
Evaluating the Effect of Irradiation on the Densities of Two RNA Viruses in Glossina morsitans morsitans



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tsetse Flies
2.2. Irradiation Conditions
2.3. The Irradiation Process and Collection of Samples
2.4. RNA Extraction and DNase Treatment
2.5. cDNA Synthesis
2.6. Relative Quantitative PCR
2.7. Statistical Analysis
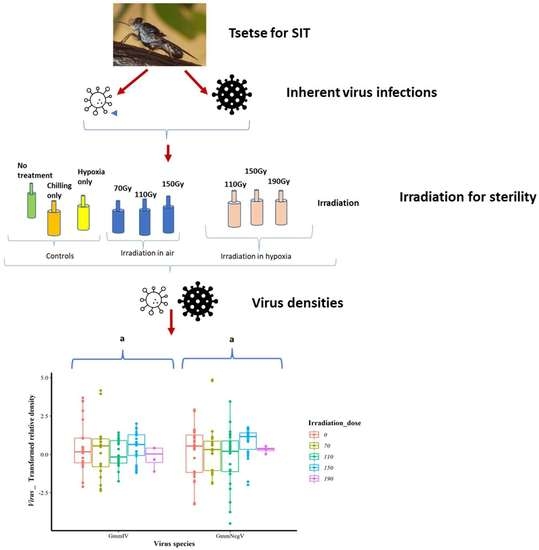
3. Results
3.1. Statistical Distribution of the Data
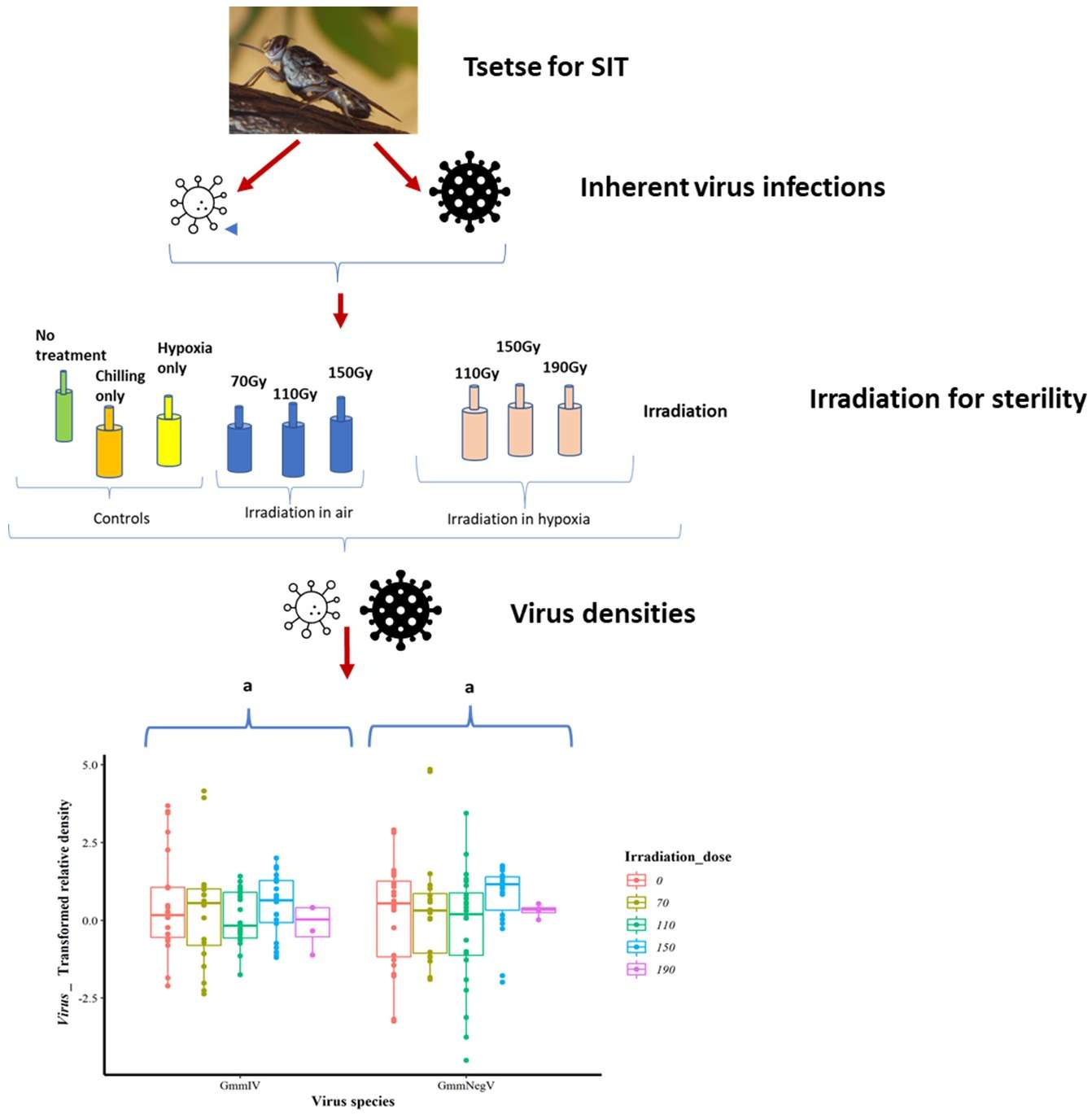
3.2. Impact of Irradiation Treatment on GmmIV Densities
3.3. Impact of Irradiation Conditions and Host Stage on GmmIV Densities
3.4. Impact of Irradiation Treatment Doses on GmmNegeV Densities
3.5. Impact of Irradiation Conditions and Host Life Stages on GmmNegeV Densities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rogers, D.; Hendrickx, G.; Slingenbergh, J. Tsetse flies and their control. Rev. Sci. Tech. Int. Off. Epizoot. 1994, 13, 1075–1124. [Google Scholar] [CrossRef]
- Steverding, D. The history of African trypanosomiasis. Parasites Vectors 2008, 1, 3–8. [Google Scholar] [CrossRef]
- Shaw, A.; Torr, S.; Waiswa, C.; Cecchi, G.; Wint, G.; Mattioli, R.; Robinson, T. Estimating the costs of tsetse control options: An example for Uganda. Prev. Veter-Med. 2013, 110, 290–303. [Google Scholar] [CrossRef]
- Simarro, P.P.; Cecchi, G.; Paone, M.; Franco, J.R.; Diarra, A.; Ruiz, J.A.; Fèvre, E.M.; Courtin, F.; Mattioli, R.C.; Jannin, J.G. The Atlas of human African trypanosomiasis: A contribution to global mapping of neglected tropical diseases. Int. J. Health Geogr. 2010, 9, 57. [Google Scholar] [CrossRef]
- Cecchi, G.; Mattioli, R.C.; Slingenbergh, J.; De La Rocque, S. Land cover and tsetse fly distributions in sub-Saharan Africa. Med. Veter-Èntomol. 2008, 22, 364–373. [Google Scholar] [CrossRef]
- Rogers, D.J.; Randolph, S.E. Distribution and abundance of tsetse flies (Glossina spp.). J. Anim. Ecol. 1986, 55, 1007–1025. [Google Scholar] [CrossRef]
- Shereni, W.; Anderson, N.E.; Nyakupinda, L.; Cecchi, G. Spatial distribution and trypanosome infection of tsetse flies in the sleeping sickness focus of Zimbabwe in Hurungwe District. Parasites Vectors 2016, 9, 605. [Google Scholar] [CrossRef]
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef]
- Muhanguzi, D.; Mugenyi, A.; Bigirwa, G.; Kamusiime, M.; Kitibwa, A.; Akurut, G.G.; Ochwo, S.; Amanyire, W.; Okech, S.G.; Hattendorf, J.; et al. African animal trypanosomiasis as a constraint to livestock health and production in Karamoja region: A detailed qualitative and quantitative assessment. BMC Veter-Res. 2017, 13, 355. [Google Scholar] [CrossRef]
