
Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera in Open Biotopes




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Placement of Traps
2.2. Identification
2.3. Data Analyses
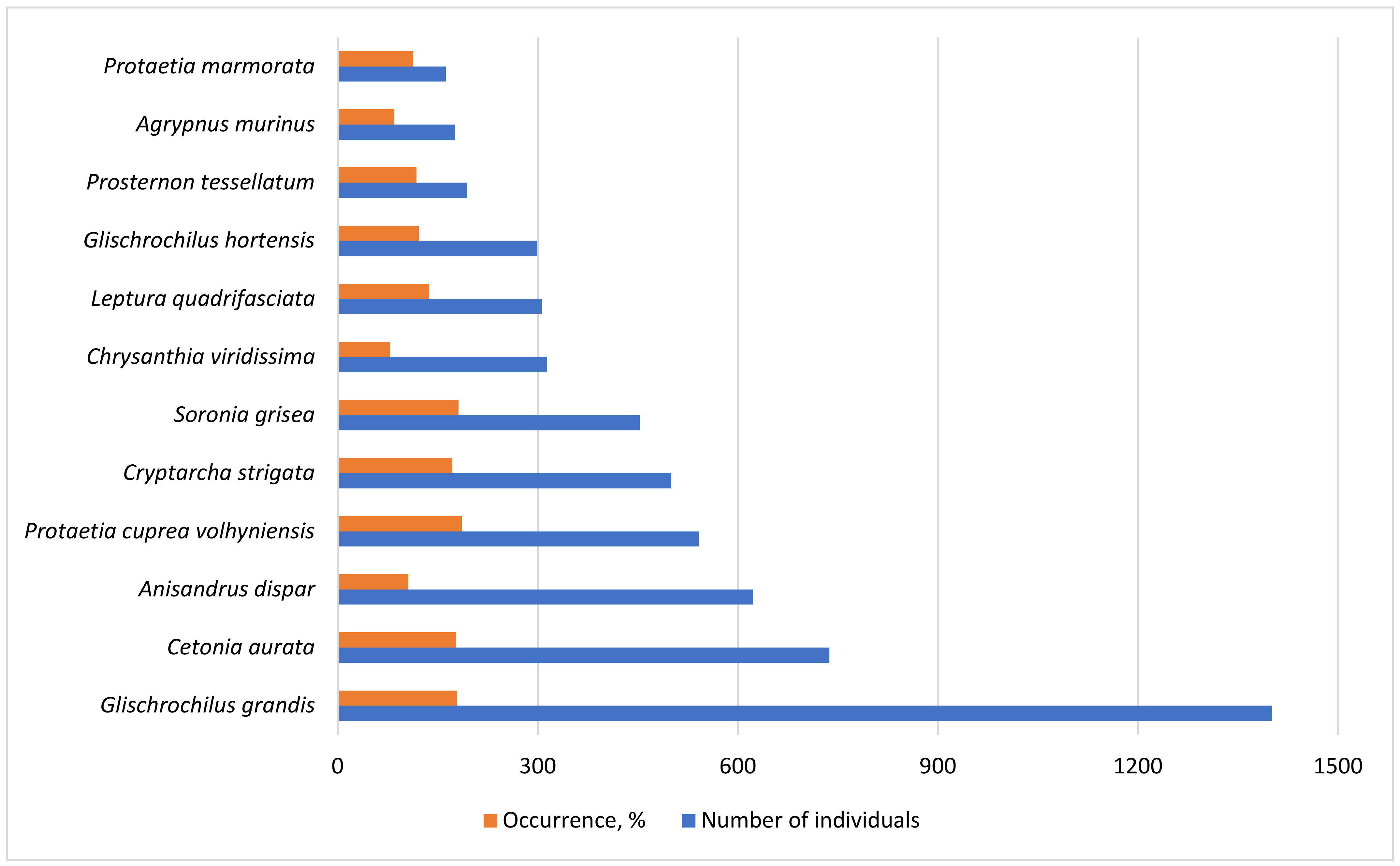
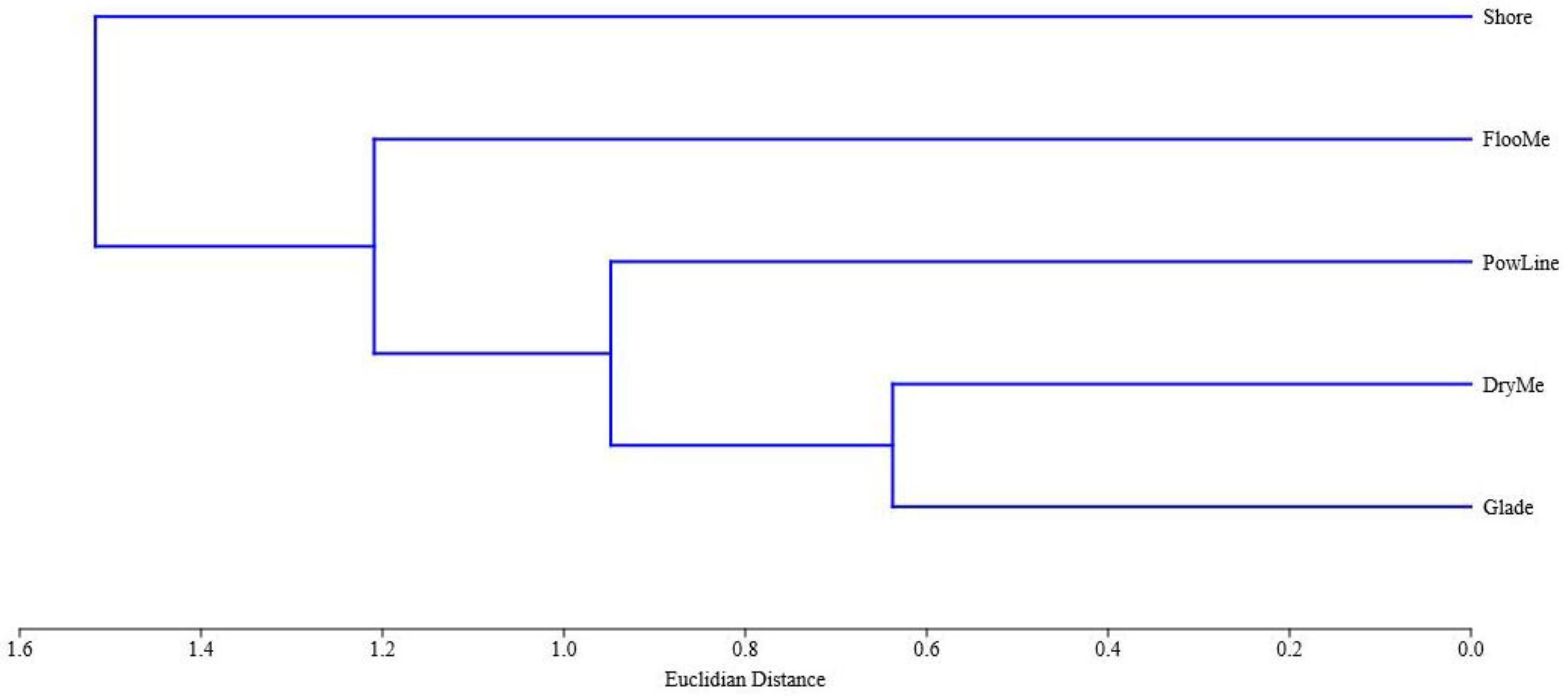
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family, Species | DryMe | Shore | FlooMe | PowLine | Glade | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | O | N | O | N | O | N | O | N | O | ||
| Carabidae | |||||||||||
| Agonum lugens (Duftschmid, 1812) | 1 | 2.0 | 1 | 1.4 | 2 | ||||||
| Amara aenea (De Geer, 1774) | 1 | 1.1 | 1 | ||||||||
| Amara tibialis (Paykull, 1798) | 2 | 2.2 | 1 | 2.0 | 3 | ||||||
| Cicindela hybrida (Linnaeus, 1758) | 2 | 5.0 | 2 | ||||||||
| Calathus fuscipes (Goeze, 1777) | 1 | 2.0 | 1 | ||||||||
| Calathus melanocephalus (Linnaeus, 1758) | 4 | 2.2 | 1 | 2.0 | 1 | 1.4 | 6 | ||||
| Harpalus luteicornis (Duftschmid, 1812) | 1 | 1.1 | 1 | ||||||||
| Harpalus signaticornis (Duftschmid, 1812) | 1 | 1.1 | 1 | ||||||||
| Lebia cruxminor (Linnaeus, 1758) | 1 | 1.1 | 1 | 1.8 | 2 | ||||||
| Lebia marginata (Geoffroy, 1785) | 1 | 1.4 | 1 | ||||||||
| Buprestidae | |||||||||||
| Agrilus sp. | 1 | 5.0 | 1 | ||||||||
| Anthaxia quadripunctata (Linnaeus, 1758) | 2 | 4.0 | 4 | 5.4 | 3 | 4.3 | 9 | ||||
| Chrysobothris chrysostigma (Linnaeus, 1758) | 2 | 4.0 | 1 | 1.8 | 3 | ||||||
| Throscidae | |||||||||||
| Trixagus sp. | 3 | 4.3 | 3 | ||||||||
| Lycidae | |||||||||||
| Lygistopterus sanguineus (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Cantharidae | |||||||||||
| Cantharis fusca (Linnaeus, 1758) | 1 | 1.1 | 10 | 5.7 | 11 | ||||||
| Cantharis livida (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Cantharis obscura (Linnaeus, 1758) | 2 | 1.4 | 2 | ||||||||
| Cantharis nigricans (O.F. Müller, 1776) | 2 | 2.2 | 1 | 2.0 | 2 | 2.8 | 5 | ||||
| Cantharis figurata (Mannerheim, 1843) | 1 | 5.0 | 3 | 4.0 | 4 | ||||||
| Cantharis pellucida (Fabricius, 1792) | 1 | 1.1 | 1 | ||||||||
| Cantharis rustica (Fallén, 1807) | 3 | 2.2 | 7 | 5.7 | 10 | ||||||
| Rhagonycha lignosa (O.F. Müller, 1764) | 1 | 2.0 | 1 | ||||||||
| Rhagonycha nigripes (W. Redtenbacher, 1842) | 1 | 1.8 | 1 | 1.4 | 2 | ||||||
| Rhagonycha nigriventris (Motschulsky, 1860) | 3 | 6.0 | 3 | ||||||||
| Elateridae | |||||||||||
| Actenicerus sjaelandicus (O.F. Müller, 1764) | 1 | 1.1 | 1 | 2.0 | 2 | ||||||
| Agriotes lineatus (Linnaeus, 1767) | 1 | 1.1 | 1 | 1.4 | 2 | ||||||
| Agriotes sputator (Linnaeus, 1758) | 1 | 2.0 | 1 | ||||||||
| Agrypnus murinus (Linnaeus, 1758) | 137 | 46.7 | 6 | 10.0 | 3 | 5.4 | 30 | 22.9 | 176 | ||
| Ampedus balteatus (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Ampedus cinnabarinus (Eschscholtz, 1829) | 3 | 3.3 | 1 | 1.8 | 6 | 4.3 | 10 | ||||
| Ampedus elongatulus (Fabricius, 1787) | 7 | 3.3 | 7 | ||||||||
| Ampedus nigroflavus (Goeze, 1777) | 1 | 1.1 | 1 | ||||||||
| Ampedus pomonae (Stephens, 1830) | 1 | 1.4 | 1 | ||||||||
| Ampedus pomorum (Herbst, 1784) | 8 | 6.7 | 3 | 2.8 | 11 | ||||||
| Ampedus praeustus (Fabricius, 1792) | |||||||||||
| Ampedus sanguinolentus (Schrank, 1776) | 5 | 5.6 | 2 | 4.0 | 3 | 2.8 | 10 | ||||
| Cidnopus aeruginosus (G.-A. Olivier, 1790) | 1 | 5.0 | 1 | ||||||||
| Ctenicera pectinicornis (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Dalopius marginatus (Linnaeus, 1758) | 1 | 1.8 | 1 | ||||||||
| Danosoma fasciatum (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Dicronychus equiseti (Herbst, 1784) | 8 | 5.6 | 1 | 5.0 | 9 | ||||||
| Hemicrepidius niger (Linnaeus, 1758) | 2 | 4.0 | 2 | ||||||||
| Limonius minutus (Linnaeus, 1758) | 3 | 4.3 | 3 | ||||||||
| Melanotus castanipes (Paykull, 1800) | 1 | 5.0 | 1 | 1.4 | 2 | ||||||
| Melanotus villosus (Geoffroy, 1785) | 1 | 1.1 | 3 | 5.4 | 3 | 4.3 | 7 | ||||
| Mosotalesus impressus (Fabricius, 1792) | 1 | 1.1 | 1 | ||||||||
| Pristilophus cruciatus (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Prosternon tessellatum (Linnaeus, 1758) | 27 | 22.2 | 25 | 16.0 | 44 | 42.9 | 98 | 37.1 | 194 | ||
| Selatosomus aeneus (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Histeridae | |||||||||||
| Gnathoncus buyssoni (Auzat, 1917) | 1 | 1.4 | 1 | ||||||||
| Margarinotus striola (C.R. Sahlberg, 1819) | 1 | 1.4 | 1 | ||||||||
| Paromalus parallelepipedus (Herbst, 1791) | 2 | 1.4 | 2 | ||||||||
| Saprinus rugifer (Paykull, 1809) | 1 | 2.0 | 1 | ||||||||
| Hydrochidae | |||||||||||
| Hydrochus brevis (Herbst, 1793) | 1 | 5.0 | 1 | ||||||||
| Staphylinidae | |||||||||||
| Oiceoptoma thoracicum (Linnaeus, 1758) | 6 | 15.0 | 11 | 14.0 | 28 | 17.1 | 45 | ||||
| Staphylinidae sp. | 13 | 10 | 38 | 45.0 | 36 | 40.0 | 18 | 23.2 | 195 | 34.3 | 300 |
| Quedius dilatatus (Fabricius, 1787) | 4 | 7.1 | 4 | ||||||||
| Scarabaeidae | |||||||||||
| Amphimallon altaicum (Mannerheim, 1825) | 1 | 2.0 | 1 | ||||||||
| Cetonia aurata (Linnaeus, 1758) | 155 | 48.9 | 1 | 5.0 | 36 | 26.0 | 490 | 57.1 | 55 | 40.0 | 737 |
| Gnorimus variabilis (Linnaeus, 1758) | |||||||||||
| Oxythyrea funesta (Poda von Neuhaus, 1761) | 8 | 8.9 | 1 | 2.0 | 139 | 66.1 | 6 | 7.1 | 154 | ||
| Phyllopertha horticola (Linnaeus, 1758) | 3 | 3.3 | 4 | 7.1 | 16 | 10.0 | 23 | ||||
| Protaetia fieberi (Kraatz, 1880) | 3 | 2.2 | 5 | 8.0 | 13 | 14.3 | 22 | 10.0 | 43 | ||
