
A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae)

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cave | Geomorphic Characteristics | Habitat Where Troglodiplura Observed |
|---|---|---|---|
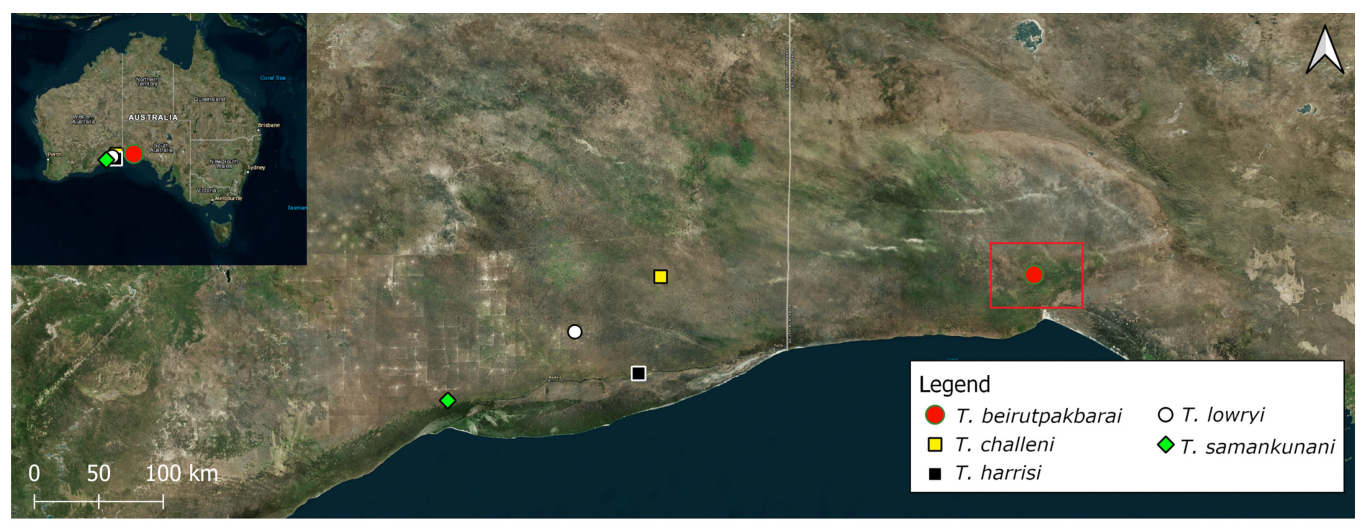
| T. beirutpakbarai Harvey & Rix, 2020 | N-253 Eagles Rest Cave (SA) | Complex cave ca. 250 m long under calcrete pavement. Single roof hole leads to a spacious chamber 60 m × 40 m × 12 m high and to a maze of passages. Ephemeral stream with ponding and significant speleothem development (calcite and halite). Some sediments have not received water ingress for some considerable time (Milner, pers. obs.). | ** Adult: dark zone on sediments. ** Adult desiccated fragments: dark zone on rocky boulder slope. ** Juveniles: diffused daylight zone near entrance, also on sediments in dark zone near live adult population. |
| T. challeni Harvey & Rix, 2020 | N-83 Old Homestead Cave (WA) | Highly complex cave with over 20 km passages. Cave is now very dry with desiccated sediments and rock piles. Abundant scattered speleothem development including calcite, gypsum, and halite. The cave shows geological evidence of former extensive ponding of still water (Milner, pers. obs.). | Dark zone up to 2 km from entrance [15] |
| T. harrisi Harvey & Rix, 2020 | N-327 Encompassing Cave (WA) | Complex cave, over 1 km long with mostly low passages. Calcite and other speleothem development. Tree roots penetrate the cave [57] | Habitat shared with Tartarus sp. in a sealed dark zone. Temperature 17.8–18.2 °C and humidity 80–90% [57] |
| T. lowryi Main 1969 | N-58 Roaches Rest Cave (WA) (Figure 1) | Large collapse chamber under doline lip with two roof holes. Cave is a 30° inclined chamber 90 m × 50 m × 13 m high. The cave floor is generally rocky, with reflected daylight reaching most aspects of the cave (Milner, pers. obs.). | Dark zone [15]. |
| T. samankunani Harvey & Rix, 2020 | N-49 Pannikin Plains Cave (WA) | Overhanging doline with very steep talus slope descends to the water table. Several kilometres of underwater passage development leads to air chambers accessible only by cave divers [58] | Dark zone in air chamber ca. 3 km north of cave entrance [15]. |
2. Materials and Methods
2.1. Study Area
2.2. Site Selection
2.3. Survey Methods
2.4. Site Characterisation
2.5. Molecular and Phylogenetic Methods
3. Results
3.1. Survey Results
| Cave | Troglodiplura beirutpakbarai Records | Geomorphic Characteristics | Cave Environment Where Troglodiplura Observed |