- Kabayo, J.P. Aiming to eliminate tsetse from Africa. Trends Parasitol. 2002, 18, 473–475. [Google Scholar] [CrossRef]
- Diall, O.; Cecchi, G.; Wanda, G.; Argilés-Herrero, R.; Vreysen, M.J.; Cattoli, G.; Viljoen, G.J.; Mattioli, R.; Bouyer, J. Developing a progressive control pathway for African animal trypanosomosis. Trends Parasitol. 2017, 33, 499–509. [Google Scholar] [CrossRef]
- Hocking, K.S.; Lamerton, J.F.; Lewis, E.A. Tsetse-fly control and eradication. Bull. World Health Organ. 1963, 28, 811–823. [Google Scholar]
- Vreysen, M.J.; Seck, M.T.; Sall, B.; Bouyer, J. Tsetse flies: Their biology and control using area-wide integrated pest management approaches. J. Invertebr. Pathol. 2012, 112, S15–S25. [Google Scholar] [CrossRef]
- Green, C.H. Bait methods for tsetse fly control. In Advances in Parasitology; Baker, J.R., Muller, R., Rollinson, D., Eds.; Elsevier: Amsterdam, The Netherlands, 1994; Volume 34, pp. 229–291. [Google Scholar]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Vreysen, M.J. Principles of area-wide integrated tsetse fly control using the sterile insect technique. Med. Trop. Rev. Corps Sante Colon. 2001, 61, 397–411. [Google Scholar]
- Vreysen, M.J.B.; Saleh, K.M.; Ali, M.Y.; Abdulla, A.M.; Zhu, Z.-R.; Juma, K.G.; Dyck, V.A.; Msangi, A.R.; Mkonyi, P.A.; Feldmann, H.U. Glossina austeni (Diptera: Glossinidae) eradicated on the Island of Unguja, Zanzibar, using the sterile insect technique. J. Econ. Èntomol. 2000, 93, 123–135. [Google Scholar] [CrossRef]
- Ciss, M.; Bassène, M.D.; Seck, M.T.; Mbaye, A.G.; Sall, B.; Fall, A.G.; Vreysen, M.J.B.; Bouyer, J. Environmental impact of tsetse eradication in Senegal. Sci. Rep. 2019, 9, 20313. [Google Scholar] [CrossRef]
- Takken, W. The Sterile Insect Technique for Tsetse Eradication in Nigeria; Cavalloro, R., Ed.; Commission of the European Communities: Brussels, Belgium, 1986; pp. 83–92. [Google Scholar]
- Alemu, T.; Kapitano, B.; Mekonnen, S.; Aboset, G.; Kiflom, M.; Bancha, B.; Woldeyes, G.; Bekele, K.; Feldmann, U. Area-wide control of tsetse and trypanosomosis: Ethiopian experience in the Southern Rift Valley. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 325–335. [Google Scholar]
- Parker, A.G.; Mamai, W.; Maiga, H. Mass-rearing for the sterile insect technique. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 283–316. [Google Scholar]
- Chambers, D.L. Quality control in mass rearing. Annu. Rev. Èntomol. 1977, 22, 289–308. [Google Scholar] [CrossRef]
