

Expression Pattern and Ligand Binding Characteristics Analysis of Chemosensory Protein SnitCSP2 from Sirex nitobei



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preservation
2.2. RNA Isolation and PCR Amplification
2.3. Tissue Expression Pattern and Sequencing Analysis
2.4. Homology Modeling and Molecular Docking
2.5. Molecular Dynamics Simulation of the SnitCSP2-Ligand Complexes
3. Results
3.1. Identification and Analysis of SnitCSP2
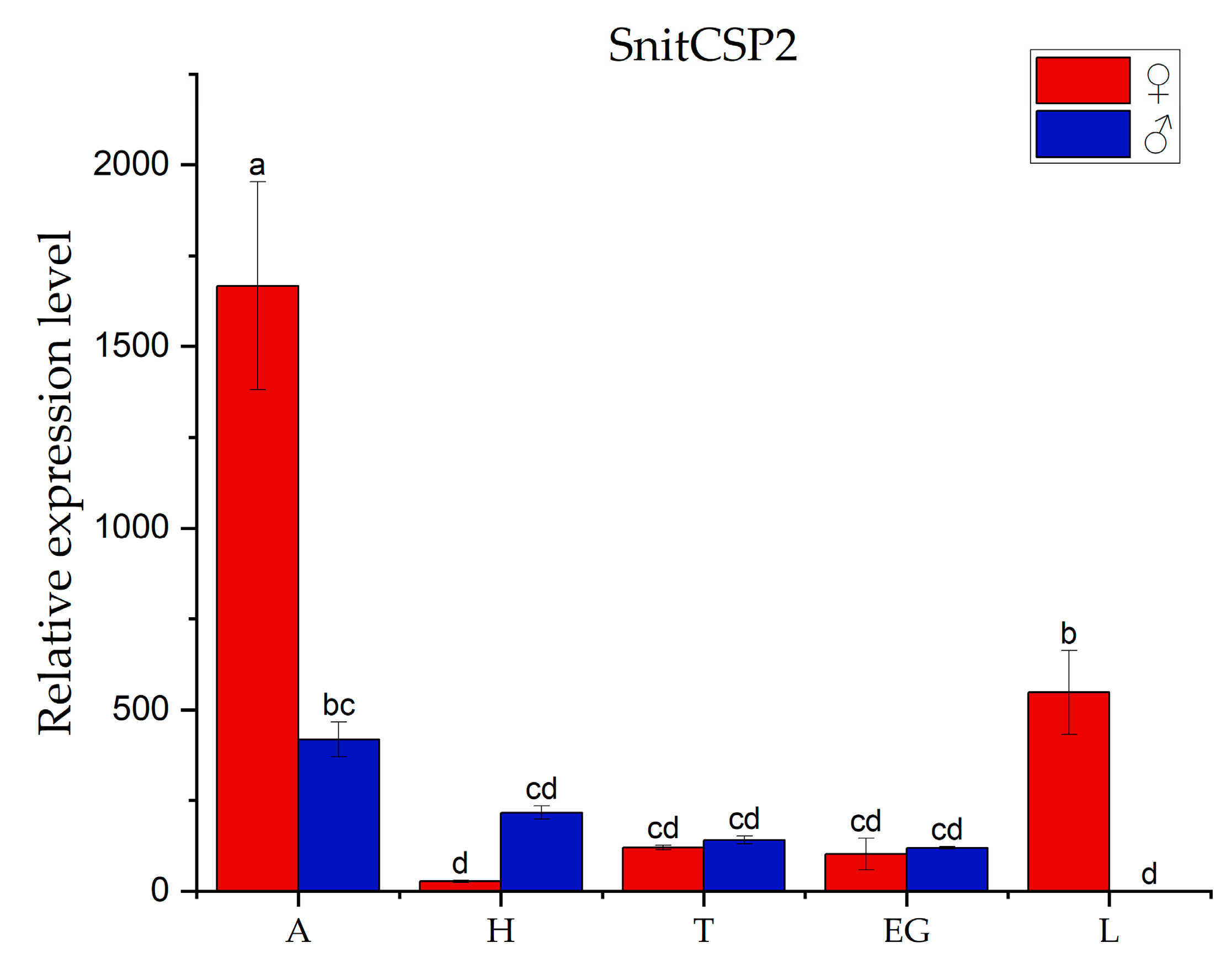
3.2. Tissues Expression Pattern Analysis
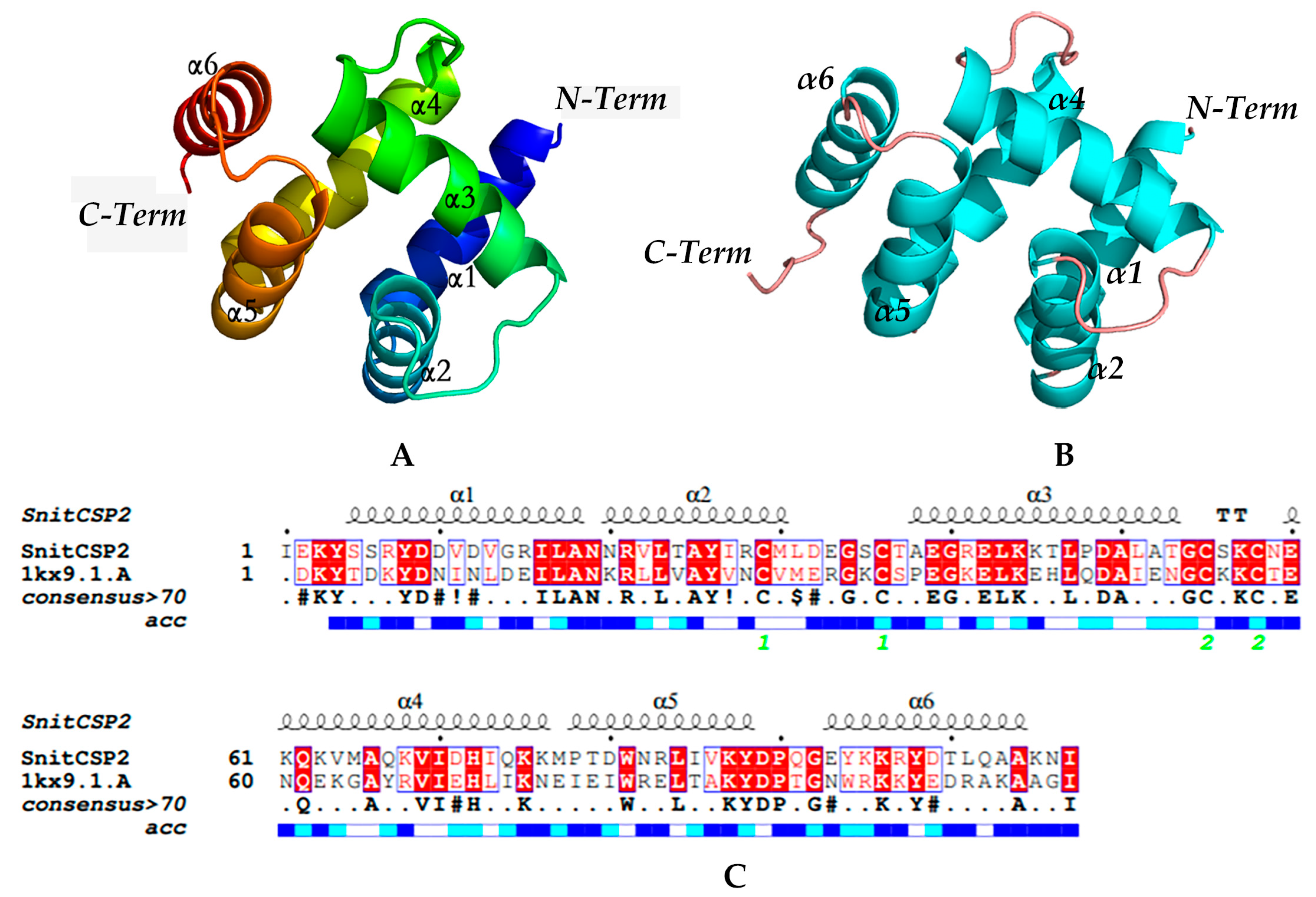