|---|---|---|---|
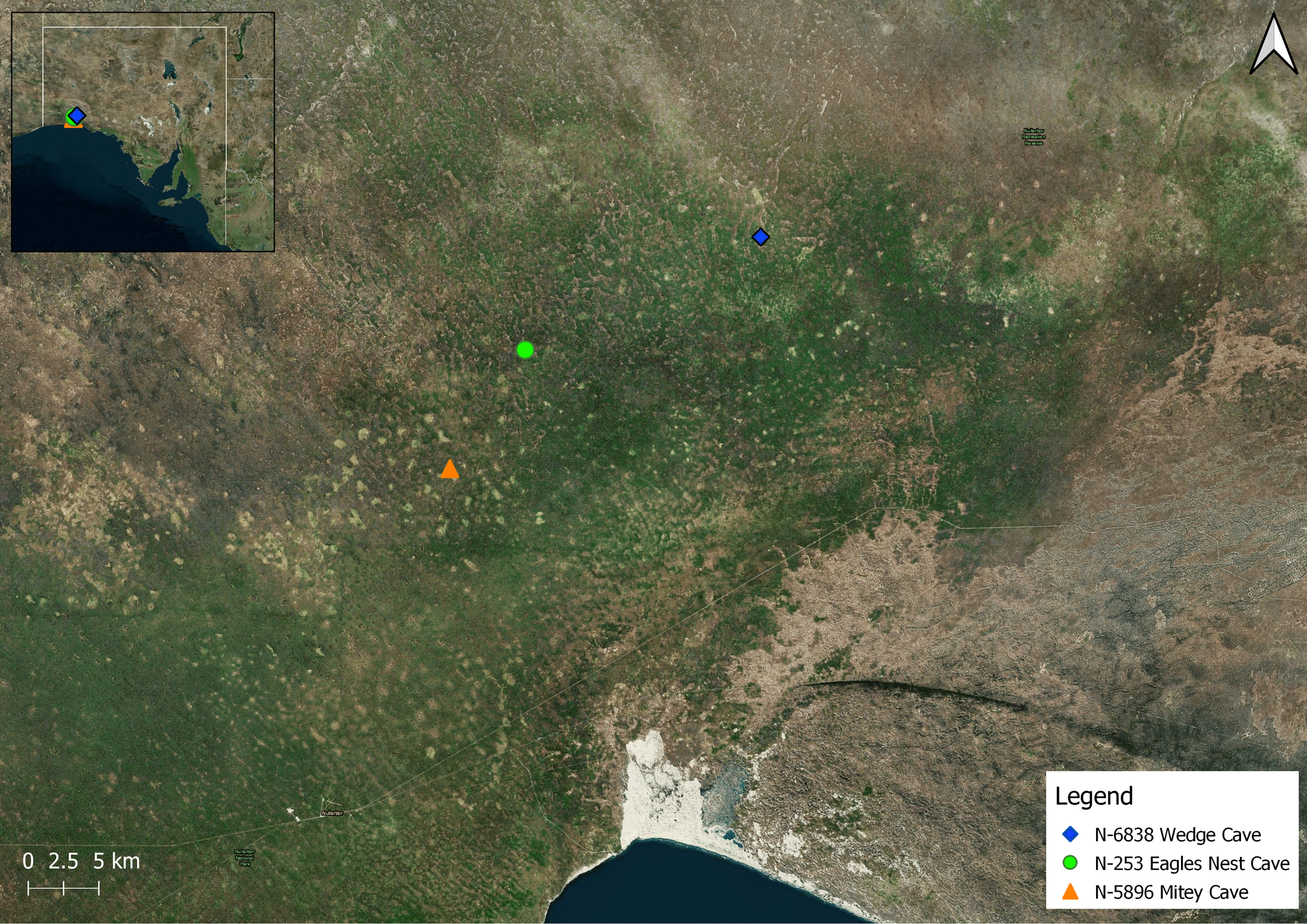
| N-253 Eagles Rest Cave (SA) | adult female (n = 2); juvenile (n = 11); desiccated, whole mature females (n = 3); fragments of exoskeleton, mature male (n = 4). Figure 5a,b. | Refer notes in Table 1. N-253 clearly takes a stream after rainfall events leaving ephemeral pools and calcite deposits; evidence of significant water flow and historical ponding in western passage. The deep cracks in the clay of the southern chamber floor suggest some considerable time since the last flood in that chamber. | Southern Chamber: 17.8 °C, 90% rh (8 March 2022) Western Passage where adults observed: 18.2 °C, 80% rh (8 March 2022) |
| N-6838 Wedge Cave (SA) | adult female (n = 2); juvenile (n = 8). | N-6838 is 18 km NE. from N-253. The collapse doline entrance rockpile descends to flat, wide, damp, hard, mud floor, ca. 10 m beneath the surface of limestone. Chamber is ca. 50 m × 30 m with a passage height of around 1.2 m (average). An occasional stream washes in with evidence of historical floods to 0.5 m deep. | 17.3 °C, 95% rh (7 March 2022) 17.5 °C, 90% rh (11 March 2022) |
| N-5896 Mitey Cave (SA) | juvenile (n = 2). | N-5896 is 10 km SW. from N-253. The 6 m deep entrance tube leads to a 60 m × 40 m chamber, 2–3 m high, with a flat floor. There is evidence of significant water ingress since 2015. | 18.5 °C, 76% rh (9 March 2022) |
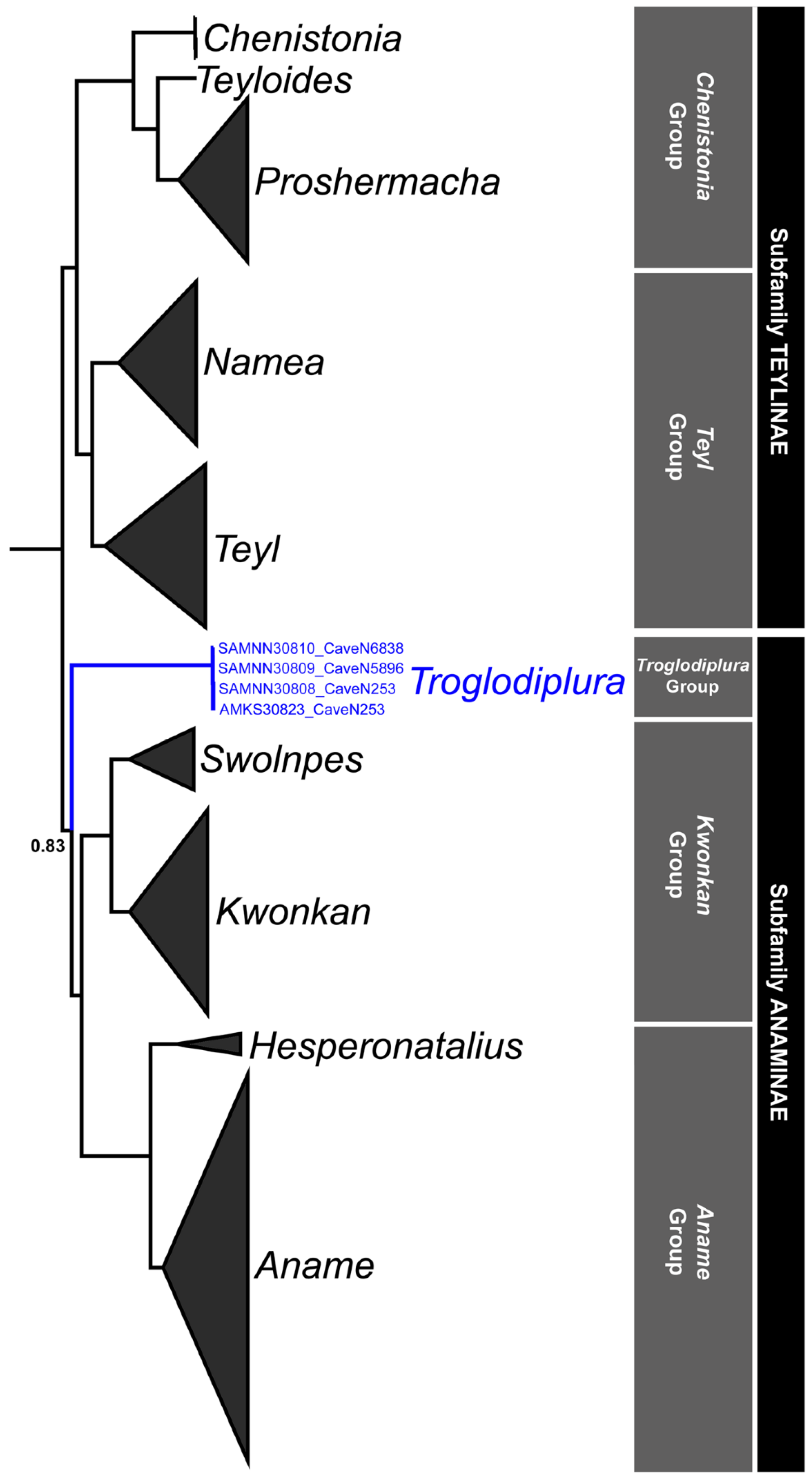
3.2. Phylogenetic Analysis
3.3. Intraspecific Variation
3.4. Biology
3.5. Conservation
3.6. Behaviour in Captivity
3.7. Symbiotic Mite
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lipar, M.; Ferk, M. Bihourly subterranean temperature and relative humidity data from the Nullarbor Plain, Australia (November 2019–March 2021). Data 2022, 7, 30. [Google Scholar] [CrossRef]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Miller, J.A. Cave adaptation in the spider genus Anthrobia (Araneae, Linyphiidae, Erigoninae). Zool. Scr. 2005, 34, 565–592. [Google Scholar] [CrossRef]
- Howarth, F.G. High-stress subterranean habitats and evolutionary change in cave-inhabiting arthropods. Am. Nat. 1993, 142, S65–S77. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S.; Isaia, M. Spiders in caves. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170193. [Google Scholar] [CrossRef] [PubMed]