| Protaetia marmorata (Fabricus, 1792) | 12 | 12.2 | 2 | 10.0 | 6 | 10.0 | 47 | 33.9 | 95 | 47.1 | 162 |
| Protaetia speciosissima (Scopoli, 1786) | 1 | 1.8 | 1 | ||||||||
| Protaetia cuprea volhyniensis (Gory & Percheron, 1833) | 139 | 56.7 | 9 | 18.0 | 303 | 71.4 | 91 | 40.0 | 542 | ||
| Serica brunnea (Linnaeus, 1758) | 1 | 2.0 | 1 | ||||||||
| Trichius fasciatus (Linnaeus, 1758) | 1 | 1.1 | 6 | 10.7 | 9 | 8.6 | 16 | ||||
| Dermestidae | |||||||||||
| Attagenus schaefferi (Herbst, 1792) | 2 | 1.1 | 4 | 5.7 | 6 | ||||||
| Dermestes lardarius (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Dermestes murinus (Linnaeus, 1758) | 1 | 1.1 | 1 | 1.8 | 2 | ||||||
| Trogoderma glabrum (Herbst, 1783) | 3 | 3.3 | 1 | 2.0 | 4 | 4.3 | 8 | ||||
| Cleridae | |||||||||||
| Thanasimus formicarius (Linnaeus, 1758) | 9 | 8.6 | 9 | ||||||||
| Trichodes apiarius (Linnaeus, 1758) | 3 | 3.3 | 2 | 4.0 | 121 | 55.4 | 13 | 15.7 | 139 | ||
| Melyridae | |||||||||||
| Charopus flavipes (Paykull, 1798) | 1 | 1.4 | 1 | ||||||||
| Cordylepherus viridis (Fabricius, 1787) | 1 | 1.1 | 1 | 5.0 | 1 | 2.0 | 6 | 7.1 | 9 | ||
| Dasytes fusculus (Illiger, 1801) | 1 | 2.0 | 1 | ||||||||
| Dasytes niger (Linnaeus, 1761) | 2 | 2.2 | 2 | 5.0 | 22 | 20.0 | 1 | 1.8 | 14 | 15.7 | 41 |
| Dolichosoma lineare (P. Rossi, 1794) | 5 | 2.0 | 1 | 1.4 | 6 | ||||||
| Malachius aeneus (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Malachius bipustulatus (Linnaeus, 1758) | 1 | 1.1 | 3 | 15.0 | 6 | 6.0 | 3 | 4.3 | 13 | ||
| Mordellidae | |||||||||||
| Mordella sp. | 1 | 1.1 | 3 | 1.8 | 6 | 5.7 | 10 | ||||
| Mordellistena humeralis (Linnaeus, 1758) | 2 | 10.0 | 2 | ||||||||
| Mordellistena sp. | 1 | 2.0 | 1 | 1.4 | 2 | ||||||
| Tomoxia bucephala (A. Costa, 1854) | 1 | 1.1 | 3 | 4.3 | 4 | ||||||
| Variimorda sp. | 1 | 2.0 | 1 | ||||||||
| Scraptiidae | |||||||||||
| Anaspis frontalis (Linnaeus, 1758) | 17 | 10.0 | 2 | 2.8 | 19 | ||||||
| Anaspis thoracica (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Oedemeridae | |||||||||||
| Chrysanthia geniculata (W.L.E. Schmidt, 1846) | 9 | 8.9 | 6 | 2.8 | 15 | ||||||
| Chrysanthia viridissima (Linnaeus, 1758) | 92 | 8.9 | 1 | 2.0 | 214 | 60.7 | 7 | 7.1 | 314 | ||
| Oedemera femorata (Scopoli, 1763) | 2 | 4.0 | 1 | 1.8 | 5 | 7.1 | 8 | ||||
| Oedemera virescens (Linnaeus, 1767) | 1 | 1.1 | 2 | 4.0 | 4 | 5.7 | 7 | ||||
| Anthicidae | |||||||||||
| Notoxus monoceros (Linnaeus, 1761) | 2 | 2.2 | 4 | 15.0 | 1 | 2.0 | 3 | 1.8 | 41 | 8.6 | 51 |
| Melandryidae | |||||||||||
| Dircaea quadriguttata (Paykull, 1798) | 1 | 5.0 | 1 | ||||||||
| Mycetophagidae | |||||||||||
| Litargus connexus (Geoffroy, 1785) | 2 | 1.4 | 2 | ||||||||
| Tenebrionidae | |||||||||||
| Bolitophagus reticulatus (Linnaeus, 1767) | 1 | 2.0 | 1 | 1.4 | 2 | ||||||
| Isomira murina (Linnaeus, 1758) | 2 | 2.0 | 15 | 16.1 | 17 | ||||||
| Lagria hirta (Linnaeus, 1758) | 1 | 1.1 | 1 | 2.0 | 1 | 1.8 | 4 | 5.7 | 7 | ||
| Pseudocistela ceramboides (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Latridiidae | |||||||||||
| Corticaria sp. | 1 | 5.0 | 1 | ||||||||
| Coccinellidae | |||||||||||
| Ceratomegilla notata (Laicharting, 1781) | 3 | 4.3 | 3 | ||||||||
| Coccinella hieroglyphica (Linnaeus, 1758) | 1 | 2.0 | 1 | ||||||||
| Coccinella magnifica (L. Redtenbacher, 1843) | 11 | 16.1 | 2 | 2.8 | 13 | ||||||
| Coccinella quinquepunctata (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Coccinella septempunctata (Linnaeus, 1758) | 16 | 15.6 | 10 | 12.0 | 17 | 21.4 | 12 | 14.3 | 55 | ||
| Coccinula quatuordecimpustulata (Linnaeus, 1758) | 4 | 4.3 | 4 | ||||||||
| Exochomus quadripustulatus (Linnaeus, 1758) | 1 | 2.0 | 1 | ||||||||
| Hippodamia variegata (Goeze, 1777) | 1 | 5.0 | 1 | 1.8 | 2 | ||||||
| Hyperaspis concolor (Suffrian, 1843) | 2 | 4.0 | 2 | ||||||||
| Platynaspis luteorubra (Goeze, 1777) | 2 | 4.0 | 2 | ||||||||
| Propylea quatuordecimpunctata (Linnaeus, 1758) | 1 | 2.0 | 1 | 1.4 | 2 | ||||||
| Psyllobora vigintiduopunctata (Linnaeus, 1758) | 2 | 2.2 | 2 | 4.0 | 1 | 1.8 | 3 | 4.3 | 8 | ||
| Subcoccinella vigintiquatuorpunctata (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Tytthaspis gebleri (Mulsant, 1850) | 1 | 1.4 | 1 | ||||||||
| Erotylidae | |||||||||||
| Triplax russica (Linnaeus, 1758) | 2 | 2.2 | 1 | 1.8 | 1 | 1.4 | 4 | ||||
| Triplax rufipes (Fabricius, 1787) | 1 | 1.4 | 1 | ||||||||
| Monotomidae | |||||||||||
| Rhizophagus fenestralis (Linnaeus, 1758) | 1 | 5.0 | 13 | 5.7 | 14 | ||||||
| Nitidulidae | |||||||||||
| Carpophilus hemipterus (Linnaeus, 1758) | 2 | 2.2 | 2 | 4.0 | 4 | ||||||
| Carpophilus marginellus (Motschulsky, 1858) | 2 | 2.0 | 1 | 1.4 | 3 | ||||||
| Carpophilus sp. | 2 | 2.8 | 2 | ||||||||
| Cryptarcha strigata (Fabricius, 1787) | 53 | 21.1 | 10 | 45.0 | 23 | 30.0 | 50 | 35.7 | 364 | 40.0 | 500 |
| Cryptarcha undata (G.-A. Olivier, 1790) | 4 | 6.0 | 2 | 3.6 | 6 | ||||||
| Cychramus luteus (Fabricius, 1787) | 1 | 2.0 | 3 | 2.8 | 4 | ||||||
| Epuraea guttata (G.-A. Olivier, 1811) | 1 | 2.0 | 9 | 4.3 | 10 | ||||||
| Epuraea sp. | 54 | 20 | 37 | 45.0 | 32 | 28.0 | 5 | 5.4 | 108 | 21.4 | 236 |
| Glischrochilus grandis (Tournier, 1872) | 109 | 33.3 | 38 | 45.0 | 53 | 28.0 | 142 | 26.8 | 1059 | 45.7 | 1401 |