- Jiang, L. Insights into the antiviral pathways of the silkworm bombyx mori. Front. Immunol. 2021, 12, 639092. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.; Cousserans, F.; Parker, A.G.; Jridi, C.; Bergoin, M.; Robinson, A.S. Quantitative PCR analysis of the salivary gland hypertrophy virus (GpSGHV) in a laboratory colony of Glossina pallidipes. Virus Res. 2009, 139, 48–53. [Google Scholar] [CrossRef]
- Eilenberg, J.; Vlak, J.; Nielsen-Leroux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. J. Insects Food Feed. 2015, 1, 87–102. [Google Scholar] [CrossRef]
- Lecocq, A.; Jensen, A.B.; Eilenberg, J. Diseases of insects in European production systems: Diagnosis, prevention and man-agement. Berl. Münch. Tierärztl. Wochenschr. 2019, 132, 245–250. [Google Scholar]
- Yang, S.; Zhao, Y.; Yu, J.; Fan, Z.; Gong, S.-T.; Tang, H.; Pan, L. Establishment of viral infection and analysis of host-virus interaction in drosophila melanogaster. J. Vis. Exp. 2019, 145, e58845. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Ros, V.I. Viruses of insects reared for food and feed. J. Invertebr. Pathol. 2017, 147, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Agboli, E.; Leggewie, M.; Altinli, M.; Schnettler, E. Mosquito-specific viruses—Transmission and interaction. Viruses 2019, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Weissman, D.B.; Gray, D.; Pham, H.T.; Tijssen, P. Billions and billions sold: Pet-feeder crickets (Orthoptera: Gryllidae), commercial cricket farms, an epizootic densovirus, and government regulations make for a potential disaster. Zootaxa 2012, 3504, 67–88. [Google Scholar] [CrossRef]
- Abd-Alla, A.M.; Adun, H.; Parker, A.G.; Vreysen, M.J.; Bergoin, M. The antiviral drug valacyclovir successfully suppresses salivary gland hypertrophy virus (SGHV) in laboratory colonies of Glossina pallidipes. PLoS ONE 2012, 7, e38417. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G. Virus-like rods associated with salivary gland hyperplasia in tsetse, Glossina pallidipes. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 234–238. [Google Scholar] [CrossRef]
- Williams, T.; Virto, C.; Murillo, R.; Caballero, P. Covert infection of insects by baculoviruses. Front. Microbiol. 2017, 8, 1337. [Google Scholar] [CrossRef]
- Wojda, I. Temperature stress and insect immunity. J. Therm. Biol. 2017, 68, 96–103. [Google Scholar] [CrossRef]
- Carballo, A.; Williams, T.; Murillo, R.; Caballero, P. Iflavirus covert infection increases susceptibility to nucleopolyhedrovirus disease in Spodoptera exigua. Viruses 2020, 12, 509. [Google Scholar] [CrossRef]