3.3. Modeling and Model Evaluation
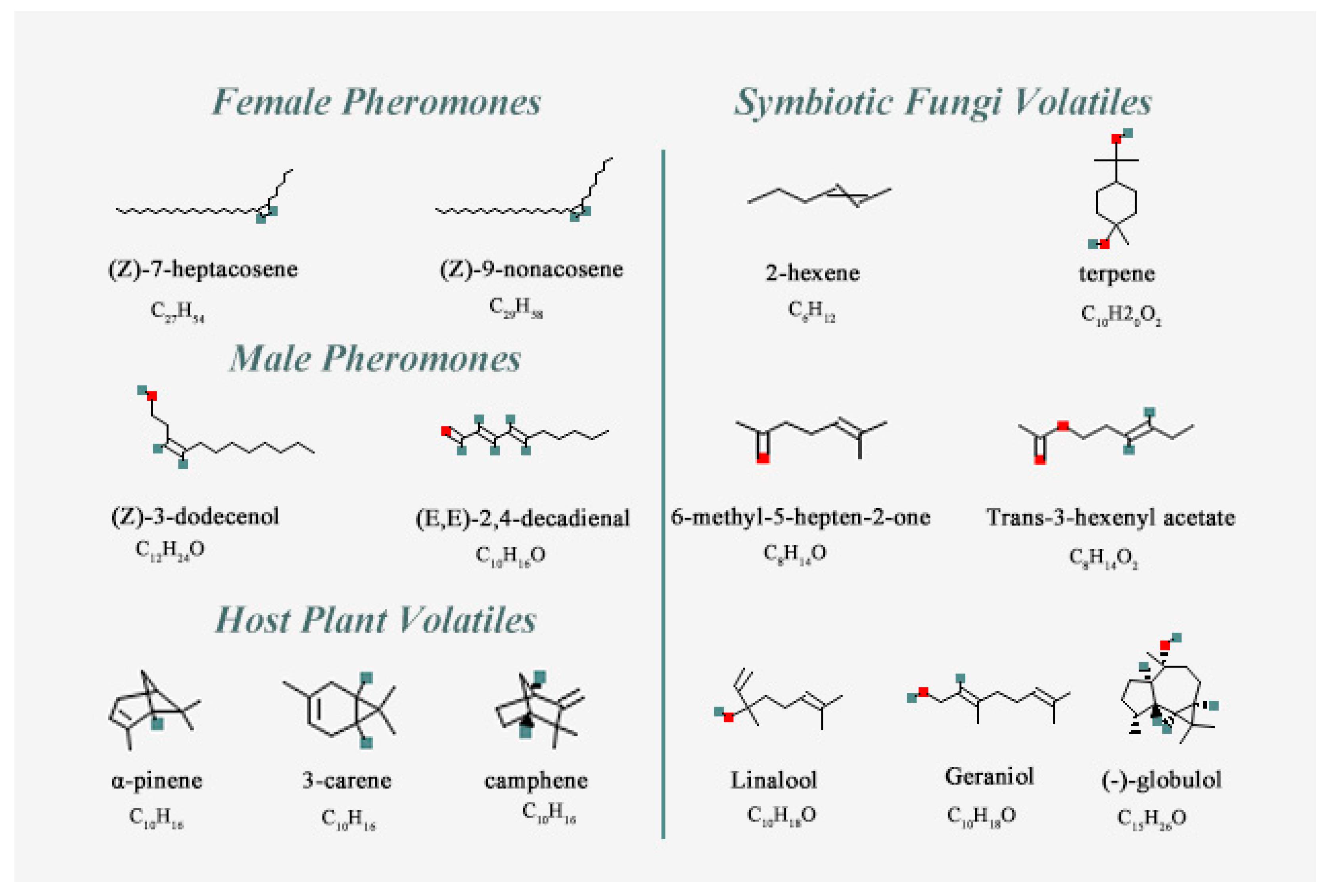
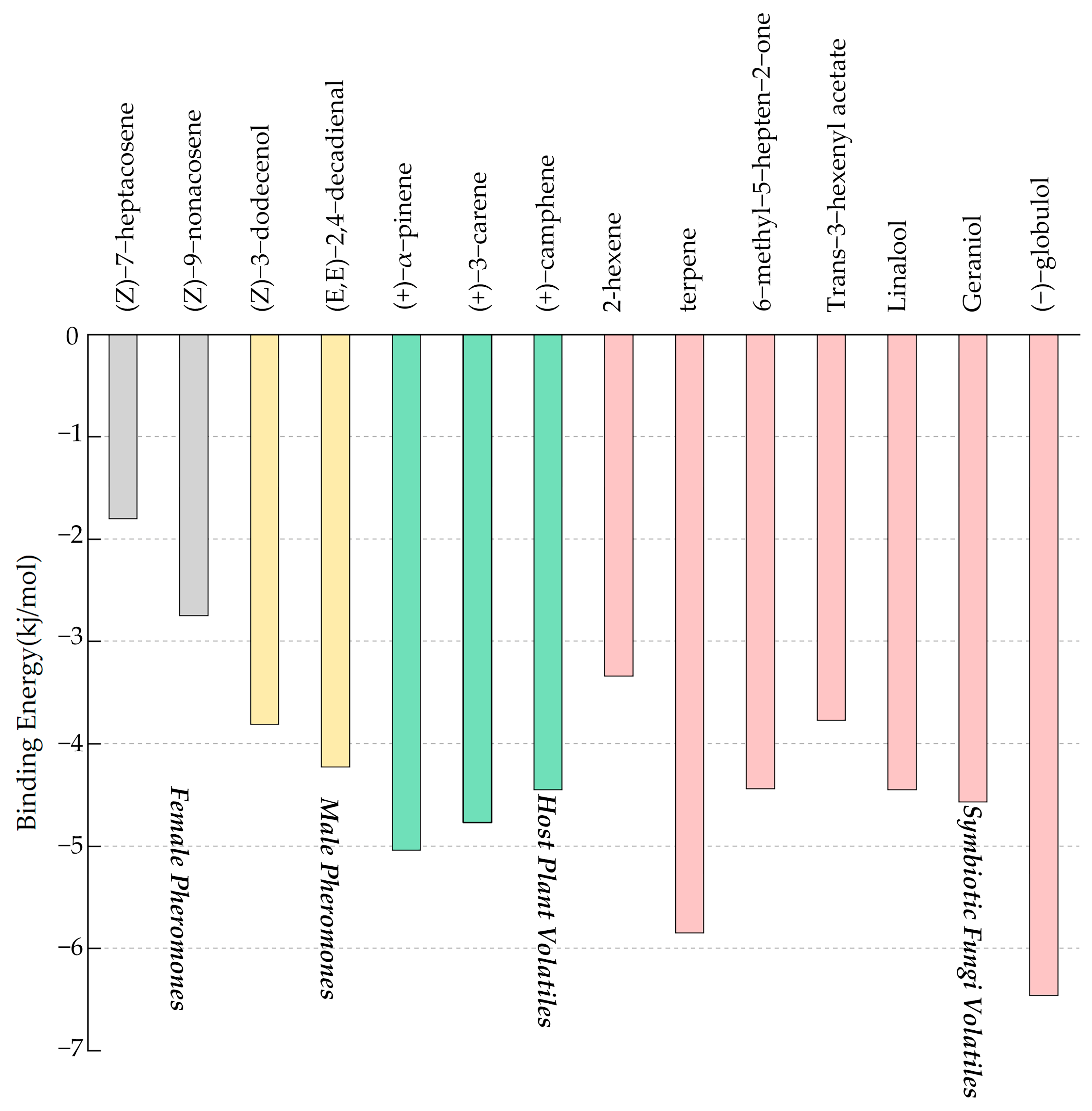