- Rix, M.G.; Harvey, M.S. The spider family Micropholcommatidae (Arachnida, Araneae, Araneoidea): A relimitation and revision at the generic level. ZooKeys 2010, 36, 1–321. [Google Scholar] [CrossRef]
- Eberhard, S.M.; Richardson, A.; Swain, R. The Invertebrate Cave Fauna of Tasmania; Zoology Department, University of Tasmania: Hobart, Australia, 1991. [Google Scholar]
- Huber, B.A. Cave-dwelling pholcid spiders (Araneae, Pholcidae): A review. Subterr. Biol. 2018, 26, 1–18. [Google Scholar] [CrossRef]
- Gray, M. Cavernicolous spiders from the Nullarbor Plain and south-west Australia. Aust. J. Entomol. 1973, 12, 207–221. [Google Scholar] [CrossRef]
- Gray, M.R.; Thompson, J.A. New lycosoid spiders from cave and surface habitats in southern Australia and Cape Range peninsula (Araneae: Lycosoidea). Rec. West. Aust. Mus. 2001, 64, 159–170. [Google Scholar] [CrossRef]
- Kulkarni, S.; Hormiga, G. Hooroo mates! Phylogenomic data suggest that the closest relatives of the iconic Tasmanian cave spider Hickmania troglodytes are in Australia and New Zealand, not in South America. Invertebr. Syst. 2021, 35, 850–856. [Google Scholar] [CrossRef]
- Millidge, A. Three new species of the spider family Linyphiidae from Australia (Araneae). Rec. West. Aust. Mus. 1993, 16, 211–219. [Google Scholar]
- Harvey, M.S. Notes on the spider genus Symphytognatha (Araneae: Symphytognathidae) in Western Australia. Rec. West. Aust. Mus. 2001, 20, 345–348. [Google Scholar]
- Platnick, N.I. A new subterranean ground spider genus from Western Australia (Araneae: Trochanteriidae). Invertebr. Syst. 2008, 22, 295–299. [Google Scholar] [CrossRef]
- Harvey, M.S.; Rix, M.G.; Hillyer, M.J.; Huey, J.A. The systematics and phylogenetic position of the troglobitic Australian spider genus Troglodiplura (Araneae: Mygalomorphae), with a new classification for Anamidae. Invertebr. Syst. 2020, 34, 799–822. [Google Scholar] [CrossRef]
- Rix, M.G.; Edwards, D.L.; Byrne, M.; Harvey, M.S.; Joseph, L.; Roberts, J.D. Biogeography and speciation of terrestrial fauna in the south-western Australian biodiversity hotspot. Biol. Rev. 2015, 90, 762–793. [Google Scholar] [CrossRef]
- Webb, J.A.; James, J.M. Karst evolution of the Nullarbor plain, Australia. Spec. Pap. Geol. Soc. Am 2006, 404, 65. [Google Scholar] [CrossRef]
- Australian Bureau of Meteorology, Station Number 018106. Climate Statistics for. Australian Locations; Australian Government: Canberra, Australia, 2021. Available online: www.bom.gov.au/climate/average/tables/cw_200288_All.shtml (accessed on 28 August 2022).
