

A Comparative Analysis of RNAi Trigger Uptake and Distribution in Mosquito Vectors of Disease
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing and Maintenance
2.2. Cell Culture
2.3. RNAi Trigger Synthesis and Labeling
2.4. RNAi Trigger Exposure of Adult-Stage Mosquitoes
2.5. RNAi Trigger Exposure of Larval-Stage Mosquitoes
2.6. Imaging of Live and Fixed Mosquito Tissues
2.7. Northern Blot Analyses
3. Results
3.1. Biodistribution of dsRNA in Ae. aegypti, An. gambiae, and Cx. pipiens
3.1.1. Tracking dsRNA following Injection in Adults and Larvae
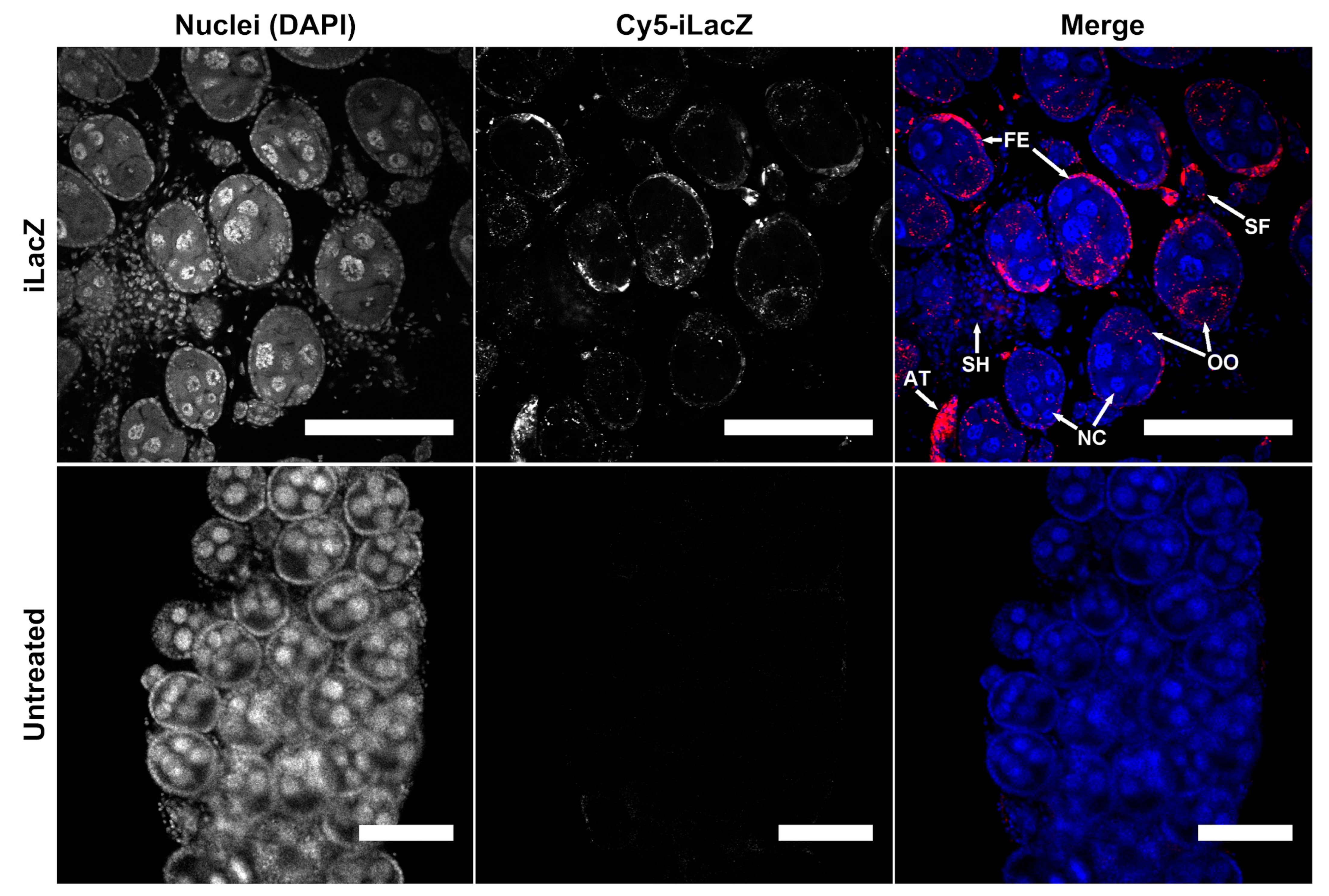
3.1.2. Ovaries Appear to Be a Primary Destination of dsRNA
3.1.3. Assessing dsRNA Uptake by Topical and Per Os Exposure Routes
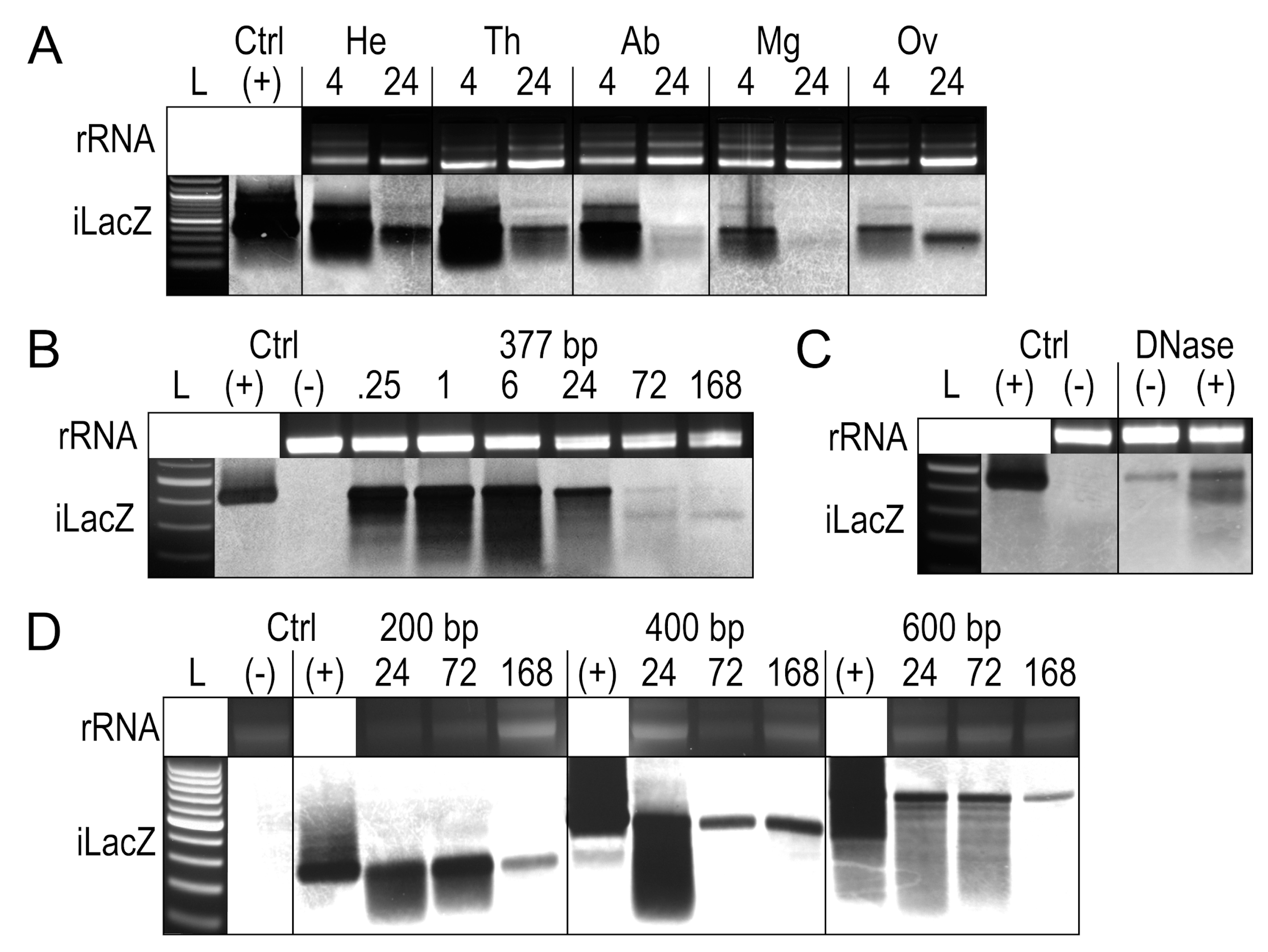
3.2. Assessing the Persistence and Integrity of dsRNA in Mosquito Tissues and Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, C.; Mueller, S.; Goto, A.; Barbier, V.; Paro, S.; Bonnay, F.; Dostert, C.; Troxler, L.; Hetru, C.; Meignin, C.; et al. Broad RNA Interference-Mediated Antiviral Immunity and Virus-Specific Inducible Responses in Drosophila. J. Immunol. 2013, 190, 650–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.-C.; Tassetto, M.; van Rij, R.P.; Goic, B.; Gausson, V.; Berry, B.; Jacquier, C.; Antoniewski, C.; Andino, R. Antiviral Immunity in Drosophila Requires Systemic RNA Interference Spread. Nature 2009, 458, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Nayak, A.; Tassetto, M.; Kunitomi, M.; Andino, R. RNA Interference-Mediated Intrinsic Antiviral Immunity in Invertebrates. Curr. Top. Microbiol. Immunol. 2013, 371, 183–200. [Google Scholar]
- Mongelli, V.; Saleh, M.-C. Bugs Are Not to Be Silenced: Small RNA Pathways and Antiviral Responses in Insects. Annu Rev. Virol. 2016, 3, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.E.; Blair, C.D. Arbovirus-Mosquito Interactions: RNAi Pathway. Curr. Opin. Virol. 2015, 15, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Blair, C.D. Mosquito RNAi Is the Major Innate Immune Pathway Controlling Arbovirus Infection and Transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.E.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue Virus Type 2 Infections of Aedes aegypti Are Modulated by the Mosquito’s RNA Interference Pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [Green Version]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-Derived DNA Drives Mosquito Vector Tolerance to Arboviral Infection. Nat. Commun. 2016, 7, 12410. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Lee, Y.S.; Harris, D.; Nakahara, K.; Carthew, R.W. The RNAi Pathway Initiated by Dicer-2 in Drosophila. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoa, N.T.; Keene, K.M.; Olson, K.E.; Zheng, L. Characterization of RNA Interference in an Anopheles Gambiae Cell Line. Insect Biochem. Mol. Biol. 2003, 33, 949–957. [Google Scholar] [CrossRef]