| Glischrochilus hortensis (Geoffroy, 1785) | 25 | 10 | 123 | 55.0 | 110 | 26.0 | 2 | 3.6 | 39 | 27.1 | 299 |
| Glischrochilus quadriguttatus (Fabricius, 1777) | 2 | 2.2 | 5 | 20.0 | 1 | 2.0 | 8 | ||||
| Glischrochilus quadripunctatus (Linnaeus, 1758) | 5 | 5.6 | 4 | 20.0 | 11 | 12.9 | 20 | ||||
| Glischrochilus quadrisignatus (Say, 1835) | 1 | 1.1 | 1 | 5.0 | 10 | 10.0 | 1 | 1.8 | 3 | 2.8 | 16 |
| Ipidia binotata (Reitter, 1875) | 1 | 5.0 | 1 | 2.0 | 2 | ||||||
| Meligethes sp. | 1 | 5.0 | 1 | 1.4 | 2 | ||||||
| Omosita depressa (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Omosita japonica (Reitter, 1874) | 1 | 2.0 | 1 | ||||||||
| Soronia grisea (Linnaeus, 1758) | 107 | 27.8 | 135 | 75.0 | 105 | 36.0 | 10 | 12.5 | 96 | 30.0 | 453 |
| Soronia punctatissima (Illiger, 1794) | 1 | 1.8 | 1 | ||||||||
| Cucujidae | |||||||||||
| Pediacus depressus (Herbst, 1797) | 1 | 1.4 | 1 | ||||||||
| Silvanidae | |||||||||||
| Ahasverus advena (Waltl, 1834) | 1 | 1.4 | 1 | ||||||||
| Psammoecus bipunctatus (Fabricius, 1792) | 1 | 2.0 | 1 | ||||||||
| Silvanus bidentatus (Fabricius, 1792) | 1 | 1.4 | 1 | ||||||||
| Phalacridae | |||||||||||
| Phalacrus sp. | 1 | 2.0 | 1 | ||||||||
| Laemophloeidae | |||||||||||
| Cryptolestes pusillus (Schoenherr, 1817) | 3 | 4.3 | 3 | ||||||||
| Cerambycidae | |||||||||||
| Aegomorphus clavipes (Schrank, 1781) | |||||||||||
| Acanthocinus aedilis (Linnaeus, 1758) | 3 | 4.3 | 3 | ||||||||
| Agapanthia villosoviridescens (De Geer, 1775) | 1 | 2.0 | 1 | ||||||||
| Anastrangalia reyi (L. Heyden, 1889) | 10 | 10.7 | 1 | 1.4 | 11 | ||||||
| Arhopalus rusticus (Linnaeus, 1758) | 1 | 1.8 | 1 | ||||||||
| Aromia moschata (Linnaeus, 1758) | 4 | 4.4 | 18 | 20.0 | 19 | 22.0 | 12 | 12.5 | 5 | 5.7 | 58 |
| Asemum striatum (Linnaeus, 1758) | 4 | 5.4 | 4 | ||||||||
| Chlorophorus herbstii (Brahm, 1790) | 1 | 1.8 | 1 | ||||||||
| Dinoptera collaris (Linnaeus, 1758) | 1 | 1.8 | 1 | ||||||||
| Leptura quadrifasciata (Linnaeus, 1758) | 60 | 14.4 | 4 | 20.0 | 3 | 6.0 | 118 | 55.4 | 121 | 41.4 | 306 |
| Leptura thoracica (Creutzer, 1799) | 10 | 11.1 | 1 | 5.0 | 1 | 1.8 | 17 | 11.4 | 29 | ||
| Lepturalia nigripes (De Geer, 1775) | 26 | 15.6 | 36 | 25 | 32 | 24.3 | 94 | ||||
| Lepturobosca virens (Linnaeus, 1758) | 1 | 1.8 | 1 | ||||||||
| Monochamus galloprovincialis pistor (Germar, 1818) | 1 | 1.4 | 1 | ||||||||
| Necydalis major (Linnaeus, 1758) | 1 | 2.0 | 1 | 1.8 | 2 | ||||||
| Nivellia sanguinosa (Gyllenhal, 1827) | 1 | 1.4 | 1 | ||||||||
| Pachyta quadrimaculata (Linnaeus, 1758) | 2 | 2.2 | 4 | 5.4 | 10 | 5.7 | 16 | ||||
| Plagionotus arcuatus (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Pseudovadonia livida (Fabricius, 1776) | 1 | 1.4 | 1 | ||||||||
| Purpuricenus globulicollis (Dejean, 1839) | 1 | 1.4 | 1 | ||||||||
| Purpuricenus kaehleri (Linnaeus, 1758) | 1 | 1.1 | 1 | 2.0 | 3 | 5.4 | 5 | ||||
| Rhagium inquisitor (Linnaeus, 1758) | 2 | 2.8 | 2 | ||||||||
| Rhagium mordax (De Geer, 1775) | 35 | 14.4 | 2 | 10.0 | 2 | 4.0 | 3 | 5.4 | 64 | 22.9 | 106 |
| Rutpela maculata (Poda von Neuhaus, 1761) | 2 | 2.2 | 46 | 37.5 | 1 | 1.4 | 49 | ||||
| Stenocorus meridianus (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Stenurella bifasciata (O.F. Müller, 1776) | 17 | 12.5 | 17 | ||||||||
| Stenurella melanura (Linnaeus, 1758) | 1 | 1.1 | 42 | 25 | 1 | 1.4 | 44 | ||||
| Stictoleptura maculicornis (De Geer, 1775) | 2 | 3.6 | 2 | 2.8 | 4 | ||||||
| Stictoleptura rubra (Linnaeus, 1758) | 6 | 10.7 | 3 | 2.8 | 9 | ||||||
| Stictoleptura variicornis (Dalman, 1817) | 1 | 1.8 | 1 | ||||||||
| Strangalia attenuata (Linnaeus, 1758) | 1 | 2.0 | 20 | 32.1 | 1 | 1.4 | 22 | ||||
| Xylotrechus antilope (Schoenherr, 1817) | 1 | 1.1 | 1 | ||||||||
| Xylotrechus arvicola (Olivier, 1795) | 1 | 1.8 | 1 | ||||||||
| Xylotrechus capricornus (Gebler, 1830) | 1 | 1.1 | 1 | 1.4 | 2 | ||||||
| Xylotrechus rusticus (Linnaeus, 1758) | 4 | 5.7 | 4 | ||||||||
| Chrysomelidae | |||||||||||
| Altica sp. | 1 | 5.0 | 1 | 1.4 | 2 | ||||||
| Bromius obscurus (Linnaeus, 1758) | 1 | 1.8 | 1 | 1.4 | 2 | ||||||
| Cassida vittata (Villers, 1789) | 1 | 5.0 | 1 | ||||||||
| Crepidodera fulvicornis (Fabricius, 1792) | 1 | 5.0 | 1 | ||||||||
| Cryptocephalus flavipes (Fabricius, 1781) | 1 | 1.8 | 1 | ||||||||
| Cryptocephalus sericeus (Linnaeus, 1758) | 1 | 1.8 | 1 | ||||||||
| Galerucella lineola (Fabricius, 1781) | 1 | 2.0 | 1 | ||||||||
| Gonioctena linnaeana (Schrank, 1781) | 5 | 15.0 | 5 | ||||||||
| Gonioctena viminalis (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Hypocassida subferruginea (Schrank, 1776) | 1 | 1.1 | 1 | 1.4 | 2 | ||||||
| Lema cyanella (Linnaeus, 1758) | 1 | 2.0 | 1 | ||||||||
| Lilioceris merdigera (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Lochmaea caprea (Linnaeus, 1758) | 1 | 5.0 | 2 | 4.0 | 3 | ||||||
| Spermophagus sericeus (Geoffroy, 1785) | 1 | 1.4 | 1 | ||||||||
| Attelabidae | |||||||||||
| Deporaus betulae (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Anthribidae | |||||||||||
| Platystomos albinus (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Tropideres albirostris (Schaller, 1783) | 4 | 6.0 | 2 | 2.8 | 6 | ||||||
| Brentidae | |||||||||||
| Eutrichapion ervi (Kirby, 1808) | 1 | 1.4 | 1 | ||||||||
| Protapion interjectum (Desbrochers des Loges, 1895) | 1 | 1.1 | 1 | ||||||||