3.4. Binding Affinities for SnitCSP2 with Ligands and Molecular Docking
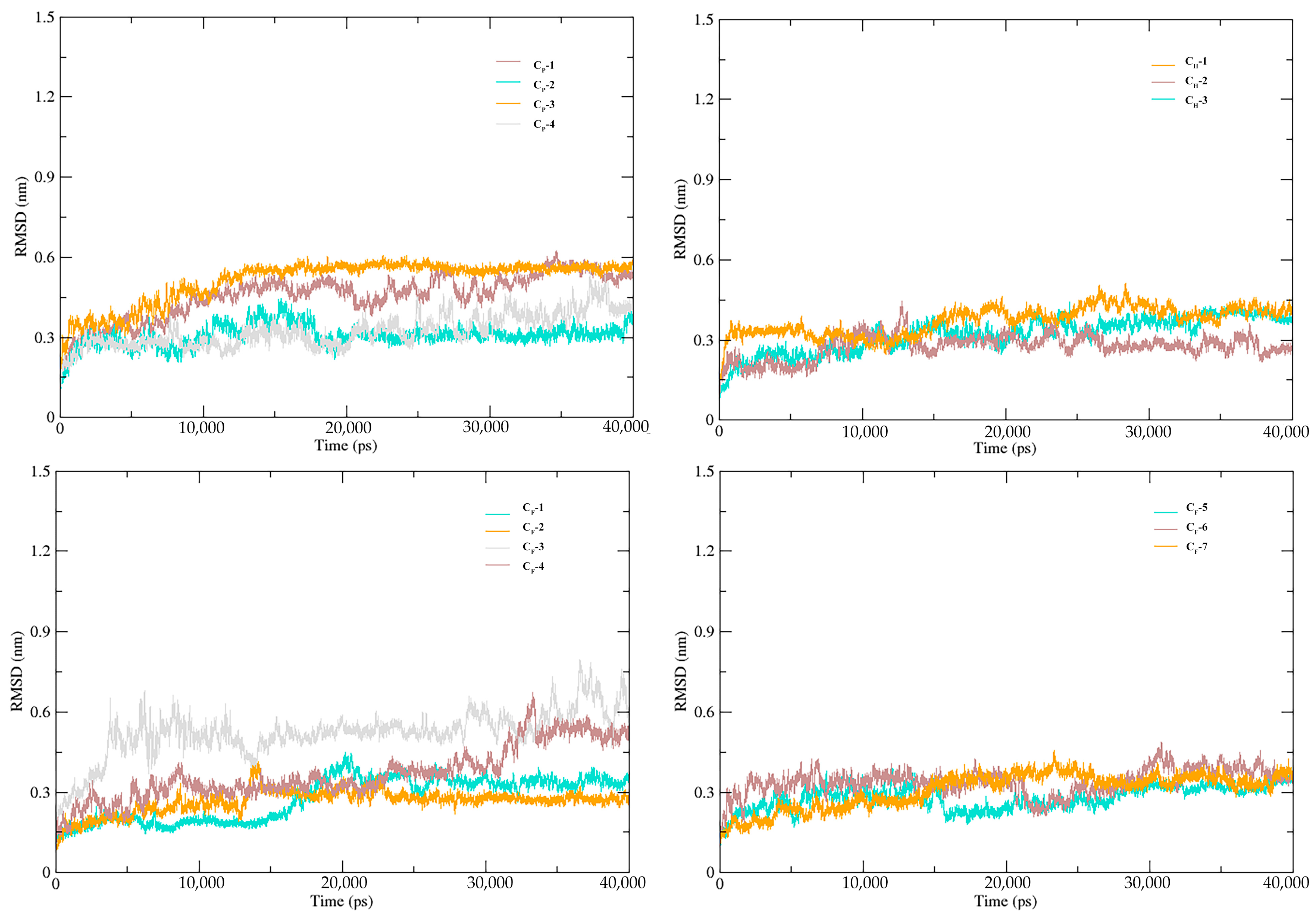
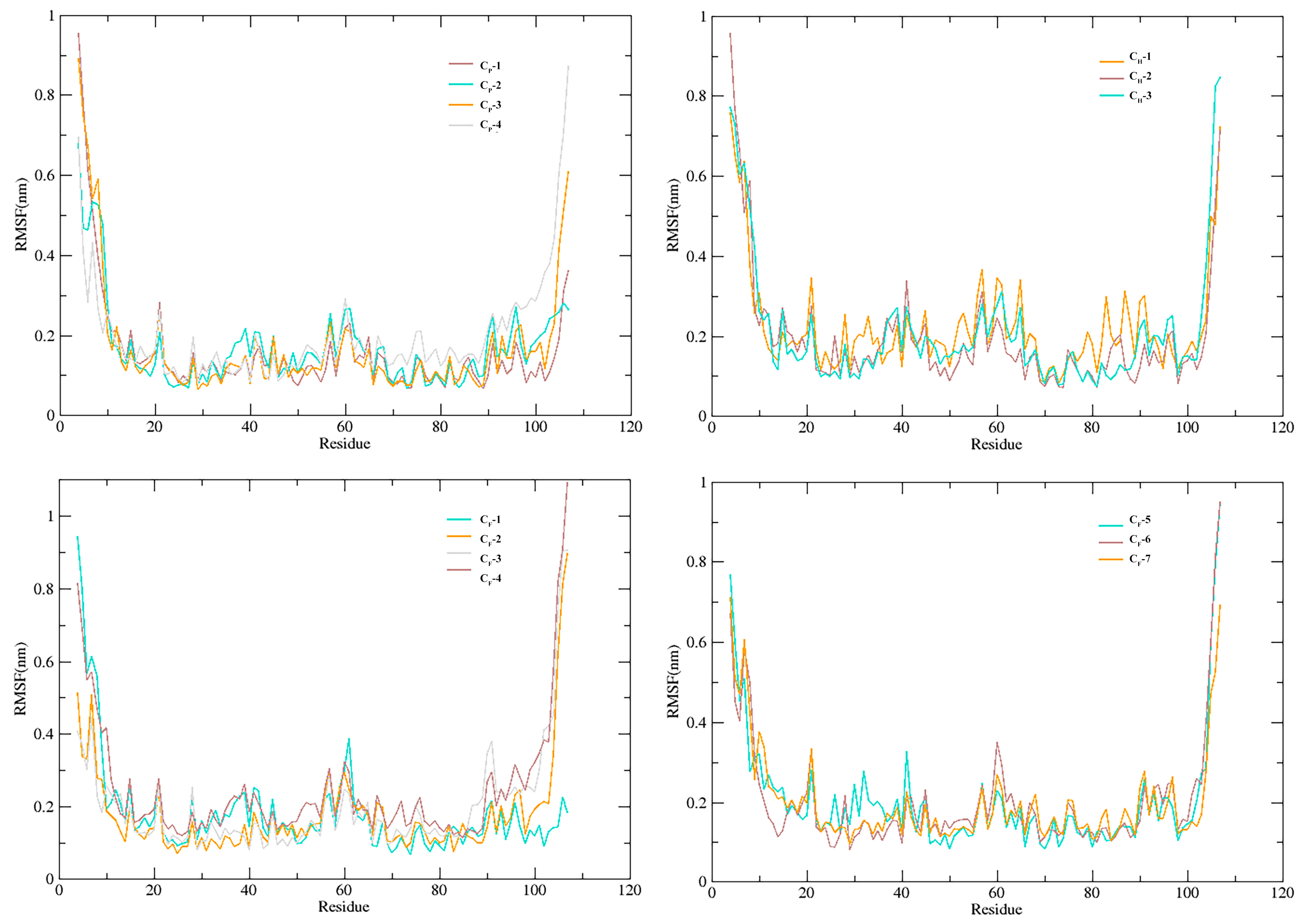
