating disruption: Challenges for a brighter future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Ünlü, L.; Akdemir, B.; Ögür, E.; Şahin, İ. Remote monitoring of European grapevine moth, Lobesia botrana (Lepidoptera: Tortricidae) population using camera-based pheromone traps in vineyards. Turk. J. Agric.-Food Sci. Technol. 2019, 7, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Ascolese, R.; Gargiulo, S.; Pace, R.; Nappa, P.; Griffo, R.; Nugnes, F.; Bernardo, U. E-traps: A valuable monitoring tool to be improved. EPPO Bull. 2022, 52, 175–184. [Google Scholar] [CrossRef]
- Okuyama, T.; Yang, E.-C.; Chen, C.-P.; Lin, T.-S.; Chuang, C.-L.; Jiang, J.-A. Using automated monitoring systems to uncover pest population dynamics in agricultural fields. Agric. Syst. 2011, 104, 666–670. [Google Scholar] [CrossRef]
- Chuang, C.-L.; Jiang, J.-A. ICT-based Remote Agro-Ecological Monitoring System—A Case Study inTaiwan. J. Commun. Navig. Sens. Serv. (CONASENSE) 2014, 1, 67–92. [Google Scholar] [CrossRef] [Green Version]
- Goldshtein, E.; Cohen, Y.; Hetzroni, A.; Gazit, Y.; Timar, D.; Rosenfeld, L.; Grinshpon, Y.; Hoffman, A.; Mizrach, A. Development of an automatic monitoring trap for Mediterranean fruit fly (Ceratitis capitata) to optimize control applications frequency. Comput. Electron. Agric. 2017, 139, 115–125. [Google Scholar] [CrossRef]
- Brydegaard, M.; Gebru, A.; Kirkeby, C.; Åkesson, S.; Smith, H. Daily evolution of the insect biomass spectrum in an agricultural landscape accessed with lidar. In Proceedings of the EPJ Web of Conferences, New York, NY, USA, 5–10 July 2015; EDP Sciences: Les Ulis, France, 2016; p. 22004. [Google Scholar]
- Song, Z.; Zhang, B.; Feng, H.; Zhu, S.; Hu, L.; Brydegaard, M.; Li, Y.; Jansson, S.; Malmqvist, E.; Svanberg, K. Application of lidar remote sensing of insects in agricultural entomology on the Chinese scene. J. Appl. Entomol. 2020, 144, 161–169. [Google Scholar] [CrossRef]
- Jansson, S.; Brydegaard, M. Passive kHz lidar for the quantification of insect activity and dispersal. Anim. Biotelemetry 2018, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Kirkeby, C.; Wellenreuther, M.; Brydegaard, M. Observations of movement dynamics of flying insects using high resolution lidar. Sci. Rep. 2016, 6, 29083. [Google Scholar] [CrossRef] [Green Version]
- Tobin, P.C.; Klein, K.T.; Leonard, D.S. Gypsy moth (Lepidoptera: Lymantriidae) flight behavior and phenology based on field-deployed automated pheromone-baited traps. Environ. Entomol. 2009, 38, 1555–1562. [Google Scholar] [CrossRef] [Green Version]
- Hirafuji, M.; Yoichi, H.; Watanabe, T.; Asai, M.; Hu, H.; Tanaka, K.; Fukatsu, T.; Kiura, T.; Ninomiya, S.; Nagatsuka, T. Real-time insect monitoring system by using Field Server. In Proceedings of the World Conference on Agricultural Information and IT, IAALD AFITA WCCA, Tokyo, Japan, 24–27 August 2008; pp. 277–282. [Google Scholar]
- Solis-Sánchez, L.O.; Castañeda-Miranda, R.; García-Escalante, J.J.; Torres-Pacheco, I.; Guevara-González, R.G.; Castañeda-Miranda, C.L.; Alaniz-Lumbreras, P.D. Scale invariant feature approach for insect monitoring. Comput. Electron. Agric. 2011, 75, 92–99. [Google Scholar] [CrossRef]