| Protapion fulvipes (Geoffroy, 1785) | 1 | 1.4 | 1 | ||||||||
| Curculionidae | |||||||||||
| Anisandrus dispar (Fabricius, 1792) | 25 | 7.8 | 282 | 45.0 | 100 | 26.0 | 8 | 5.4 | 208 | 21.4 | 623 |
| Anthonomus phyllocola (Herbst, 1795) | 1 | 1.1 | 1 | 1.4 | 2 | ||||||
| Anthonomus pomorum (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Anthonomus rubi (Herbst, 1795) | 1 | 2.0 | 1 | ||||||||
| Cleopomiarus distinctus (Boheman, 1845) | 1 | 2.0 | 1 | ||||||||
| Dorytomus dorsalis (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Hylurgus ligniperda (Fabricius, 1787) | 1 | 1.1 | 1 | ||||||||
| Hypera conmaculata (Herbst, 1795) | 1 | 2.0 | 1 | ||||||||
| Larinus obtusus (Gyllenhal, 1835) | 1 | 1.8 | 1 | ||||||||
| Larinus sturnus (Schaller, 1783) | 1 | 1.4 | 1 | ||||||||
| Larinus turbinatus (Gyllenhal, 1835) | 1 | 1.4 | 1 | ||||||||
| Miarus ajugae (Herbst, 1795) | 1 | 1.1 | 2 | 2.8 | 3 | ||||||
| Mononychus punctumalbum (Herbst, 1784) | 1 | 2.0 | 1 | ||||||||
| Otiorhynchus ovatus (Linnaeus, 1758) | 1 | 1.1 | 1 | ||||||||
| Phyllobius argentatus (Linnaeus, 1758) | 2 | 2.8 | 2 | ||||||||
| Phyllobius maculicornis (Germar, 1823) | 4 | 2.2 | 1 | 1.4 | 5 | ||||||
| Phyllobius oblongus (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Phyllobius pomaceus (Gyllenhal, 1834) | 3 | 6.0 | 3 | ||||||||
| Phyllobius pyri (Linnaeus, 1758) | 32 | 15.6 | 1 | 5.0 | 9 | 10.0 | 4 | 5.7 | 46 | ||
| Pissodes castaneus (De Geer, 1775) | 11 | 6.7 | 11 | ||||||||
| Pissodes pini (Linnaeus, 1758) | 1 | 1.4 | 1 | ||||||||
| Polydrusus tereticollis (De Geer, 1775) | 1 | 1.1 | 1 | ||||||||
| Sitona lineatus (Linnaeus, 1758) | 1 | 5.0 | 1 | ||||||||
| Strophosoma capitatum (De Geer, 1775) | 6 | 6.7 | 6 | ||||||||
| Tachyerges decoratus (Germar, 1821) | 1 | 5.0 | 1 | 1.4 | 2 | ||||||
| Xyleborinus saxesenii (Ratzeburg, 1837) | 1 | 2.0 | 1 | ||||||||
| Total | 1275 | 751 | 747 | 2040 | 3093 | 7906 | |||||
References
- Hodge, S.; Marshall, S.A.; Oliver, H.; Berry, J.; Marris, J.; Andrew, I. A preliminary survey of the insects collected using mushroom baits in native and exotic New Zealand woodlands. N. Z. Entomol. 2010, 33, 43–54. [Google Scholar] [CrossRef]
- Fagundes, C.K.; Di Mare, R.A.; Wink, C.; Manfio, D. Diversity of the families of Coleoptera captured with pitfall traps in five different environments in Santa Maria, RS, Brazil. Braz. J. Biol. 2011, 71, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Schmeelk, T.C.; Millar, J.G.; Hanks, L.M. Influence of trap height and bait type on abundance and species diversity of cerambycid beetles captured in forests of East-Central Illinois. J. Econ. Entomol. 2016, 109, 1750–1757. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, J.T.; Pawlowski, S.P.; Mayo, P.D.; Silk, P.J.; Sweeney, J.D.; Hillier, N.K. Influence of trap colour, type, deployment height, and a host volatile on monitoring Orchestes fagi (Coleoptera: Curculionidae) in Nova Scotia, Canada. Can. Entomol. 2020, 152, 98–109. [Google Scholar] [CrossRef]
- Skvarla, M.J.; Larson, J.L.; Fisher, J.R.; Dowling, A.P.G. A review of terrestrial and canopy Malaise traps. Ann. Entomol. Soc. Am. 2021, 114, 27–47. [Google Scholar] [CrossRef]
- Ivanova, N.; Shashkov, M. Tree stand assessment before and after windthrow based on open-access biodiversity data and aerial photography. Nat. Conserv. Res. 2022, 7 (Suppl. 1), 52–63. [Google Scholar] [CrossRef]
- Ghassemi-Khademi, T.; Khosravi, R.; Sajjad, A. Climate niche modeling of Scorpio kruglovi (Scorpiones: Scorpionidae) in Iran. J. Wildl. Biodivers. 2022, 6, 87–101. [Google Scholar] [CrossRef]
- Dedyukhin, S.V. Fauna and biotopic distribution of weevils (Coleoptera: Curculionoidea) of the Zhiguli State Nature Reserve, Russia. Nat. Conserv. Res. 2022, 7, 55–69. [Google Scholar] [CrossRef]
- Marques, M.I.; Adis, J.; Dos Santos, G.B.; Battirola, L.D. Terrestrial arthropods from tree canopies in the Pantanal of Mato Grosso, Brazil. Rev. Bras. Entomol. 2006, 50, 257–267. [Google Scholar] [CrossRef]
- Hodge, S.; Williams, A. Beetles collected using rotting vegetable baits in a Kent Garden. Entomol. Mon. Mag. 2011, 146, 179–188. [Google Scholar]
- Parhomenko, O.; Langraf, V.; Petrovičová, K.; Komlyk, V.; Brygadyrenko, V. Morphometric variability of ground beetles Bembidion minimum (Coleoptera, Carabidae): Who should change more, males or females? Nat. Conserv. Res. 2022, 7, 42–69. [Google Scholar] [CrossRef]
- Allemand, R.; Aberlenc, H.-P. Une méthode efficace d’echantillonage de l’entomofaune des frondaisons: Le piège attractif aérien. Bull. Soc. Entomol. Suisse 1991, 64, 293–305. [Google Scholar]
- DeVries, P.J.; Murray, D.; Lande, R. Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian rainforest. Biol. J. Linn. Soc. 1997, 62, 343–364. [Google Scholar] [CrossRef]
- De Zan, L.R.; Bardiani, M.; Antonini, G.; Campanaro, A.; Chiari, S.; Mancini, E.; Maura, M.; Sabatelli, S.; Solano, E.; Zauli, A.; et al. Guidelines for the monitoring of Cerambyx cerdo. Nat. Conserv. 2017, 20, 129–164. [Google Scholar] [CrossRef]
- MacRae, T.C.; Rice, M.E. Distributional and biological observations on North American Cerambycidae (Coleoptera). Coleopt. Bull. 2007, 61, 227–263. [Google Scholar] [CrossRef]