- Sontheimer, E.J. Assembly and Function of RNA Silencing Complexes. Nat. Rev. Mol. Cell Biol. 2005, 6, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.-C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The RNA Silencing Endonuclease Argonaute 2 Mediates Specific Antiviral Immunity in Drosophila Melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airs, P.M.; Bartholomay, L.C. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects 2017, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Wiltshire, R.M.; Duman-Scheel, M. Advances in Oral RNAi for Disease Vector Mosquito Research and Control. Curr. Opin. Insect Sci. 2020, 40, 18–23. [Google Scholar] [CrossRef]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.-L.; Barthel, A.; et al. RNA Interference in Lepidoptera: An Overview of Successful and Unsuccessful Studies and Implications for Experimental Design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the Elements of Successful Insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Peng, Y.; Pu, J.; Fu, W.; Wang, J.; Han, Z. Variation in RNAi Efficacy among Insect Species Is Attributable to dsRNA Degradation In Vivo. Insect Biochem. Mol. Biol. 2016, 77, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.-C.; van Rij, R.P.; Hekele, A.; Gillis, A.; Foley, E.; O’Farrell, P.H.; Andino, R. The Endocytic Pathway Mediates Cell Entry of dsRNA to Induce RNAi Silencing. Nat. Cell Biol. 2006, 8, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating Immune Cells Mediate a Systemic RNAi-Based Adaptive Antiviral Response in Drosophila. Cell 2017, 169, 314–325.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesbrecht, D.; Heschuk, D.; Wiens, I.; Boguski, D.; LaChance, P.; Whyard, S. RNA Interference Is Enhanced by Knockdown of Double-Stranded RNases in the Yellow Fever Mosquito Aedes aegypti. Insects 2020, 11, 327. [Google Scholar] [CrossRef]
- Boisson, B.; Jacques, J.C.; Choumet, V.; Martin, E.; Xu, J.; Vernick, K.; Bourgouin, C. Gene Silencing in Mosquito Salivary Glands by RNAi. FEBS Lett. 2006, 580, 1988–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telang, A.; Rechel, J.A.; Brandt, J.R.; Donnell, D.M. Analysis of Ovary-Specific Genes in Relation to Egg Maturation and Female Nutritional Condition in the Mosquitoes Georgecraigius atropalpus and Aedes aegypti (Diptera: Culicidae). J. Insect Physiol. 