- Doitsidis, L.; Fouskitakis, G.N.; Varikou, K.N.; Rigakis, I.I.; Chatzichristofis, S.A.; Papafilippaki, A.K.; Birouraki, A.E. Remote monitoring of the Bactrocera oleae (Gmelin) (Diptera: Tephritidae) population using an automated McPhail trap. Comput. Electron. Agric. 2017, 137, 69–78. [Google Scholar] [CrossRef]
- Guarnieri, A.; Maini, S.; Molari, G.; Rondelli, V. Automatic trap for moth detection in integrated pest management. Bull. Insectol. 2011, 64, 247–251. [Google Scholar]
- Liu, B.; Hu, Z.; Zhao, Y.; Bai, Y.; Wang, Y. Recognition of Pyralidae Insects Using Intelligent Monitoring Autonomous Robot Vehicle in Natural Farm Scene. arXiv 2019, arXiv:1903.10827. [Google Scholar]
- Ding, W.; Taylor, G. Automatic moth detection from trap images for pest management. Comput. Electron. Agric. 2016, 123, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Sipos, K.; Madár, S.; Markó, M.; Pénzes, B. The possibility of automation of sex pheromone trapping: Tested on Resseliella theobaldi (Barnes) (Dip). Afr. J. Agric. Res. 2012, 7, 1410–1413. [Google Scholar]
- Geissmann, Q.; Abram, P.K.; Wu, D.; Haney, C.H.; Carrillo, J. Sticky Pi is a high-frequency smart trap that enables the study of insect circadian activity under natural conditions. PLoS Biol. 2022, 20, e3001689. [Google Scholar] [CrossRef]
- Preti, M.; Moretti, C.; Scarton, G.; Giannotta, G.; Angeli, S. Developing a smart trap prototype equipped with camera for tortricid pests remote monitoring. Bull. Insectology 2021, 74, 147–160. [Google Scholar]
- Schrader, M.J.; Smytheman, P.; Beers, E.H.; Khot, L.R. An open-source low-cost imaging system plug-in for pheromone traps aiding remote insect pest population monitoring in fruit crops. Machines 2022, 10, 52. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Host and Symptoms | Flight Period | Daily Rhythm | Wingspan/Body Length (mm) | Test Site | Test Period | Number of Probes |
|---|---|---|---|---|---|---|---|
| Agrotis segetum Den. and Schiff., 1775 (turnip moth) | polyphagous, larvae feeding on the part of the host plant at the soil level or above 1–2 cm | 2 generations/year 1—beginning of May to the middle of June 2—beginning of July to September | 4–5 h to the scotophase [27] | 30–40/15–20 | Érd-Elvira major orchard | 2018.06.06.–2018.10.15. | 2 |
| Autographa gamma L., 1785 (silver Y) | polyphagous, larvae feeding on the leaves | 2 generations/year 1—beginning of May to the middle of June 2—middle of June to the beginning of October | during the scotophase [28] | 35–40/15–20 | Érd-Elvira major orchard | 2018.06.22.–2018.10.15. | 2 |
| Cameraria ohridella Deschka and Dimič, 1986 (horse-chestnut leafminer) | horse-chestnut (Aesculus hippocastanum), larvae bore mines into the leaves | 3 generations/year 1—end of April to the beginning of May 2—in June 3—middle of July to the beginning of September | beginning of photophase, lasts for 4–5 h (7 p.m.–2 p.m. in July in Hungary) [29] | 6–8/5 | Julianna-major, row of horse chestnut trees | 2018.06.25.–2018.10.15. | 1 |
| Grapholita funebrana Treitschke, 1985 (plum moth) | Prunus spp. (plum, apricot) Larvae bore inside the fruit feed on the flesh around the seeds | 3 generations/year 1—middle of April to the middle of May 2—end of May to the end of June 3—beginning of July to the beginning of August | end of the scotophase to the beginning of photophase [30] | 9–13/6–9 | Érd- Elvira major orchard | 2018.06.21.–2018.10.15. | 1 |