- Richards, A.M. An ecological study of the cavernicolous fauna of the Nullarbor Plain Southern Australia. J. Zool. 1971, 164, 1–60. [Google Scholar] [CrossRef]
- Rix, M.G.; Huey, J.A.; Main, B.Y.; Waldock, J.M.; Harrison, S.E.; Comer, S.; Austin, A.D.; Harvey, M.S. Where have all the spiders gone? Highlighting the decline of a poorly known invertebrate fauna in the agricultural and arid zones of southern Australia. Austral. Entomol. 2017, 56, 14–22. [Google Scholar] [CrossRef]
- Rix, M.G.; Wilson, J.D.; Laidlaw, M.J.; Harvey, M.S.; Rix, A.G.; Rix, D.C. Demography, passive surveillance and potential habitat modelling of an Australian giant trapdoor spider (Idiopidae: Euoplos grandis) from the Queensland Brigalow Belt: Half a decade of population monitoring for conservation outcomes. Austral. Entomol 2023. [Google Scholar] [CrossRef]
- Howarth, F.G. Ecology of cave arthropods. Annu. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Matthews, T.J. On the biogeography of habitat islands: The importance of matrix effects, noncore species, and source-sink dynamics. Q. Rev. Biol. 2021, 96, 73–104. [Google Scholar] [CrossRef]
- Culver, D.C.; Master, L.L.; Christman, M.C.; Hobbs, H.H., III. Obligate cave fauna of the 48 contiguous United States. Cons. Biol. 2000, 14, 386–401. [Google Scholar] [CrossRef]
- Moulds, T.A.; Murphy, N.; Adams, M.; Reardon, T.; Harvey, M.S.; Jennings, J.; Austin, A.D. Phylogeography of cave pseudoscorpions in southern Australia. J. Biogeogr. 2007, 34, 951–962. [Google Scholar] [CrossRef]
- Balogh, A.; Ngo, L.; Zigler, K.S.; Dixon, G. Population genomics in two cave-obligate invertebrates confirms extremely limited dispersal between caves. Sci. Rep. 2020, 10, 11. [Google Scholar] [CrossRef]
- Snowman, C.V.; Zigler, K.S.; Hedin, M. Caves as islands: Mitochondrial phylogeography of the cave-obligate spider species Nesticus barri (Araneae: Nesticidae). J. Arachnol. 2010, 38, 49–56. [Google Scholar] [CrossRef]
- Derkarabetian, S.; Paquin, P.; Reddell, J.; Hedin, M. Conservation genomics of federally endangered Texella harvester species (Arachnida, Opiliones, Phalangodidae) from cave and karst habitats of central Texas. Conserv. Genet. 2022, 23, 401–416. [Google Scholar] [CrossRef]
- Hedin, M.C. Molecular phylogenetics at the population/species interface in cave spiders of the southern Appalachians (Araneae: Nesticidae: Nesticus). Mol. Biol. Evol. 1997, 14, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Cesaroni, D.; Allegrucci, G.; Caccone, A.; Cobolli Sbordoni, M.; De Matthaeis, E.; Di Rao, M.; Sbordoni, V. Genetic-variability and divergence between populations and species of Nesticus cave spiders. Genetica 1981, 56, 81–92. [Google Scholar] [CrossRef]
- Mammola, S.; Isaia, M.; Arnedo, M.A. Alpine endemic spiders shed light on the origin and evolution of subterranean species. PeerJ 2015, 3, e1384. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S. A spider species complex revealed high cryptic diversity in South China caves. Mol. Phylogenet. Evol. 2014, 79, 353–358. [Google Scholar] [CrossRef]
- Pavlek, M.; Gauthier, J.; Tonzo, V.; Bilat, J.; Arnedo, M.A.; Alvarez, N. Life-history traits drive spatial genetic structuring in Dinaric cave spiders. Front. Ecol. 2022, 10, 932. [Google Scholar] [CrossRef]
- Pavlek, M.; Mammola, S. Niche-based processes explaining the distributions of closely related subterranean spiders. J. Biogeogr. 2021, 48, 118–133. [Google Scholar] [CrossRef]
- May, R.M. Caves as islands. Nature 1977, 268, 589. [Google Scholar] [CrossRef]
- Culver, D.C. Analysis of simple cave communities.1. Caves as islands. Evolution 1970, 24, 463. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.C.; Holsinger, J.R. Speciation in cave faunas. Annu. Rev. Ecol. Evol. Syst. 1985, 16, 313–337. [Google Scholar] [CrossRef]
- Moldovan, O.T.; Meleg, I.N.; Persoiu, A. Habitat fragmentation and its effects on groundwater populations. Ecohydrology 2012, 5, 445–452. [Google Scholar] [CrossRef]
- Malard, F.; Capderrey, C.; Churcheward, B.; Eme, E.; Kaufmann, B.; Konecny-Dupré, L.; Léna, J.-P.; Liébault, F.; Douady, C.J. Geomorphic influence on intraspecific genetic differentiation and diversity along hyporheic corridors. Freshw. Biol. 2017, 62, 1955–1970. [Google Scholar] [CrossRef]
- Juan, C.; Guzik, M.T.; Jaume, D.; Cooper, S.J. Evolution in caves: Darwin’s ‘wrecks of ancient life’ in the molecular era. Mol. Ecol. 2010, 19, 3865–3880. [Google Scholar] [CrossRef]
- Faille, A.; Ribera, I.; Deharveng, L.; Bourdeau, C.; Garnery, L.; Quéinnec, E.; Deuve, T. A molecular phylogeny shows the single origin of the Pyrenean subterranean Trechini ground beetles (Coleoptera: Carabidae). Mol. Phylogenet. Evol. 2010, 54, 97–106. [Google Scholar] [CrossRef]
- Ribera, I.; Fresneda, J.; Bucur, R.; Izquierdo, A.; Vogler, A.P.; Salgado, J.M.; Cieslak, A. Ancient origin of a Western Mediterranean radiation of subterranean beetles. BMC Evol. Biol. 2010, 10, 29. [Google Scholar] [CrossRef]
- Arnedo, M.A.; Oromí, P.; Múrria, C.; Macías-Hernández, N.; Ribera, C. The dark side of an island radiation: Systematics and evolution of troglobitic spiders of the genus Dysdera Latreille (Araneae: Dysderidae) in the Canary Islands. Invertebr. Syst. 2007, 21, 623–660. [Google Scholar] [CrossRef]
- Hedin, M.; Derkarabetian, S.; Blair, J.; Paquin, P. Sequence capture phylogenomics of eyeless Cicurina spiders from Texas caves, with emphasis on US federally-endangered species from Bexar County (Araneae, Hahniidae). ZooKeys 2018, 769, 49–76. [Google Scholar] [CrossRef]