2013, 59, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sijen, T.; Fleenor, J.; Simmer, F.; Thijssen, K.L.; Parrish, S.; Timmons, L.; Plasterk, R.H.; Fire, A. On the Role of RNA Amplification in dsRNA-Triggered Gene Silencing. Cell 2001, 107, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. Elegans Requires the Putative Transmembrane Protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.H.; Quarles, K.A.; Yang, Y.; Tanguy, M.; Frézal, L.; Smith, S.A.; Sharma, P.P.; Cordaux, R.; Gilbert, C.; Giraud, I.; et al. Pan-Arthropod Analysis Reveals Somatic piRNAs as an Ancestral Defence against Transposable Elements. Nat. Ecol. Evol. 2018, 2, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Pinzón, N.; Bertrand, S.; Subirana, L.; Busseau, I.; Escrivá, H.; Seitz, H. Functional Lability of RNA-Dependent RNA Polymerases in Animals. PLoS Genet. 2019, 15, e1007915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlikow, M.; Goic, B.; Saleh, M.-C. RNAi and Antiviral Defense in Drosophila: Setting up a Systemic Immune Response. Dev. Comp. Immunol. 2014, 42, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.-S.; Kim, K.; Palli, S.R. Double-Stranded RNA in Exosomes: Potential Systemic RNA Interference Pathway in the Colorado Potato Beetle, Leptinotarsa Decemlineata. J. Asia. Pac. Entomol. 2020, 23, 1160–1164. [Google Scholar] [CrossRef]
- Mingels, L.; Wynant, N.; Santos, D.; Peeters, P.; Gansemans, Y.; Billen, J.; Van Nieuwerburgh, F.; Vanden Broeck, J. Extracellular Vesicles Spread the RNA Interference Signal of Tribolium Castaneum TcA Cells. Insect Biochem. Mol. Biol. 2020, 122, 103377. [Google Scholar] [CrossRef]
- Zamanian, M.; Fraser, L.M.; Agbedanu, P.N.; Harischandra, H.; Moorhead, A.R.; Day, T.A.; Bartholomay, L.C.; Kimber, M.J. Release of Small RNA-Containing Exosome-like Vesicles from the Human Filarial Parasite Brugia Malayi. PLoS Negl. Trop. Dis. 2015, 9, e0004069. [Google Scholar] [CrossRef] [Green Version]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes Secreted by Nematode Parasites Transfer Small RNAs to Mammalian Cells and Modulate Innate Immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, N.; Tojo, K.; Hatakeyama, M. A Survey of the Effectiveness of Non-Cell Autonomous RNAi throughout Development in the Sawfly, Athalia rosae (Hymenoptera). J. Insect Physiol. 2013, 59, 400–407. [Google Scholar] [CrossRef]
- Airs, P.M.; Kudrna, K.E.; Bartholomay, L.C. Impact of Sugar Composition on Meal Distribution, Longevity, and Insecticide Toxicity in Aedes aegypti. Acta Trop. 2019, 191, 221–227. [Google Scholar] [CrossRef]
- Singh, A.D.; Wong, S.; Ryan, C.P.; Whyard, S. Oral Delivery of Double-Stranded RNA in Larvae of the Yellow Fever Mosquito, Aedes aegypti: Implications for Pest Mosquito Control. J. Insect Sci. 2013, 13, 69. [Google Scholar] [CrossRef] [Green Version]
- Bryant, W.B.; Michel, K. Blood Feeding Induces Hemocyte Proliferation and Activation in the African Malaria Mosquito, Anopheles Gambiae Giles. J. Exp. Biol. 2014, 217, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, J.M.; Bratu, D.P. Drosophila Melanogaster Oogenesis: An Overview. Methods Mol. Biol. 2015, 1328, 1–20. [Google Scholar] [PubMed]