| Grapholita molesta Busck, 1916 (oriental fruit moth) | peach, almond, apricot, medlar, pear, apple spring–larvae bore into fresh shoots summer–larvae bore into ripening fruits | 3 generations/year 1—middle of April to the middle of May 2—end of May to the end of June 3—beginning of July to the beginning of August | during the scotophase [31] | 9–13/5–7 | Érd-Elvira major orchard | 2018.06.21.–2018.10.15. | 1 |
| Helicoverpa armigera Hübner, 1808 (cotton bollworm) | polyphagous, larvae feed on the generative parts of the plant | 3 generations/year 1—end of May to the beginning of June 2—middle of July to the middle of August 3—end of August to September | during the scotophase [32] | 30–40/12–20 | Érd-Elvira major orchard | 2018.06.22.–2018.10.15. | 2 |
| Tordas orchard | 2018.07.25.–2018.10.15. | 1 |
| Target Species | TP | TN | FP | FN | Filtering Accuracy | Precision | Recall | F1 Score |
|---|---|---|---|---|---|---|---|---|
| Agrotis segetum | 130 | 1132 | 63 | 28 | 0.93 | 0.67 | 0.82 | 0.74 |
| Autographa gamma | 201 | 1113 | 74 | 22 | 0.93 | 0.73 | 0.90 | 0.81 |
| Cameraria ohridella | 597 | 242 | 487 | 64 | 0.60 | 0.55 | 0.90 | 0.68 |
| Grapholita funebrana | 72 | 158 | 65 | 53 | 0.66 | 0.53 | 0.58 | 0.55 |
| Grapholita molesta | 78 | 205 | 6 | 20 | 0.92 | 0.93 | 0.80 | 0.86 |
| Helicoverpa armigera | 868 | 1055 | 246 | 23 | 0.88 | 0.78 | 0.97 | 0.87 |
| Target Species | Number of Measurement Periods | Average Accuracy | R2 | Slope | Estimation | ||
|---|---|---|---|---|---|---|---|
| y | p | intercept | p | ||||
| Agrotis segetum | 94 | 85.64% | 0.853 | 0.86 ± 0.04 | <0.001 | 0.09 ± 0.04 | 0.072 |
| Autographa gamma | 80 | 89.94% | 0.848 | 0.90 ± 0.04 | <0.001 | 0.14 ± 0.06 | 0.030 |
| Cameraria ohridella | 16 | 91.55% | 0.932 | 0.92 ± 0.06 | <0.001 | 1.00 ± 0.17 | <0.001 |
| Grapholita funebrana | 38 | 88.02% | 0.837 | 0.88 ± 0.06 | <0.001 | −0.14 ± 0.11 | 0.449 |
| Grapholita molesta | 40 | 84.29% | 0.614 | 0.84 ± 0.11 | <0.001 | −0.01 ± 0.13 | 1 |
| Helicoverpa armigera | 111 | 88.05% | 0.887 | 0.88 ± 0.03 | <0.001 | 0.41 ± 0.06 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flórián, N.; Jósvai, J.K.; Tóth, Z.; Gergócs, V.; Sipőcz, L.; Tóth, M.; Dombos, M. Automatic Detection of Moths (Lepidoptera) with a Funnel Trap Prototype. Insects 2023, 14, 381. https://doi.org/10.3390/insects14040381
Flórián N, Jósvai JK, Tóth Z, Gergócs V, Sipőcz L, Tóth M, Dombos M. Automatic Detection of Moths (Lepidoptera) with a Funnel Trap Prototype. Insects. 2023; 14(4):381. https://doi.org/10.3390/insects14040381
Chicago/Turabian StyleFlórián, Norbert, Júlia Katalin Jósvai, Zsolt Tóth, Veronika Gergócs, László Sipőcz, Miklós Tóth, and Miklós Dombos. 2023. "Automatic Detection of Moths (Lepidoptera) with a Funnel Trap Prototype" Insects 14, no. 4: 381. https://doi.org/10.3390/insects14040381
APA StyleFlórián, N., Jósvai, J. K., Tóth, Z., Gergócs, V., Sipőcz, L., Tóth, M., & Dombos, M. (2023). Automatic Detection of Moths (Lepidoptera) with a Funnel Trap Prototype. Insects, 14(4), 381. https://doi.org/10.3390/insects14040381