- Ledford, J.; Paquin, P.; Cokendolpher, J.; Campbell, J.; Griswold, C. Systematics of the spider genus Neoleptoneta Brignoli, 1972 (Araneae: Leptonetidae) with a discussion of the morphology and relationships for the North American Leptonetidae. Invertebr. Syst. 2011, 25, 334–388. [Google Scholar] [CrossRef]
- Mammola, S. Finding answers in the dark: Caves as models in ecology fifty years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.; Griebler, C.; Halse, S.; Humphreys, W.F.; et al. Scientists’ Warning on the Conservation of Subterranean Ecosystems. Bioscience 2019, 69, 641–650. [Google Scholar] [CrossRef]
- Badino, G. Cave temperatures and global climatic change. Int. J. Speleol. 2004, 33, 10. [Google Scholar] [CrossRef]
- Covington, M.D.; Perne, M. Consider a cylindrical cave: A physicist’s view of cave and karst science. Acta Carsologica 2015, 44, 1925. [Google Scholar] [CrossRef]
- Domínguez-Villar, D. Heat Flux; Wiley-Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]
- Mammola, S.; Goodacre, S.L.; Isaia, M. Climate change may drive cave spiders to extinction. Ecography 2018, 41, 233–243. [Google Scholar] [CrossRef]
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Domínguez-Villar, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Ribera, I.; Cieslak, A.; Faille, A.; Fresneda, J. Historical and ecological factors determining cave diversity. In Cave Ecology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 229–252. [Google Scholar] [CrossRef]
- Pallarés, S.; Colado, R.; Botella-Cruz, M.; Montes, A.; Balart-García, P.; Bilton, D.T.; Millán, A.; Ribera, I.; Sánchez-Fernández, D. Loss of heat acclimation capacity could leave subterranean specialists highly sensitive to climate change. Anim. Conserv. 2021, 24, 482–490. [Google Scholar] [CrossRef]
- Rix, M.G.; Wilson, J.D.; Huey, J.A.; Hillyer, M.J.; Gruber, K.; Harvey, M.S. Diversification of the mygalomorph spider genus Aname (Araneae: Anamidae) across the Australian arid zone: Tracing the evolution and biogeography of a continent-wide radiation. Mol. Phylogenet. Evol. 2020, 160, 107–127. [Google Scholar] [CrossRef]
- Poulter, N. Troglobitic Spiders of the Nullarbor Plain. In Under Way 2003: 24th Biennial Conference of the Australian Speleological Federation; Speleological Research Group Western Australia Inc.: East Victoria Park, Australia, 2003; pp. 83–85. [Google Scholar]
- Australian Speleological Federation, Karst Index Database. Available online: www.kid.caves.org.au/kid (accessed on 28 August 2022).
- Department of Climate Change, Energy, Environment and Water. Australia’s Bioregiions (IBRA). Available online: https://www.dcceew.gov.au/environment/land/nrs/science/ibra (accessed on 28 August 2022).
- Australian Speleological Federation. ASF Cave & Karst Numbering Code. 2006. Available online: www.caves.org.au/administration/codes-and-standards/send/8-codes-and-standards/50-asf-caves-and-karst-numbering-code. (accessed on 29 April 2023).
- Burnett, S.; Webb, J.A.; White, S. Shallow caves and blowholes on the Nullarbor Plain, Australia—Flank margin caves on a low gradient limestone platform. Geomorphology 2013, 201, 246–253. [Google Scholar] [CrossRef]
- Farrant, A.R.; Smart, P.L. Role of sediment in speleogenesis; sedimentation and paragenesis. Geomorphology 2011, 134, 79–93. [Google Scholar] [CrossRef]
- Calic-Ljubojevic, J. Bedding-plane anastomoses as one of the early stages of karst evolution. Acta Geol. Pol. 2002, 52, 111–115. [Google Scholar]
- Davey, A.G.; Gray, M.; Grimes, K.G.; Hamilton-Smith, E.; James, J.M.; Spate, A.P. World Heritage Significance of Karst and Other Landforms in the Nullarbor Region; Report; Commonwealth of Australia, Department of the Arts, Sport, the Environment and Territories: Canberra, Australia, 1992. [Google Scholar]
- Chapman, A.; Grafton, O. Guide to Best Practices for Generalising Sensitive Species Occurrence Data, Version 1.0; GBIF Secretariat: Copenhagen, Denmark, 2008. [Google Scholar]
- Rix, M.G.; Wilson, J.D.; Harvey, M.S. First phylogenetic assessment and taxonomic synopsis of the open-holed trapdoor spider genus Namea (Mygalomorphae: Anamidae): A highly diverse mygalomorph lineage from Australia’s tropical eastern rainforests. Invertebr. Syst. 2020, 34, 679–726. [Google Scholar] [CrossRef]
- Buzatto, B.A.; Haeusler, L.; Tamang, N. Trapped indoors? Long-distance dispersal in mygalomorph spiders and its effect on species ranges. J. Comp. Physiol. A 2021, 207, 279–292. [Google Scholar] [CrossRef]
- Paquin, P.; Hedin, M. The power and perils of ‘molecular taxonomy’: A case study of eyeless and endangered Cicurina (Araneae: Dictynidae) from Texas caves. Mol. Ecol. 2004, 13, 3239–3255. [Google Scholar] [CrossRef]
- Byrne, M.; Yeates, D.K.; Joseph, L.; Kearney, M.; Bowler, J.; Williams, M.A.J.; Cooper, S.; Donnellan, S.C.; Keogh, J.S.; Leys, R.; et al. Birth of a biome: Insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 2008, 17, 4398–4417. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, L.G.; James, N.P.; Bone, Y. The Miocene Nullarbor Limestone, southern Australia; deposition on a vast subtropical epeiric platform. Sediment. Geol. 2012, 253–254, 1–16. [Google Scholar] [CrossRef]
- Hampe, A.; Jump, A.S. Climate Relicts: Past, Present, Future. In Annual Review of Ecology, Evolution, and Systematics; Futuyma, D.J., Shaffer, H.B., Simberloff, D., Eds.; Annual Reviews: San Mateo, CA, USA, 2011; Volume 42, pp. 313–333. [Google Scholar] [CrossRef]
- Lee, D.C. The Rhodacaridae (Acari: Mesostigmata): Classification, external morphology and distribution of genera. Rec. South. Aust. Mus. 1970, 16, 1–219. Available online: https://www.biodiversitylibrary.org/item/127009#page/1/mode/1up (accessed on 29 April 2023).