- Rukavina, I.; Kostanjšek, F.; Jelaska, S.D.; Pirnat, A.; Šerić Jelaska, L. Distribution and habitat suitability of two rare saproxylic beetles in Croatia–a piece of puzzle missing for South-Eastern Europe. IForest 2018, 11, 765–774. [Google Scholar] [CrossRef]
- Barros, R.C.; Fonseca, M.G.; Jardim, M.T.; Vendramini, V.E.; Damiani, B.C.B.; Julio, C.E.A. Species of Cerambycinae (Insecta, Coleoptera, Cerambycidae) from east Paraná State (Brazil), with new geographic records. Zootaxa 2020, 4845, 1–25. [Google Scholar] [CrossRef]
- Dvořák, L.; Dvořáková, K.; Oboňa, J.; Ruchin, A.B. Selected Diptera families caught with beer traps in the Republic of Mordovia (Russia). Nat. Conserv. Res. 2020, 5, 65–77. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. On the use of wine vinegar as an attractant in crown traps. Proc. Mord. State Nat. Res. 2021, 29, 3–12. [Google Scholar]
- Ruchin, A.B.; Esin, M.N. Seasonal dynamics of Diptera in individual biotopes in the center of the European part of Russia. Biosyst. Divers. 2021, 29, 374–379. [Google Scholar] [CrossRef]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Ried, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [PubMed]
- Putchkov, A.V.; Brygadyrenko, V.V. Rare species of Carabidae and Cicindelidae in Dnipropetrovsk Region. Biosyst. Divers. 2022, 30, 310–337. [Google Scholar] [CrossRef]
- Anselmo, L.; Rizzioli, B. Side threats: Further possible effects of warming on the high alpine narrow endemic Carabus cychroides (Coleoptera: Carabidae). Nat. Conserv. Res. 2022, 7, 88–94. [Google Scholar] [CrossRef]
- Sundukov, Y.N.; Makarov, K.V. The ground beetles of the tribus Trechini (Carabidae) on the Southern Kuril Islands. Nat. Conserv. Res. 2021, 6, 15–51. [Google Scholar] [CrossRef]
- Bondarenko, A.S.; Zamotajlov, A.S.; Belyi, A.I.; Khomitskiy, E.E. Fauna and ecological characteristics of ground beetles (Coleoptera, Carabidae) of the Nature Sanctuaries «Prichernomorskiy» and «Tuapsinskiy» (Russia). Nat. Conserv. Res. 2020, 5, 66–85. [Google Scholar] [CrossRef]
- Avtaeva, T.A.; Sukhodolskaya, R.A.; Brygadyrenko, V.V. Modeling the bioclimatic range of Pterostichus melanarius (Coleoptera, Carabidae) in conditions of global climate change. Biosyst. Divers. 2021, 29, 140–150. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Alekseev, S.K.; Khapugin, A.A. Post-fire fauna of carabid beetles (Coleoptera, Carabidae) in forests of the Mordovia State Nature Reserve (Russia). Nat. Conserv. Res. 2019, 4 (Suppl. 1), 11–20. [Google Scholar] [CrossRef]
- Sergeev, M.E. Species composition and biotopic distribution of leaf beetles (Coleoptera: Megalopodidae, Chrysomelidae) in the Sikhote-Alin State Nature Reserve (Russia). Nat. Conserv. Res. 2020, 5, 80–88. [Google Scholar] [CrossRef]
- Asbeck, T.; Großmann, J.; Paillet, Y.; Winiger, N.; Bauhus, J. The use of tree-related microhabitats as forest biodiversity indicators and to guide integrated forest management. Curr. For. Rep. 2021, 7, 59–68. [Google Scholar] [CrossRef]
- Dunn, E.; Hough-Goldstein, J.; Hanks, L.M.; Millar, J.G.; D’Amico, V. Range of attraction of pheromone lures and dispersal behavior of cerambycid beetles. Ann. Entomol. Soc. Am. 2016, 109, 872–880. [Google Scholar] [CrossRef]
- Kozel, P.; Sebek, P.; Platek, M.; Benes, J.; Zapletal, M.; Dvorsky, M.; Lanta, V.; Dolezal, J.; Bace, R.; Zbuzek, B.; et al. Connectivity and succession of open structures as a key to sustaining light-demanding biodiversity in deciduous forests. J. Appl. Ecol. 2021, 58, 2951–2961. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Dung beetles in continuous forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodivers. Conserv. 2002, 11, 1903–1918. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A.; Vikhrev, N.E.; Esin, M.N. The use of simple crown traps for the insects collection. Nat. Conserv. Res. 2020, 5, 87–108. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. Vertical stratification of beetles in deciduous forest communities in the centre of European Russia. Diversity 2021, 13, 508. [Google Scholar] [CrossRef]
- Cai, C.; Tihelka, E.; Giacomelli, M.; Lawrence, J.F.; Ślipiński, A.; Kundrata, R.; Yamamoto, S.; Thayer, M.K.; Newton, A.F.; Leschen, R.A.B.; et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. R. Soc. Open Sci. 2022, 9, 211771. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrichh, F.; Letsch, H.; et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef]
- Löbl, I.; Smetana, A. (Eds.) Curculionoidea I. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2011; Volume 7, p. 373. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Curculionoidea II. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2013; Volume 8, p. 707. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Hydrophiloidea–Staphylinoidea. In Catalogue of Palaearctic Coleoptera; Revised and Updated Version; Brill: Leiden, The Netherlands; Boston, MA, USA, 2015; Volume 2/1, p. 1702. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Scarabaeoidea–Scirtoidea–Dascilloidea–Buprestoidea–Byrrhoidea. In Catalogue of Palaearctic Coleoptera; Revised and Updated Version; Brill: Leiden, The Netherlands; Boston, MA, USA, 2016; Volume 3, p. 983. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Archostema-ta–Adephaga–Myxophaga. In Catalogue of Palaearctic Coleoptera; Revised and Updated Version; Brill: Leiden, The Netherlands; Boston, MA, USA,, 2017; Volume 1, p. 1443. [Google Scholar]