- Demirbas-Uzel, G.; Kariithi, H.M.; Parker, A.G.; Vreysen, M.J.B.; Mach, R.L.; Abd-Alla, A.M.M. Susceptibility of tsetse species to Glossina pallidipes salivary gland hypertrophy virus (GpSGHV). Front. Microbiol. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Mutika, G.N.; Marin, C.; Parker, A.G.; Boucias, D.G.; Vreysen, M.J.B.; Abd-Alla, A. Impact of salivary gland hypertrophy virus infection on the mating success of male Glossina pallidipes: Consequences for the sterile insect technique. PLoS ONE 2012, 7, e42188. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, A.M.; Bergoin, M.; Parker, A.G.; Maniania, N.K.; Vlak, J.M.; Bourtzis, K.; Boucias, D.G.; Aksoy, S. Improving Sterile Insect Technique (SIT) for tsetse flies through research on their symbionts and pathogens. J. Invertebr. Pathol. 2013, 112, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, A.M.M.; Parker, A.G.; Vreysen, M.J.B.; Bergoin, M. Tsetse salivary gland hypertrophy virus: Hope or hindrance for tsetse control? PLoS Neglected Trop. Dis. 2011, 5, e1220. [Google Scholar] [CrossRef]
- Yimer, M.M.; Bula, D.G.; Tesama, T.K.; Tadesse, K.A.; Abera, B.H. Prevalence of salivary gland hypertrophy syndrome in laboratory colonies and wild flies of Glossina pallidipes in Ethiopia. Onderstepoort J. Veter-Res. 2015, 82, e1–e6. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.; Otieno, L.H.; Mwangi, R.W. The Effects of a DNA virus infection on the reproductive potential of female tsetse flies, Glossina morsitans centralis and Glossina morsitans morsitans (Diptera: Glossinidae). Mem. Inst. Oswaldo Cruz 1998, 93, 861–864. [Google Scholar] [CrossRef]
- Sang, R.C.; Jura, W.G.; Otieno, L.H.; Mwangi, R.W.; Ogaja, P. The effects of a tsetse DNA virus infection on the functions of the male accessory reproductive gland in the host fly Glossina morsitans centralis (Diptera; Glossinidae). Curr. Microbiol. 1999, 38, 349–354. [Google Scholar] [CrossRef]
- Meki, I.K.; Huditz, H.-I.; Strunov, A.; van der Vlugt, R.A.A.; Kariithi, H.M.; Rezapanah, M.; Miller, W.J.; Vlak, J.M.; van Oers, M.M.; Abd-Alla, A.M.M. Characterization and tissue tropism of newly identified iflavirus and negeviruses in Glossina morsitans morsitans tsetse flies. Viruses 2021, 13, 2472. [Google Scholar] [CrossRef]
- van Oers, M.M. Genomics and biology of iflaviruses. In Insect Virology; Johndon, K., Asgari, S., Eds.; Caister Academic Press: Norfolk, UK, 2010; pp. 231–250. [Google Scholar]