- Beaulieu, F.; Walter, D.E.; Proctor, H.C.; Kitching, R.L. The canopy starts at 0.5 m: Predatory mites (Acari: Mesostigmata) differ between rainforest floor soil and suspended soil at any height. Biotropica 2010, 42, 704–709. [Google Scholar] [CrossRef]
- Dylewska, M.; Błoszyk, J.; Halliday, R.B. Three new species of mites in the genus Castriidinychus. Hirschmann from Tasmania (Acari: Uropodidae). Zootaxa 2010, 2546, 1–30. [Google Scholar] [CrossRef]
- Ebermann, E.; Goloboff, P.A. Association between Neotropical Burrowing Spiders (Araneae: Nemesiidae) and Mites (Acari: Heterostigmata, Scutacaridae). Acarologia 2002, 42, 173–184. [Google Scholar]
- Doran, N.E.; Kiernan, K.; Swain, R.; Richardson, A.M.M. Hickmania troglodytes, the Tasmanian cave spider, and its potential role in cave management. J. Insect. Conserv. 1999, 3, 257–262. [Google Scholar] [CrossRef]
- Marshall, B.M.; Strine, C.T.; Fukushima, C.S.; Cardoso, P.; Orr, M.C.; Hughes, A.C. Searching the web builds fuller picture of arachnid trade. Commun. Biol. 2022, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Morton, O.; Scheffers, B.R.; Haugaasen, T.; Edwards, D.P. Impacts of wildlife trade on terrestrial biodiversity. Nat. Ecol. Evol. 2021, 5, 540–548. [Google Scholar] [CrossRef] [PubMed]
- New, T. ‘Inordinate fondness’: A threat to beetles in south east Asia? J. Insect. Conserv. 2005, 9, 147–150. [Google Scholar] [CrossRef]
- Losey, J.E.; Chen, C.; Davis, A.E.; Deitsch, J.F.; Gertin, J.G.; Gorneau, J.A.; Hallock, E.M.; Jordán, J.P.; Kim, Z.J.; Kubinski, E.G.; et al. Insects and spiders on the web: Monitoring and mitigating online exploitation of species and services. Glob. Ecol. Conserv. 2022, 36, e02098. [Google Scholar] [CrossRef]
- Branco, V.V.; Cardoso, P. An expert-based assessment of global threats and conservation measures for spiders. Glob. Ecol. Conserv. 2020, 24, e01290. [Google Scholar] [CrossRef]
- Osborne, R.A.L. Saving and conserving the caves: Reflections on 37 years of listings, disputes, submissions and court cases. Aust. J. Earth. Sci. 2019, 66, 767–778. [Google Scholar] [CrossRef]
- Doherty, T.S.; Davis, R.A.; van Etten, E.J.; Algar, D.; Collier, N.; Dickman, C.R.; Edwards, G.; Masters, P.; Palmer, R.; Robinson, S. A continental-scale analysis of feral cat diet in Australia. J. Biogeogr. 2015, 42, 964–975. [Google Scholar] [CrossRef]
- Fleming, P.A.; Crawford, H.M.; Stobo-Wilson, A.M.; Dawson, S.J.; Dickman, C.R.; Dundas, S.J.; Gentle, M.N.; Newsome, T.M.; O’Connor, J.; Palmer, R.; et al. Diet of the introduced red fox Vulpes vulpes in Australia: Analysis of temporal and spatial patterns. Mammal. Rev. 2021, 51, 508–527. [Google Scholar] [CrossRef]
- Kutt, A.S. The diet of the feral cat (Felis catus) in north-eastern Australia. Acta Theriol. 2011, 56, 157–169. [Google Scholar] [CrossRef]
- Lunghi, E.; Valle, B.; Guerrieri, A.; Bonin, A.; Cianferoni, F.; Manenti, R.; Ficetola, G.F. Environmental DNA of insects and springtails from caves reveals complex processes of eDNA transfer in soils. Sci. Total Environ. 2022, 826, 154022. [Google Scholar] [CrossRef] [PubMed]







| Specimen & Depository | COI | 12S | 16S | H3 | 18S | 28S | EF-1γ |
|---|---|---|---|---|---|---|---|
| ARANEOMORPHAE HYPOCHILIDAE | |||||||
| Hypochilus Marx, 1888 [1] Outgroup | |||||||
| H. pococki (#chimera) | AF303512 | KY015437 | KY015913 | – | KY016493 | KY017132 | – |
| MYGALOMORPHAE—ATYPOIDEA ATYPIDAE Atypus Latreille, 1804 [2] | |||||||
| A. affinis (#ARANS000021) | KY017595 | KY015315 | KY015750 | – | KY016328 | KY016939 | – |
| A. karschi (#ARAMY002283) | – | – | KY015751 | – | DQ639769 | KY016940 | DQ680323 |
| Sphodros Walckenaer, 1835 [1] | |||||||
| S. atlanticus (#ARAMY000643) | KY017596 | KY015316 | KY015752 | KY018126 | – | KY016941 | – |
| MYGALOMORPHAE—AVICULARIOIDEA (NON-NEMESIOIDINA) EUAGRIDAE | |||||||
| Australothele Raven, 1984 [1] | |||||||
| A. jamiesoni (#ARAMY002084) | – | – | – | – | JX069739 | KY017061 | JQ358731 |
| Cethegus Thorell, 1881 [1] | |||||||
| C. fugax (WAM T129260) | KY295227 | KY320448 | KY320451 | KY295101 | KY294840 | KY294963 | – |
| PORRHOTHELIDAE Porrhothele Simon, 1892 [1] | |||||||
| P. sp. indet. (QMB S111386) | – | MT280964 | MT281032 | MT281253 | MT281107 | MT281179 | – |
| MYGALOMORPHAE—AVICULARIOIDEA (NEMESIOIDINA) ANAMIDAE Aname L. Koch, 1873 [20] | |||||||
| A. aragog (WAM T95409) | KJ745403 | KY214181 | KY241234 | KY241287 | KY241250 | KY241265 | MG800219 |
| A. ellenae (WAM T98890) | KJ745484 | KY214186 | KY241238 | KY241291 | KY241255 | KY241270 | – |
| A. exulans (WAM T121042) | MG800165 | MG799896 | MG799962 | MG800298 | MG800035 | MG800112 | MG800236 |
| A. grothi (WAM T133820) | MN635075 | MN634958 | MN634775 | MN635127 | MN634925 | MN634748 | – |