- Iwan, D.; Löbl, I. (Eds.) Tenebrionoidea. In Catalogue of Palaearctic Coleoptera; Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 5, p. 945. [Google Scholar]
- Danilevsky, M. (Ed.) Chrysomeloidea I (Vesperidae, Disteniidae, Cerambycidae). In Catalogue of Palaearctic Coleoptera; Updated and Revised Second Edition; Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 6/1, p. 712. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Elateroidea–Derodontoidea–Bostrichoidea–Lymexyloidea–Cleroidea–Cucujoidea. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2007; Volume 4, p. 935. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Chrysomeloidae. In Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2010; Volume 6, p. 924. [Google Scholar]
- Robertson, J.; Ślipiński, A.; Moulton, M.; Shockley, F.W.; Giorgi, A.; Lord, N.P.; McKenna, D.D.; Tomaszewska, W.; Forrest-er, J.; Miller, K.B.; et al. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst. Entomol. 2015, 40, 745–778. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Ko-ro-tyaev, B.; Lyal, C.H.C.; et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea. Monogr. Electrón. SEA 2017, 8, 1–729. [Google Scholar]
- Bousquet, Y. Litteratura Coleopterologica (1758–1900): A guide to selected books related to the taxonomy of Coleoptera with publication dates and notes. ZooKeys 2016, 583, 1–776. [Google Scholar] [CrossRef]
- Oleksa, A.; Chybicki, I.J.; Gawronski, R.; Svensson, G.P.; Burczyk, J. Isolation by distance in saproxylic beetles may increase with niche specialization. J. Insects Conserv. 2013, 17, 219–233. [Google Scholar] [CrossRef]
- Nikitsky, N.B. The Beetles (Insecta, Coleoptera) of the Moscow Oblast, Part 1; Direct-Media: Moscow, Russia, 2016; 712p. [Google Scholar]
- Nikitsky, N.B. The Beetles (Insecta, Coleoptera) of the Moscow Oblast, Part 2; Direct-Media: Moscow, Russia, 2019; 808p. [Google Scholar]
- Olenici, N.; Fodor, E. The diversity of saproxylic beetles’ from the Natural Reserve Voievodeasa forest, North-Eastern Romania. Ann. For. Res. 2021, 64, 31–60. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest Sci. 2021, 94, 871–883. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–659. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Chapman & Hall: London, UK, 1988; p. 179. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Champlain, A.B.; Knull, J.N. Fermenting bait traps for trapping Elateridae and Cerambycidae (Coleop.). Entomol. News 1932, 43, 253–257. [Google Scholar]
- Frost, S.W. New records from bait traps (Dipt.; Coleop.; Corrodentia). Entomol. News 1937, 48, 201–202. [Google Scholar]
- Frost, S.W.; Dietrich, H. Coleoptera taken from bait-traps. Ann. Entomol. Soc. Am. 1929, 22, 427–436. [Google Scholar] [CrossRef]
- Daily, G.C.; Ehrlich, P.R. Preservation of biodiversity in small rainforest patches: Rapid evaluations using butterfly trapping. Biodivers. Conserv. 1995, 4, 35–55. [Google Scholar] [CrossRef]
- Wong, J.C.H.; Hanks, L.M. Influence of fermenting bait and vertical position of traps on attraction of cerambycid beetles to pheromone lures. J. Econ. Entomol. 2016, 109, 2145–2150. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Solodovnikov, A.Y.; Antropov, A.V. Abundance patterns of Quedius dilatatus Leach (Coleoptera, Staphylinidae) and Vespa crabro L. (Hymenoptera, Vespidae) in Central European Russia suggest close adaptation of the inquiline rove beetle life cycle to the nest dynamics of its wasp host. Entomol. Rev. 2022, 102, 958–970. [Google Scholar] [CrossRef]
- Dvořák, L.; Ruchin, A.B.; Egorov, L.V.; Aleksanov, V.V.; Alekseev, S.K.; Shulaev, N.V.; Zakharova, E.Y. Distribution of species from the genus Panorpa (Mecoptera, Panorpidae) in European Russia except the Caucasus. Nat. Conserv. Res. 2023, 8, 24–33. [Google Scholar] [CrossRef]
- Makarkin, V.N.; Ruchin, A.B.; Lukyanova, Y.A. Neuropteran Assemblage (Insecta) of a Pine Forest in the Republic of Tatarstan Revealed by Crown Bait Traps. Contemp. Probl. Ecol. 2023, 16, 142–148. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Usage of fermental traps for studying the species diversity of Coleoptera. Insects 2021, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Ruchin, A.B.; Egorov, L.V.; Artaev, O.N.; Esin, M.N. Dataset: Coleoptera (Insecta) collected from beer traps in “Smolny” National Park (Russia). Data 2022, 7, 161. [Google Scholar] [CrossRef]
- Karolyi, F.; Gorb, S.N.; Krenn, H.W. Trapping pollen by the moist mouth: Structure and function of the mouthparts in the flower visiting Cetonia aurata (Scarabeidae, Coleoptera). Arthropod-Plant Interact. 2009, 3, 1–8. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Polumordvinov, O.A. Coleoptera of the Penza region, Russia based on fermental crown trap). Biodiversitas 2021, 22, 1946–1960. [Google Scholar] [CrossRef]
- Landvik, M.; Niemelä, P.; Roslin, T. Mother knows the best mould: An essential role for non-wood dietary components in the life cycle of a saproxylic scarab beetle. Oecologia 2016, 182, 163–176. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. Vertical stratification and seasonal dynamics of Coleoptera in open biotopes of forest ecosystems (Centre of European Russia). Forests 2022, 13, 1014. [Google Scholar] [CrossRef]