- Lu, G.; Ye, Z.-X.; He, Y.-J.; Zhang, Y.; Wang, X.; Huang, H.-J.; Zhuo, J.-C.; Sun, Z.-T.; Yan, F.; Chen, J.-P.; et al. Discovery of two novel negeviruses in a dungfly collected from the arctic. Viruses 2020, 12, 692. [Google Scholar] [CrossRef]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribution. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef]
- Chen, Y.P.; Becnel, J.J.; Valles, S.M. RNA viruses infecting pest insects. In Insect Pathology, 2nd ed.; Vega, F.E., Kaya, H.K., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 133–170. ISBN 9780123849847. [Google Scholar]
- Eberle, K.E.; Wennmann, J.T.; Kleespies, R.G.; Jehle, J.A. Basic techniques in insect virology. In Manual of Techniques in In-Vertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 15–74. ISBN 978-0-12-386899-2. [Google Scholar]
- Carrillo-Tripp, J.; Krueger, E.N.; Harrison, R.; Toth, A.L.; Miller, W.A.; Bonning, B.C. Lymantria dispar iflavirus 1 (LdIV1), a new model to study iflaviral persistence in lepidopterans. J. Gen. Virol. 2014, 95, 2285–2296. [Google Scholar] [CrossRef] [PubMed]
- Suvanto, M.T.; Nguyen, P.T.; Uusitalo, R.; Korhonen, E.M.; Faolotto, G.; Vapalahti, O.; Huhtamo, E.; Smura, T. A novel negevirus isolated from Aedes vexans mosquitoes in Finland. Arch. Virol. 2020, 165, 2989–2992. [Google Scholar] [CrossRef] [PubMed]
- Vreysen, M.J.B. Radiation Induced Sterility to Control Tsetse Flies. The Effect of Ionising Radiation and Hybridisation on Tsetse Biology and the Use of the Sterile Insect Technique in Integrated Tsetse Control. Ph.D. Thesis, Wageningen Agricultural University, Wageningen, The Netherlands, 1995. [Google Scholar]
- Bakri, A.; Mehta, K.; Lance, D.R. Sterilizing insects with ionizing radiation. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Sayed, W.A.A.-E.; El-Helaly, A.M.A. Effect of gamma irradiation on the susceptibility of the cotton leaf worm, Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae) to the infection with nucleopolyhedrosis virus. Egypt J. Biol. Pest Control 2018, 28, 1–4. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, S.; Abd-Alla, A.M.M.; Bourtzis, K. The effect of radiation on the gut bacteriome of Aedes albopictus. Front. Microbiol. 2021, 12, 671699. [Google Scholar] [CrossRef] [PubMed]
- Woruba, D.N.; Morrow, J.L.; Reynolds, O.L.; Chapman, T.A.; Collins, D.P.; Riegler, M. Diet and irradiation effects on the bacterial community composition and structure in the gut of domesticated teneral and mature Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 281. [Google Scholar] [CrossRef]