| A. lorica (WAM T113826) | KJ744825 | MN634945 | MN634841 | MN635093 | MN634935 | MN634733 | MN635140 |
| A. mainae (WAM T144398) | MN635077 | MN634938 | MN634859 | MN635092 | – | MN634735 | – |
| A. marae (WAM T98424) | KJ745450 | KY214185 | – | KY241290 | KY241254 | KY241269 | – |
| A. mccleeryorum (WAM T53979) | MT611168 | MT603527 | MT604146 | MT623663 | MT604125 | MT604135 | – |
| A. mellosa (WAM T107182) | KJ745440 | KY214184 | KY241237 | MG800294 | KY241253 | KY241268 | MG800231 |
| A. pallida (QMB S86817) | KY241278 | KY214179 | KY241230 | KY241283 | – | – | – |
| A. phillipae (WAM T110142) | – | – | MT604160 | MT623668 | MT604128 | MT604141 | MT623659 |
| A. salina (WAM T148204) | OQ922000 | OQ925882 | OQ925872 | OQ924319 | OQ925885 | OQ925881 | OQ924318 |
| A. simoneae (WAM T110261) | MT611175 | – | MT604158 | MT623669 | MT604129 | MT604144 | MT623660 |
| A. sinuata (WAM T129020) | MN635073 | MN634957 | MN634758 | MN635099 | MN634924 | MN634745 | MN635131 |
| A. vernonorum (WAM T98767) | MG800161 | MG799887 | MG799953 | MG800290 | MG800025 | MG800102 | MG800226 |
| A. watsoni (WAM T96018) | KJ745407 | MN634960 | MN634776 | MN635125 | MN634929 | MN634750 | – |
| A. whitei (WAM T127202) | KJ745174 | – | MN634900 | MN635108 | MN634928 | MN634726 | MN635134 |
| A. sp. ‘Goodnight’ (QMB S111402) | – | – | MT281027 | MT281268 | MT281106 | MT281182 | MT281249 |
| A. sp. ‘Goodnight’ (QMB S111405) | MT280892 | – | MT281026 | MT281267 | MT281105 | MT281181 | MT281248 |
| A. sp. ‘Paluma’ (QMB S111473) | – | MT280963 | MT281028 | MT281266 | MT281104 | MT281180 | – |
| Chenistonia Hogg, 1901 [3] | |||||||
| C. sp. ‘MYG348′ (WAM T72687) | KJ745221 | KY214180 | KY241231 | KY241284 | KY241247 | KY241262 | MG800196 |
| C. sp. ‘MYG348′ (WAM T81017) | MG800151 | MG799878 | MG799940 | MG800276 | MG800011 | MG800088 | MG800210 |
| C. sp. ‘MYG348′ (WAM T81018) | MG800152 | MG799879 | MG799941 | MG800277 | MG800012 | MG800089 | MG800211 |
| Hesperonatalius Castalanelli, Huey, Hillyer & Harvey, 2017 [2] | |||||||
| H. langlandsi (WAM T108988) | KJ744689 | KY214189 | KY241243 | – | KY241258 | KY241274 | MG800232 |
| H. maxwelli (WAM T108989) | KJ744690 | KY214190 | KY241244 | KY241293 | KY241259 | KY241275 | MG800233 |
| Kwonkan Main, 1983 [11] | |||||||
| K. turrigera (WAM T134203) | MG800182 | MG799911 | – | MG800313 | MG800056 | MG800134 | MG800254 |
| K. wonganensis (WAM T157120) | OQ922001 | OQ925883 | OQ925873 | OQ924322 | OQ925884 | OQ925877 | OQ924317 |
| K. sp. ‘MYG088’ (WAM T54237) | KJ745206 | MG799864 | MG799923 | MG800267 | MG799994 | MG800071 | MG800191 |
| K. sp. ‘MYG165’ (WAM T99672) | KJ745512 | KY214187 | KY241239 | KY241292 | KY241256 | KY241271 | MG800229 |
| K. sp. ‘MYG195’ (WAM T132749) | MG800177 | MG799906 | – | MG800308 | MG800049 | MG800127 | MG800250 |
| K. sp. ‘MYG197’ (WAM T130375) | MG800166 | – | MG799963 | MG800301 | MG800038 | MG800116 | MG800239 |
| K. sp. ‘MYG343’ (WAM T57563) | KJ745210 | MG799866 | MG799925 | – | MG799996 | MG800073 | MG800193 |
| K. sp. ‘MYG390’ (WAM T132361) | MG800173 | MG799902 | MG799970 | MG800304 | MG800045 | MG800123 | MG800247 |
| K. sp. ‘MYG392’ (WAM T132363) | MG800175 | MG799904 | MG799972 | MG800306 | MG800047 | MG800125 | – |
| K. sp. ‘MYG458’ (WAM T88514) | MG800155 | MG799880 | MG799944 | MG800280 | MG800015 | MG800092 | MG800213 |
| K. sp. ‘MYG650’ (WAM T145316) | MT656266 | MT656258 | – | – | – | – | – |
| Namea Raven, 1984 [9] | |||||||
| N. brisbanensis (QMB S111356) | MT280866 | MT280912 | MT281000 | – | MT281078 | MT281155 | MT281223 |
| N. flavomaculata (WAM T133310) | KY241282 | KY214192 | KY241246 | KY241294 | KY241261 | KY241277 | MG800253 |
| N. salanitri (QMB S111396) | MT280830 | MT280924 | MT280977 | – | MT281057 | MT281119 | MT281224 |
| N. dahmsi (QMB S111381) | MT280891 | – | MT281022 | MT281262 | MT281056 | MT281173 | MT281244 |
| N. jimna (QMB S111410) | MT280875 | – | MT281011 | – | MT281093 | MT281132 | MT281236 |
| N. excavans (QMB S111535) | MT280884 | MT280941 | MT281025 | – | – | – | – |
| N. sp. ‘Goomboor.’ (QMB S11135) | MT280881 | MT280950 | MT281013 | MT281261 | MT281089 | MT281139 | MT281238 |
| N. sp. ‘Kroombit’ (QMB S111330) | MT280887 | MT280960 | – | – | MT281051 | MT281171 | MT281231 |
| N. sp. ‘Ravensb.2’ (QMB S111445) | MT280883 | MT280942 | MT281019 | MT281264 | MT281043 | MT281175 | MT281228 |
| Proshermacha Simon, 1908 [9] | |||||||
| P. sp. ‘MYG344’ (WAM T132981) | MG800181 | – | MG799978 | MG800311 | MG800054 | MG800132 | MG800251 |
| P. sp. ‘MYG346’ (WAM T80952) | MG800150 | – | MG799939 | MG800275 | MG800010 | MG800087 | MG800209 |