- Chapman, R.F. The Insects: Structure and Function; Cambridge University Press: Cambridge, UK, 2013; ISBN 9780521113892. [Google Scholar]
- Livshits, M.A.; Amosova, O.A. Lyubchenko YuL Flexibility Difference between Double-Stranded RNA and DNA as Revealed by Gel Electrophoresis. J. Biomol. Struct. Dyn. 1990, 7, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Poirier, E.Z.; Goic, B.; Tomé-Poderti, L.; Frangeul, L.; Boussier, J.; Gausson, V.; Blanc, H.; Vallet, T.; Loyd, H.; Levi, L.I.; et al. Dicer-2-Dependent Generation of Viral DNA from Defective Genomes of RNA Viruses Modulates Antiviral Immunity in Insects. Cell Host Microbe 2018, 23, 353–365.e8. [Google Scholar] [CrossRef] [Green Version]
- Brackney, D.E.; Scott, J.C.; Sagawa, F.; Woodward, J.E.; Miller, N.A.; Schilkey, F.D.; Mudge, J.; Wilusz, J.; Olson, K.E.; Blair, C.D.; et al. C6/36 Aedes Albopictus Cells Have a Dysfunctional Antiviral RNA Interference Response. PLoS Negl. Trop. Dis. 2010, 4, e856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, D.; Aradhya, R.; Ashoka, D.; Inamdar, M. Macromolecular Uptake in Drosophila Pericardial Cells Requires Rudhira Function. Exp. Cell Res. 2008, 314, 1804–1810. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Schmidt, S.L.; Christensen, B.M. Hemocyte-Mediated Phagocytosis and Melanization in the Mosquito Armigeres Subalbatus Following Immune Challenge by Bacteria. Cell Tissue Res. 2003, 313, 117–127. [Google Scholar] [CrossRef]
- Cantera, R.; Technau, G.M. Glial Cells Phagocytose Neuronal Debris during the Metamorphosis of the Central Nervous System in Drosophila Melanogaster. Dev. Genes Evol. 1996, 206, 277–280. [Google Scholar] [CrossRef]
- Kurant, E.; Axelrod, S.; Leaman, D.; Gaul, U. Six-Microns-under Acts Upstream of Draper in the Glial Phagocytosis of Apoptotic Neurons. Cell 2008, 133, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.; Deri, P. Cell Death in Ovarian Chambers of Drosophila Melanogaster. J. Embryol. Exp. Morphol. 1976, 35, 521–533. [Google Scholar] [CrossRef]
- Anderson, W.A.; Spielman, A. Permeability of the Ovarian Follicle of Aedes aegypti Mosquitoes. J. Cell Biol. 1971, 50, 201–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulvila, J.; Parikka, M.; Kleino, A.; Sormunen, R.; Ezekowitz, R.A.; Kocks, C.; Rämet, M. Double-Stranded RNA Is Internalized by Scavenger Receptor-Mediated Endocytosis in Drosophila S2 Cells. J. Biol. Chem. 2006, 281, 14370–14375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, J.J.E.; Korolchuk, V.I.; Robinson, I.M.; O’Kane, C.J. A Phagocytic Route for Uptake of Double-Stranded RNA in RNAi. PLoS ONE 2011, 6, e19087. [Google Scholar] [CrossRef]
- League, G.P.; Hillyer, J.F. Functional Integration of the Circulatory, Immune, and Respiratory Systems in Mosquito Larvae: Pathogen Killing in the Hemocyte-Rich Tracheal Tufts. BMC Biol. 2016, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Cai, Z.; Song, J.; Wu, Z.; Zhou, S. dsRNA Uptake and Persistence Account for Tissue-Dependent Susceptibility to RNA Interference in the Migratory Locust, Locusta Migratoria. Insect Mol. Biol. 2014, 23, 175–184. [Google Scholar] [CrossRef]