- Weiss, B.L.; Wang, J.; Aksoy, S. Tsetse immune system maturation requires the presence of obligate symbionts in Larvae. PLoS Biol. 2011, 9, e1000619. [Google Scholar] [CrossRef]
- Mirieri, C.K.; Mutika, G.N.; Bruno, J.; Seck, M.T.; Sall, B.; Parker, A.G.; van Oers, M.M.; Vreysen, M.J.B.; Bouyer, J.; Abd-Alla, A.M.M. A new automated chilled adult release system for the aerial distribution of sterile male tsetse flies. PLoS ONE 2020, 15, e0232306. [Google Scholar] [CrossRef]
- Seck, M.T.; Pagabeleguem, S.; Bassene, M.D.; Fall, A.G.; Diouf, T.A.R.; Sall, B.; Vreysen, M.J.B.; Rayaissé, J.-B.; Takac, P.; Sidibé, I.; et al. Quality of sterile male tsetse after long distance transport as chilled, irradiated pupae. PLoS Neglected Trop. Dis. 2015, 9, e0004229. [Google Scholar] [CrossRef]
- Datta, K.; Suman, S.; Kallakury, B.V.S.; Fornace, A.J. Exposure to heavy ion radiation induces persistent oxidative stress in mouse intestine. PLoS ONE 2012, 7, e42224. [Google Scholar] [CrossRef]
- López-Martínez, G.; Carpenter, J.E.; Hight, S.D.; Hahn, D.A. Low-oxygen atmospheric treatment improves the performance of irradiation-sterilized male cactus moths used in SIT. J. Econ. Èntomol. 2014, 107, 185–197. [Google Scholar] [CrossRef]
- López-Martínez, G.; Hahn, D.A. Short-term anoxic conditioning hormesis boosts antioxidant defenses, lowers oxidative damage following irradiation and enhances male sexual performance in the Caribbean fruit fly, Anastrepha suspensa. J. Exp. Biol. 2012, 215, 2150–2161. [Google Scholar] [CrossRef] [PubMed]
- Teets, N.M.; Dias, V.S.; Pierce, B.K.; Schetelig, M.F.; Handler, A.M.; Hahn, D.A. Overexpression of an antioxidant enzyme improves male mating performance after stress in a lek-mating fruit fly. Proc. R. Soc. B Boil. Sci. 2019, 286, 20190531. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, U. Guidelines for the rearing of tsetse flies using the membrane feeding technique. In Techniques of Insect Rearing for the Development of Integrated Pest and Vector Management Strategies; Ochieng’-Odero, J.P.R., Ed.; ICIPE Science Press: Nairobi, Kenya, 1994; Volume 2, pp. 449–471. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 15 August 2022)ISBN 978-3-319-24277-4.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-162. 2023. Available online: https://cran.r-project.org/package=nlme (accessed on 11 March 2023).
- Bartoń, K. MuMIn: Multi-Model Inference. 2018. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 August 2022).
- Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation. 2023. Available online: https://dplyr.tidyverse.org (accessed on 15 August 2022).