| P. sp. ‘MYG349’ (WAM T72701) | KJ745222 | – | MG799928 | – | MG799999 | MG800076 | MG800197 |
| P. sp. ‘MYG357’ (WAM T78535) | KY241279 | – | KY241232 | KY241285 | KY241248 | KY241263 | MG800206 |
| P. sp. ‘MYG465’ (WAM T132960) | MG800180 | – | MG799977 | – | MG800053 | MG800131 | – |
| P. sp. ‘MYG467’ (WAM T131982) | MG800169 | – | MG799966 | – | MG800041 | MG800119 | MG800242 |
| P. sp. ‘MYG468’ (WAM T96060) | MG800158 | – | MG799948 | MG800285 | MG800020 | MG800097 | MG800220 |
| P. sp. ‘MYG469’ (WAM T94765) | MG800156 | – | MG799945 | MG800281 | MG800016 | MG800093 | MG800214 |
| P. sp. ‘MYG471’ (WAM T132903) | – | MG799907 | MG799974 | MG800309 | MG800050 | MG800128 | – |
| Swolnpes Main & Framenau, 2009 [4] | |||||||
| S. darwini (WAM T97003) | KY241280 | KY214183 | KY241236 | KY241289 | KY241252 | KY241267 | MG800223 |
| S. darwini (WAM T97503) | MG800160 | MG799884 | MG799950 | MG800287 | MG800022 | MG800099 | MG800224 |
| S. sp. ‘MYG234’ (WAM T114056) | – | MG799894 | – | – | MG800032 | MG800109 | – |
| S. sp. ‘MYG415’ (WAM T53579) | MG800145 | MG799863 | – | MG800266 | MG799993 | MG800070 | – |
| Teyl Main, 1975 [9] | |||||||
| T. damsonoides (WAM T137482) | MG800187 | MG799915 | MG799984 | – | MG800061 | – | MG800259 |
| T. heuretes (WAM T91918) | MN101151 | MN104643 | MN104652 | – | MN104658 | MN104650 | MN101153 |
| T. luculentus (WAM T141133) | MN101147 | – | – | – | – | – | – |
| T. meridionalis (WAM T147625) | MT280814 | MT280897 | MT280969 | – | MT281039 | MT281115 | MT281188 |
| T. vancouveri (WAM T16804) | – | MN104644 | MN104653 | – | – | – | – |
| T. sp. ‘MYG012’ (WAM T96062) | KJ745412 | KY214182 | KY241235 | KY241288 | KY241251 | KY241266 | MG800221 |
| T. sp. ‘MYG053’ (WAM T96326) | MG800159 | MG799883 | MG799949 | MG800286 | MG800021 | MG800098 | MG800222 |
| T. sp. ‘MYG358’ (WAM T78529) | KJ745277 | MG799875 | MG799936 | MG800274 | MG800007 | MG800084 | MG800205 |
| T. sp. ‘MYG412’ (WAM T116018) | MG800164 | MG799895 | MG799961 | MG800297 | MG800034 | MG800111 | MG800189 |
| Teyloides Main, 1985 [1] | |||||||
| T. bakeri (SAM NN29525) | MG800144 | MG799861 | – | MG800265 | MG799991 | MG800068 | MG800190 |
| Troglodiplura Main, 1969 [4] | |||||||
| T. beirutpakbarai (AM KS30823) | – | – | MT656259 | – | – | – | – |
| T. beirutpakbarai (SAM NN30808) | OQ922002 | – | OQ925874 | – | OQ925886 | OQ925878 | OQ924314 |
| T. beirutpakbarai (SAM NN30809) | OQ922003 | – | OQ925875 | OQ924320 | OQ925887 | OQ925880 | OQ924315 |
| T. beirutpakbarai (SAM NN30810) | OQ922004 | – | OQ925876 | OQ924321 | OQ925888 | OQ925879 | OQ924316 |
| DIPLURIDAE Linothele Karsch, 1879 [1] | |||||||
| L. sp. indet. (#ZSMA20170069) | – | MG273475 | MG273475 | MG273544 | MG273579 | MG273624 | – |
| MICROSTIGMATIDAE Ixamatus Simon, 1887 [1] | |||||||
| I. sp. indet. (#ARAMY002102) | KY017807 | – | KY015981 | KY018321 | DQ639831 | KY017212 | JQ358747 |
| Kiama Main & Mascord, 1969 [1] | |||||||
| K. lachrymoides (#ARAMY002094) | – | – | KY015982 | KY018322 | DQ639796 | DQ639884 | JQ358748 |
| Xamiatus Raven, 1981 [2] | |||||||
| X. rubrifrons (QMB S111351) | MT656265 | – | MT656260 | MT656268 | MT656262 | MT656264 | – |
| X. sp. indet. (QMB S111284) | – | – | – | MT656267 | MT656261 | MT656263 | – |
| PYCNOTHELIDAE Acanthogonatus Karsch, 1880 [1] | |||||||
| A. campanae (#ARAMYCAS539) | KY017803 | – | KY015977 | – | DQ639843 | KY017208 | JQ358720 |
| Stanwellia Rainbow & Pulleine, 1918 [4] | |||||||
| S. nebulosa (SAM NN28449) | MG800186 | MG799858 | MG799983 | – | MG800060 | MG800138 | MG800258 |
| S. sp. indet. (QMB S111338) | MT280818 | – | MT281031 | – | MT281096 | MT281178 | MT281251 |
| S. sp. indet. (QMB S111388) | – | – | MT281030 | – | MT281095 | MT281177 | – |
| S. sp. indet. (QMB S111428) | MT280817 | – | MT281029 | MT281252 | MT281094 | MT281176 | MT281250 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marsh, J.R.; Milner, S.J.; Shaw, M.; Stempel, A.J.; Harvey, M.S.; Rix, M.G. A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae). Insects 2023, 14, 449. https://doi.org/10.3390/insects14050449
Marsh JR, Milner SJ, Shaw M, Stempel AJ, Harvey MS, Rix MG. A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae). Insects. 2023; 14(5):449. https://doi.org/10.3390/insects14050449
Chicago/Turabian StyleMarsh, Jessica R., Steven J. Milner, Matthew Shaw, Andrew J. Stempel, Mark S. Harvey, and Michael G. Rix. 2023. "A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae)" Insects 14, no. 5: 449. https://doi.org/10.3390/insects14050449
APA StyleMarsh, J. R., Milner, S. J., Shaw, M., Stempel, A. J., Harvey, M. S., & Rix, M. G. (2023). A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae). Insects, 14(5), 449. https://doi.org/10.3390/insects14050449