- Kolliopoulou, A.; Swevers, L. Functional Analysis of the RNAi Response in Ovary-Derived Silkmoth Bm5 Cells. Insect Biochem. Mol. Biol. 2013, 43, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Went, D.F. Oocyte Maturation without Follicular Epithelium Alters Egg Shape in a Dipteran Insect. J. Exp. Zool. 1978, 205, 149–155. [Google Scholar] [CrossRef]
- Raikhel, A.S.; Lea, A.O. Control of Follicular Epithelium Development and Vitelline Envelope Formation in the Mosquito; Role of Juvenile Hormone and 20-Hydroxyecdysone. Tissue Cell 1991, 23, 577–591. [Google Scholar] [CrossRef]
- Cheung, H.K.; Serano, T.L.; Cohen, R.S. Evidence for a Highly Selective RNA Transport System and Its Role in Establishing the Dorsoventral Axis of the Drosophila Egg. Development 1992, 114, 653–661. [Google Scholar] [CrossRef]
- Cooley, L.; Verheyen, E.; Ayers, K. Chickadee Encodes a Profilin Required for Intercellular Cytoplasm Transport during Drosophila Oogenesis. Cell 1992, 69, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Lasko, P. mRNA Localization and Translational Control in Drosophila Oogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a012294. [Google Scholar] [CrossRef] [Green Version]
- Coy, M.R.; Sanscrainte, N.D.; Chalaire, K.C.; Inberg, A.; Maayan, I.; Glick, E.; Paldi, N.; Becnel, J.J. Gene Silencing in Adult Aedes aegypti Mosquitoes through Oral Delivery of Double-Stranded RNA: Aedes aegypti RNAi. J. Appl. Entomol. 2012, 136, 741–748. [Google Scholar] [CrossRef]
- Van Ekert, E.; Powell, C.A.; Shatters, R.G., Jr.; Borovsky, D. Control of Larval and Egg Development in Aedes aegypti with RNA Interference against Juvenile Hormone Acid Methyl Transferase. J. Insect Physiol. 2014, 70, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Hixson, B.; Bing, X.-L.; Yang, X.; Bonfini, A.; Nagy, P.; Buchon, N. A Transcriptomic Atlas of Aedes aegypti Reveals Detailed Functional Organization of Major Body Parts and Gut Regional Specializations in Sugar-Fed and Blood-Fed Adult Females. Elife 2022, 11, e76132. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, J.; Li, D.; Cooper, A.M.W.; Silver, K.; Li, T.; Liu, X.; Ma, E.; Zhu, K.Y.; Zhang, J. A Double-Stranded RNA Degrading Enzyme Reduces the Efficiency of Oral RNA Interference in Migratory Locust. Insect Biochem. Mol. Biol. 2017, 86, 68–80. [Google Scholar] [CrossRef]