- Vuts, J.; Imrei, Z.; Toth, M. Development of an attractant-baited trap for Oxythyrea funesta Poda (Coleoptera: Scarabaeidae, Cetoniinae). Z. Nat. C 2008, 63, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Subchev, M.; Toshova, T.; Andreev, R.; Petrova, V.; Maneva, V.; Spasova, T.; Marinova, N.; Minkov, P.; Velchev, D. Using floral baited colour traps for detection and seasonal monitoring of Oxythyrea funesta (Poda) (Coleoptera: Cetoniidae) in Bulgaria. Acta Zool. Bulg. 2012, 64, 439–443. [Google Scholar]
- Egorov, L.; Ruchin, A.; Esin, M.; Artaev, O. Biodiversity of Coleoptera (Insecta) in Mordovia State Nature Reserve (Russia) using fermental traps. Biodivers. Data J. 2022, 10, e96989. [Google Scholar] [CrossRef] [PubMed]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Vertical distribution of beetles (Coleoptera) in pine forests in Central European Russia. Diversity 2022, 14, 622. [Google Scholar] [CrossRef]
- Tauzin, P. Ethologie et chorologie de Protaetia (Liocola) lugubris Herbst, 1786 sur le territoire français (Coleoptera, Cetoniidae, Cetoniinae, Cetoniini). Cetoniimania 2006, 3, 4–38. [Google Scholar]
- Urban, P.; Schulze, W. Ein aktueller Nachweis des Marmorierten Rosenkäfers Protaetia marmorata (Fabricius, 1792) in der Senne (Nordrhein-Westfalen) (Mitteilungen zur Insektenfauna Westfalens XXII). Mitt. ArbGem. Westfäl. Entomol. 2017, 33, 15–19. [Google Scholar]
- Bardiani, M.; Tini, M.; Carpaneto, G.M.; Audisio, P.; Bussola, E.; Campanaro, A.; Cini, A.; Maurizi, E.; Mason, F.; Peverieri, G.S.; et al. Effects of trap baits and height on stag beetle and lower chafer monitoring: Ecological and conservation implications. J. Insect Conserv. 2017, 21, 157–168. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; MacGowan, I.; Makarkin, V.N.; Antropov, A.V.; Gornostaev, N.G.; Khapugin, A.A.; Dvořák, L.; Esin, M.N. Post-fire insect fauna explored by crown fermental traps in forests of the European Russia. Sci. Rep. 2021, 11, 21334. [Google Scholar] [CrossRef] [PubMed]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze Coleoptera, Scarabaeoidea, Dascilloidea, Byrrhoidea i Parnoidea. Kat. Fauny Polski 1983, 23, 9. [Google Scholar]
- Parmentier, T.; Dekoninck, W.; Wenseleers, T. A highly diverse microcosm in a hostile world: A review on the associates of red wood ants (Formica rufa group). Insectes Sociaux 2014, 61, 229–237. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Semishin, G.B. Fauna of click beetles (Coleoptera: Elateridae) in the interfluve of Rivers Moksha and Sura, Republic of Mordovia, Russia. Biodiversitas 2018, 19, 1352–1365. [Google Scholar] [CrossRef]
- Koval, A.G.; Guseva, O.G. Click Beetles (Coleoptera, Elateridae) in Agrolandscapes of Northwestern Russia. Entmol. Rev. 2019, 99, 744–752. [Google Scholar] [CrossRef]
- Dettner, K.; Beran, A. Chemical Defense of the Fetid Smelling Click beetle Agrypnus murinus (Coleoptera: Elateridae). Entomol. Gen. 2000, 25, 27–32. [Google Scholar] [CrossRef]
- Traugott, M.; Schallhart, N.; Kaufmann, R.; Juen, A. The feeding ecology of elaterid larvae in central European arable land: New perspectives based on naturally occurring stable isotopes. Soil Biol. Biochem. 2008, 40, 342–349. [Google Scholar] [CrossRef]
- Zaharia, L. Studies regarding the diversity and abundance of Elaterid species (Coleoptera, Elateridae) in the soil of deciduous hill forest ecosystem from Moldavia, Romania. IUFRO Work. Party 2006, 7, 246–252. [Google Scholar]
- Gerstmeier, R. Checkered Beetles: Illustrated Key to Cleridae and Thanerocleridae of the Western Palaearctic; Margraf Verlag: Weirsheim, Germany, 1998. [Google Scholar]
- Flügel, H.J. Über einige Fundnachweise und den Blütenbesuch von Trichodes-Arten (Coleoptera: Cleridae). Philippia 2014, 16, 155–171. [Google Scholar]
- Kurochkin, A.S. Fauna and bionomy of sap beetles (Coleoptera, Nitidulidae) and kateretid beetles (Coleoptera, Kateretidae) of Krasnosamarskoe forestry farm (Samara Region, Russia). Vestnik of Samara University. Nat. Sci. Ser. 2007, 8, 120–128. [Google Scholar]
- Lasoń, A.; Holly, M. Glischrochilus grandis Tournier, 1872–new species of beetle for the Polish fauna and new data on the occurrence of genus Glischrochilus Reitter, 1873 (Coleoptera: Nitidulidae: Cryptarchinae). Acta Entomol. Sil. 2015, 23, 1–4. [Google Scholar]
- Oude, J.E. Naamlijst van de glanskevers van Nederland en het omliggende gebied (Coleoptera: Nitidulidae and Brachypteridae). Ned. Faunist. Meded. 1999, 8, 11–32. [Google Scholar]
- Avgin, S.S.; Antonini, G.; Lason, A.; Jansson, N.; Abacigil, T.Ö.; Varli, S.V.; Biase, A.; Audisio, P. New data on distribution, ecology, and taxonomy of Turkish Nitidulidae (Coleoptera). Turk. J. Zool. 2015, 39, 314–322. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Seasonal activity of Coleoptera attracted by fermental crown traps in forest ecosystems of Central Russia. Ecol. Quest. 2021, 32, 37–53. [Google Scholar] [CrossRef]
- Atanassova, J.; Sivilov, O. Pollen analysis of the crop contents of adult Oedemeridae (Coleoptera) in Bulgaria. Eur. J. Entomol. 2014, 111, 588–593. [Google Scholar] [CrossRef]