- Wright, M.N.; Ziegler, A. Ranger: A fast implementation of random forests for high dimensional data in C++ and R. J. Stat. Softw. 2017, 77, 1–17. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Xie, Y. Dynamic Documents with R and Knitr, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2015; ISBN 978-1498716963. [Google Scholar]
- Xie, Y.; Dervieux, C.; Riederer, E. R Markdown Cookbook; Chapman and Hall/CRC: Boca Raton, FL, USA, 2020; ISBN 9780367563837. [Google Scholar]
- Wang, F. Sending out alarms: A perspective on intercellular communications in insect antiviral immune response. Front. Immunol. 2021, 12, 613729. [Google Scholar] [CrossRef]
- Meki, I.K.; Ince, I.A.; Kariithi, H.; Boucias, D.G.; Ozcan, O.; Parker, A.G.; Vlak, J.M.; Van Oers, M.M.; Abd-Alla, A. Expression profile of Glossina pallidipes microRNAs during symptomatic and asymptomatic infection with Glossina pallidipes salivary gland hypertrophy virus (hytrosavirus). Front. Microbiol. 2018, 9, 2037. [Google Scholar] [CrossRef]
- Meki, I.K.; Kariithi, H.; Parker, A.G.; Vreysen, M.J.B.; Ros, V.I.D.; Vlak, J.M.; Van Oers, M.M.; Abd-Alla, A.M.M. RNA interference-based antiviral immune response against the salivary gland hypertrophy virus in Glossina pallidipes. BMC Microbiol. 2018, 18, 170. [Google Scholar] [CrossRef]
- Yue, C.; Schröder, M.; Gisder, S.; Genersch, E. Vertical-transmission routes for deformed wing virus of honeybees (Apis mellifera). J. Gen. Virol. 2007, 88, 2329–2336. [Google Scholar] [CrossRef]
- Rozo-Lopez, P.; Londono-Renteria, B.; Drolet, B.S. Impacts of infectious dose, feeding behavior, and age of Culicoides sonorensis biting midges on infection dynamics of vesicular stomatitis virus. Pathogens 2021, 10, 816. [Google Scholar] [CrossRef] [PubMed]
- Min, K.-J.; Tatar, M. Unraveling the molecular mechanism of immunosenescence in Drosophila. Int. J. Mol. Sci. 2018, 19, 2472. [Google Scholar] [CrossRef] [PubMed]
- Sciambra, N.; Chtarbanova, S. The impact of age on response to infection in Drosophila. Microorganisms 2021, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Morinet, F.; Parent, M.; Bergeron, C.; Pillet, S.; Capron, C. Oxygen and viruses: A breathing story. J. Gen. Virol. 2015, 96, 1979–1982. [Google Scholar] [CrossRef]
- Guo, Z.S. The impact of hypoxia on oncolytic virotherapy. Virus Adapt. Treat. 2011, 3, 71–82. [Google Scholar] [CrossRef]
- López-Martínez, G.; Hahn, D.A. Early life hormetic treatments decrease irradiation-induced oxidative damage, increase longevity, and enhance sexual performance during old age in the caribbean fruit fly. PLoS ONE 2014, 9, e88128. [Google Scholar] [CrossRef]
- Oliveira, M.F.; Geihs, M.A.; França, T.F.A.; Moreira, D.C.; Hermes-Lima, M. Is “preparation for oxidative stress” a case of physiological conditioning hormesis? Front. Physiol. 2018, 9, 945. [Google Scholar] [CrossRef]
- Benelli, M.; Mainali, B.; Taylor, P.W.; Rempoulakis, P. Reduced quality of sterile Queensland fruit fly following post-production stress from hypoxia, irradiation and vibration. J. Pest Sci. 2020, 94, 473–485. [Google Scholar] [CrossRef]
- Michalkova, V.; Benoit, J.B.; Attardo, G.M.; Medlock, J.; Aksoy, S. Amelioration of reproduction-associated oxidative stress in a viviparous insect is critical to prevent reproductive senescence. PLoS ONE 2014, 9, e87554. [Google Scholar] [CrossRef]