- Roy, S.G.; Hansen, I.A.; Raikhel, A.S. Effect of Insulin and 20-Hydroxyecdysone in the Fat Body of the Yellow Fever Mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2007, 37, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandon, M.C.; Pennington, J.E.; Isoe, J.; Zamora, J.; Schillinger, A.-S.; Miesfeld, R.L. TOR Signaling Is Required for Amino Acid Stimulation of Early Trypsin Protein Synthesis in the Midgut of Aedes aegypti Mosquitoes. Insect Biochem. Mol. Biol. 2008, 38, 916–922. [Google Scholar] [CrossRef] [Green Version]
- Spit, J.; Philips, A.; Wynant, N.; Santos, D.; Plaetinck, G.; Vanden Broeck, J. Knockdown of Nuclease Activity in the Gut Enhances RNAi Efficiency in the Colorado Potato Beetle, Leptinotarsa Decemlineata, but Not in the Desert Locust, Schistocerca Gregaria. Insect Biochem. Mol. Biol. 2017, 81, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Mueller, C.R.; Fuchs, J.F.; McElroy, K.; Wessely, V.; Higgs, S.; Christensen, B.M. Evaluation of the Function of a Type I Peritrophic Matrix as a Physical Barrier for Midgut Epithelium Invasion by Mosquito-Borne Pathogens in Aedes aegypti. Vector Borne Zoonotic Dis. 2008, 8, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Raphemot, R.; Estévez-Lao, T.Y.; Rouhier, M.F.; Piermarini, P.M.; Denton, J.S.; Hillyer, J.F. Molecular and Functional Characterization of Anopheles Gambiae Inward Rectifier Potassium (Kir1) Channels: A Novel Role in Egg Production. Insect Biochem. Mol. Biol. 2014, 51, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, C.; Denlinger, D.L. Catalase and Superoxide Dismutase-2 Enhance Survival and Protect Ovaries during Overwintering Diapause in the Mosquito Culex Pipiens. J. Insect Physiol. 2011, 57, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.C.; Brackney, D.E.; Campbell, C.L.; Bondu-Hawkins, V.; Hjelle, B.; Ebel, G.D.; Olson, K.E.; Blair, C.D. Comparison of Dengue Virus Type 2-Specific Small RNAs from RNA Interference-Competent and -Incompetent Mosquito Cells. PLoS Negl. Trop. Dis. 2010, 4, e848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miesen, P.; Girardi, E.; van Rij, R.P. Distinct Sets of PIWI Proteins Produce Arbovirus and Transposon-Derived piRNAs in Aedes aegypti Mosquito Cells. Nucleic Acids Res. 2015, 43, 6545–6556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodovar, N.; Bronkhorst, A.W.; van Cleef, K.W.R.; Miesen, P.; Blanc, H.; van Rij, R.P.; Saleh, M.-C. Arbovirus-Derived piRNAs Exhibit a Ping-Pong Signature in Mosquito Cells. PLoS ONE 2012, 7, e30861. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | HPE | Related Figures | Tissue-Affiliated Hemocytes | Intracellular Signal Locations | Extracellular Signal Locations | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HD | LE | TH | HC-FB | TA-MT | TA-OV | PC | FB | VC | FO | AT | DV-CA | FG | MG | HG | RC | CT | IS | |||
| Peritoneal exposure by intrathoracic injection—adult female | ||||||||||||||||||||
| Ae. aegypti | 24–120 | S1 | +++ | +++ | ++ | +++ | +++ | ++ | ++++ | - | ++ | +++ | - | - | - | - | ++ | ++ | - | ++ |