- Bílý, S.; Mehl, O. Longhorn Beetles (Coleoptera, Cerambycidae) of Fennoscandia and Denmark; Brill: Leiden, The Netherlands, 1989; p. 200. [Google Scholar]
- Babendreier, D.; Jeanneret, P.; Pilz, C.; Toepfer, S. Non-target effects of insecticides, entomopathogenic fungi and nematodes applied against western corn rootworm larvae in maize. J. Appl. Entomol. 2015, 139, 457–467. [Google Scholar] [CrossRef]
- Geiler, H. Beitrag zur Biologie und Phänologie von Notoxus monoceros L. (Coleoptera: Anthicidae). Beitr. Entomol. 1953, 3, 569–576. [Google Scholar]
- Klausnitzer, B. Beobachtungen zur Lebensweise von Meloe proscarabaeus Linnaeus, 1758 (Coleoptera: Meloidae). Gredleriana 2005, 5, 209–216. [Google Scholar]
- Balestra, G.M.; Bucini, D.; Paparatti, B.; Speranza, S.; Proietti Zolla, C.; Pucci, C.; Varvaro, L. Bio-etology of Anisandrus dispar F. and its possible involvment in dieback (Moria) disease of hazelnut (Corylus avellana L.) plants in central Italy. Acta Hortic. 2005, 686, 435–443. [Google Scholar]
- Saruhan, İ.; Akyol, H. Monitoring population density and fluctuations of Anisandrus dispar and Xyleborinus saxesenii (Coleoptera: Scolytinae, Curculionidae) in hazelnut orchards. Afr. J. Biotechnol. 2012, 11, 4202–4207. [Google Scholar]
- Mani, E.; Remund, U.; Schwaller, F. Der Ungleiche Holzbohrer, Xyleboryus dispar F. (Coleoptera: Scolytidae) im Obst und Weinbau. Landwirtsch. Schweiz. Bauer 1990, 3, 105–112. [Google Scholar]
- Ciglar, I.; Boric, B. Bark beetle (Scolytidae) in Croation orchards. Acta Hortic. 1998, 525, 299–305. [Google Scholar]







| No. | Biotopes | Brief Description |
|---|---|---|
| 1 | Dry meadow (DryMe) | It is a meadow with well-developed vegetation. The percent cover is about 80%. There are open ground gaps without herb cover. The vegetation is represented predominantly by grasses, with some ruderal plants (e.g., Agrimonia eupatoria, Cichorium intybus, and Erigeron annuus). The height of the herb cover is about 30 cm. The study site is surrounded by pine (Pinus sylvestris) undergrowth of 1.0–3.0 m height. |
| 2 | Shore (Shore) | It is a river sand bank, formed due to the river sediments of the sand. This site is 0.8 ha. Beer traps are located 5 m from the water. On one side, this site borders the river; on other sides, the site is surrounded by willow thickets. The soil is sandy and highly mobile. The herb vegetation is slightly pronounced, represented mainly by communities of Petasites spurius. |
| 3 | Floodplain meadow (FlooMe) | It is a floodplain meadow site with well-developed vegetation and sufficient humidity. The percent cover is about 95–100%. There are almost no open gaps of the ground. The height of the vegetation is up to 50–60 cm. The vegetation is represented mainly by cereal and forbs species. |
| 4 | Cuttings under power lines (PowLine) | In these habitats, cutting of trees and shrubs is constantly carried out to protect power lines. These habitats stretched along power lines are surrounded by forest ecosystems, mainly consisting of Pinus sylvestris and Betula pendula, with the participation of other deciduous species of trees and shrubs. In contrast to forest glades, these habitats have a linear allocation of open space (along power lines). The width of the open strip is 180–200 m. Low shrubs occur in some sites. Herb vegetation is very well expressed. The percent cover is up to 80%. There are open areas with bare soil here. |
| 5 | Forest glade (Glade) | These are open habitats surrounded by mixed or deciduous forests on all sides. The area ranges from 0.02 ha to 0.95 ha. Herb vegetation is well expressed. The percent cover is up to 90–100%. Areas with bare soil are few. |
| DryMe | Shore | FlooMe | PowLine | Glade | Total | |
|---|---|---|---|---|---|---|
| Number of traps | 90 | 20 | 50 | 56 | 70 | 286 |
| Total of individuals | 1275 | 751 | 747 | 2040 | 3093 | 7906 |
| Shannon index | 3.06 | 1.88 | 3.06 | 2.69 | 2.62 | |
| Simpson index | 0.07 | 0.25 | 0.09 | 0.11 | 0.18 | |
| Number of species (excluding unidentified ones) | 81 | 43 | 80 | 66 | 118 | 208 |
| Number of saproxylic | 44 | 23 | 34 | 45 | 67 | 106 |
| Number of saproxylic species (% of the total number of species per biotope) | 54.3 | 53.5 | 42.5 | 68.2 | 56.8 | 51.0 |
| Number of anthophilic | 44 | 19 | 42 | 45 | 43 | 91 |
| Number of anthophilic species (% of the total number of species per biotope) | 54.3 | 44.2 | 52.5 | 68.2 | 36.4 | 43.8 |
| Total of families | 21 | 18 | 23 | 19 | 31 | 35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera in Open Biotopes. Insects 2023, 14, 404. https://doi.org/10.3390/insects14040404
Ruchin AB, Egorov LV, Khapugin AA. Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera in Open Biotopes. Insects. 2023; 14(4):404. https://doi.org/10.3390/insects14040404
Chicago/Turabian StyleRuchin, Alexander B., Leonid V. Egorov, and Anatoliy A. Khapugin. 2023. "Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera in Open Biotopes" Insects 14, no. 4: 404. https://doi.org/10.3390/insects14040404
APA StyleRuchin, A. B., Egorov, L. V., & Khapugin, A. A. (2023). Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera in Open Biotopes. Insects, 14(4), 404. https://doi.org/10.3390/insects14040404