- Munks, R.J.L.; Sant’Anna, M.R.V.; Grail, W.; Gibson, W.; Igglesden, T.; Yoshiyama, M.; Lehane, S.M.; Lehane, M.J. Antioxidant gene expression in the blood-feeding fly Glossina morsitans morsitans. Insect Mol. Biol. 2005, 14, 483–491. [Google Scholar] [CrossRef]
- MacLEOD, E.T.; Maudlin, I.; Darby, A.C.; Welburn, S.C. Antioxidants promote establishment of trypanosome infections in tsetse. Parasitology 2007, 134, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Nims, R.W.; Gauvin, G.; Plavsic, M. Gamma irradiation of animal sera for inactivation of viruses and mollicutes—A review. Biologicals 2011, 39, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Aronstein, K.; Goblirsch, M.; Rinkevich, F.; de Guzman, L. Gamma irradiation inactivates honey bee fungal, microsporidian, and viral pathogens and parasites. J. Invertebr. Pathol. 2018, 153, 57–64. [Google Scholar] [CrossRef]
- Stiles, J.; Molyneux, D.H.; Wallbanks, K.R.; Van Der Vloedt, A.M.V. Effects of γ irradiation on the midgut ultrastructure of Glossina palpalis subspecies. Radiat. Res. 1989, 118, 353. [Google Scholar] [CrossRef] [PubMed]
- Langley, P.A.; Curtis, C.F.; Brady, J. The viability, fertility and behaviour of tsetse flies (Glossina morsitans) sterilized by irradiation under various conditions. Èntomol. Exp. Appl. 1974, 17, 97–111. [Google Scholar] [CrossRef]
- Mutika, G.N.; Parker, A.G. Tolerance of low temperature and sterilizing irradiation in males of Glossina pallidipes (Diptera: Glossinidae). J. Insect Sci. 2014, 14, 262. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Tang, T.; Song, Q.; Wang, Z.; He, K.; Liu, X.; Song, J.; Wang, L.; Yang, Y.; Feng, C. Transcription analysis of the stress and immune response genes to temperature stress in Ostrinia furnacalis. Front. Physiol. 2019, 10, 1289. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Genome | Primer Name | Sequence 5′ to 3′ End |
|---|---|---|
| β-tubulin (Tsetse) | Tse-TubqPCR-F | GATGGTCAAGTGCGATCCT |
| β-tubulin (Tsetse) | Tse-TubqPCR-R | TGAGAACTCGCCTTCTTCC |
| Iflavirus (Gmm) | Ifla_qPCR2_7848F | AGAAATTGAAGGACAGATGTTTGGT |
| Iflavirus (Gmm) | Ifla_qPCR2_7947R | ACCTAAGAAATTACCAGTACCCTCC |
| Negevirus (Gmm) | Nege-qPCR1-2411F | CAACATAGACTTGAACCAGAGCA |
| Negevirus (Gmm) | Nege-qPCR1-2529R | GAAACATCAAACACACTCCCATTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirieri, C.K.; Abd-Alla, A.M.M.; Ros, V.I.D.; van Oers, M.M. Evaluating the Effect of Irradiation on the Densities of Two RNA Viruses in Glossina morsitans morsitans. Insects 2023, 14, 397. https://doi.org/10.3390/insects14040397
Mirieri CK, Abd-Alla AMM, Ros VID, van Oers MM. Evaluating the Effect of Irradiation on the Densities of Two RNA Viruses in Glossina morsitans morsitans. Insects. 2023; 14(4):397. https://doi.org/10.3390/insects14040397
Chicago/Turabian StyleMirieri, Caroline K., Adly M.M. Abd-Alla, Vera I.D. Ros, and Monique M. van Oers. 2023. "Evaluating the Effect of Irradiation on the Densities of Two RNA Viruses in Glossina morsitans morsitans" Insects 14, no. 4: 397. https://doi.org/10.3390/insects14040397
APA StyleMirieri, C. K., Abd-Alla, A. M. M., Ros, V. I. D., & van Oers, M. M. (2023). Evaluating the Effect of Irradiation on the Densities of Two RNA Viruses in Glossina morsitans morsitans. Insects, 14(4), 397. https://doi.org/10.3390/insects14040397