| An. gambiae | 24–120 | S2 | ++ | - | + | ++ | +++ | +++ | ++++ | - | - | +++ | - | - | - | + | + | + | - | ++ |
| Cx. pipiens | 24–120 | S3 | + | nd | - | ++ | ++ | +++ | ++++ | - | - | +++ | - | - | + | + | ++ | + | - | +++ |
| Peritoneal exposure by intrathoracic injection 24 h post bloodmeal—adult female | ||||||||||||||||||||
| Ae. aegypti | 1–24 | S4 | - | - | + | + | - | + | ++++ | - | - | +++ | - | - | - | - | - | - | - | - |
| An. gambiae | 1–24 | S5 | +++ | ++ | - | +++ | + | ++ | ++++ | - | - | +++ | - | - | - | - | - | - | - | + |
| Cx. pipiens | 1–24 | S6 | + | + | + | +++ | + | + | ++++ | + | - | +++ | - | - | - | - | - | - | - | ++ |
| Peritoneal exposure by intrathoracic injection—4th instar larvae | ||||||||||||||||||||
| Ae. aegypti | 24 | S7 | - | na | - | - | - | na | ++ | - | - | na | - | - | - | + | - | - | - | na |
| An. gambiae | 24 | S8 | + | na | - | + | - | na | ++ | - | - | na | - | - | - | + | - | - | - | na |
| Cx. pipiens | 24 | S9 | - | na | - | + | - | na | ++ | - | - | na | - | - | - | + | - | - | - | na |
| Topical exposure—adult female | ||||||||||||||||||||
| Ae. aegypti | 72 | S10 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ++++ | na |
| An. gambiae | 72 | S11 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ++++ | na |
| Cx. pipiens | 72 | S12 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ++++ | na |
| Per os exposure—adult female | ||||||||||||||||||||
| Ae. aegypti | 24–120 | S13 | - | - | - | - | - | - | + | - | - | - | - | +++ | +++ | +++ | +++ | +++ | - | na |
| Cx. pipiens | 24–120 | S14 | ++ | ++ | ++ | - | + | ++ | ++ | - | - | + | ++ | +++ | +++ | +++ | +++ | +++ | - | na |
| Soaking exposure—1st instar larvae | ||||||||||||||||||||
| Ae. aegypti | 24–72 | S15 | na | na | - | - | - | na | - | - | - | na | na | na | - | +++ | +++ | +++ | - | na |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Airs, P.M.; Kudrna, K.E.; Lubinski, B.; Phanse, Y.; Bartholomay, L.C. A Comparative Analysis of RNAi Trigger Uptake and Distribution in Mosquito Vectors of Disease. Insects 2023, 14, 556. https://doi.org/10.3390/insects14060556
Airs PM, Kudrna KE, Lubinski B, Phanse Y, Bartholomay LC. A Comparative Analysis of RNAi Trigger Uptake and Distribution in Mosquito Vectors of Disease. Insects. 2023; 14(6):556. https://doi.org/10.3390/insects14060556
Chicago/Turabian StyleAirs, Paul M., Katherine E. Kudrna, Bailey Lubinski, Yashdeep Phanse, and Lyric C. Bartholomay. 2023. "A Comparative Analysis of RNAi Trigger Uptake and Distribution in Mosquito Vectors of Disease" Insects 14, no. 6: 556. https://doi.org/10.3390/insects14060556
APA StyleAirs, P. M., Kudrna, K. E., Lubinski, B., Phanse, Y., & Bartholomay, L. C. (2023). A Comparative Analysis of RNAi Trigger Uptake and Distribution in Mosquito Vectors of Disease. Insects, 14(6), 556. https://doi.org/10.3390/insects14060556