3.5. Stability of SnitCSP2-Ligand Complexes in MD Simulation
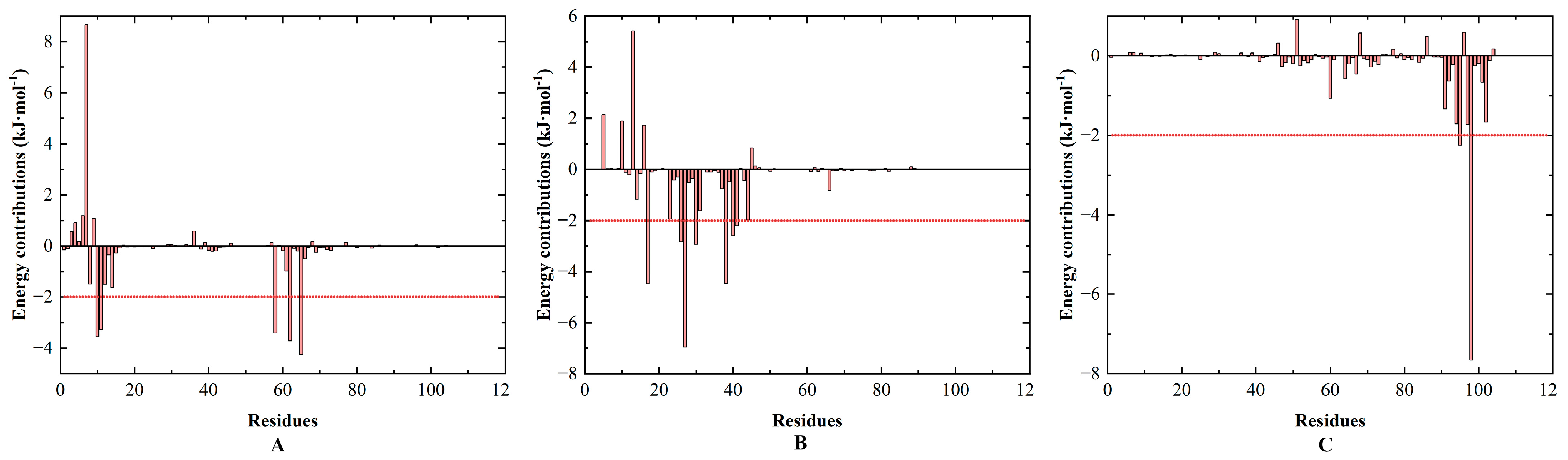
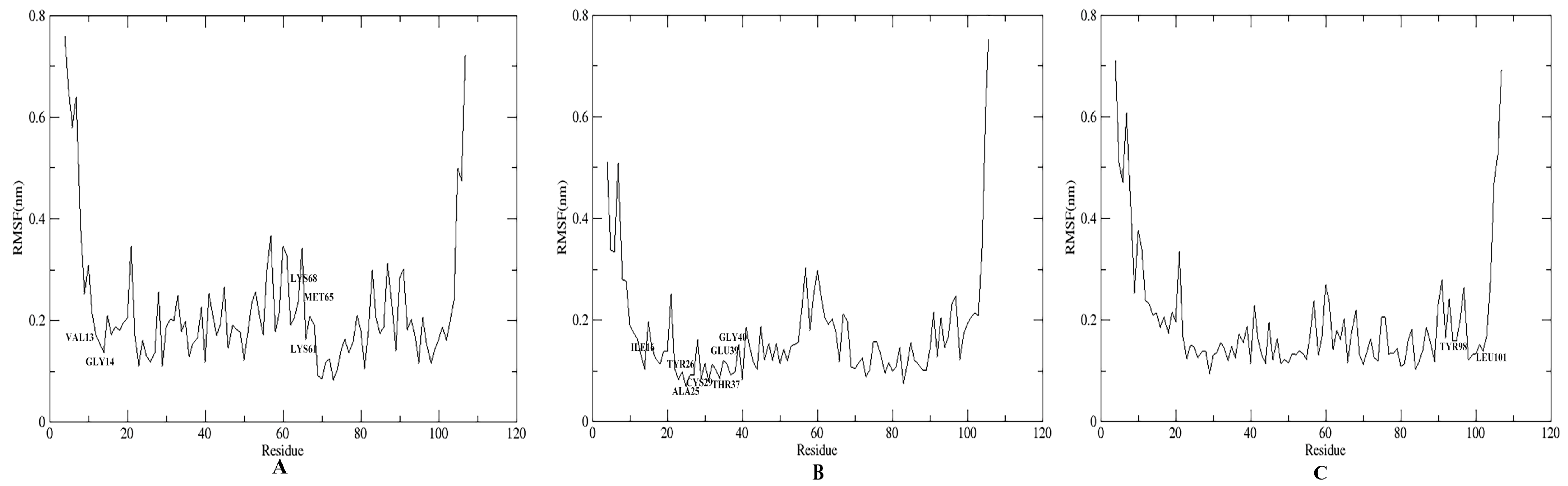
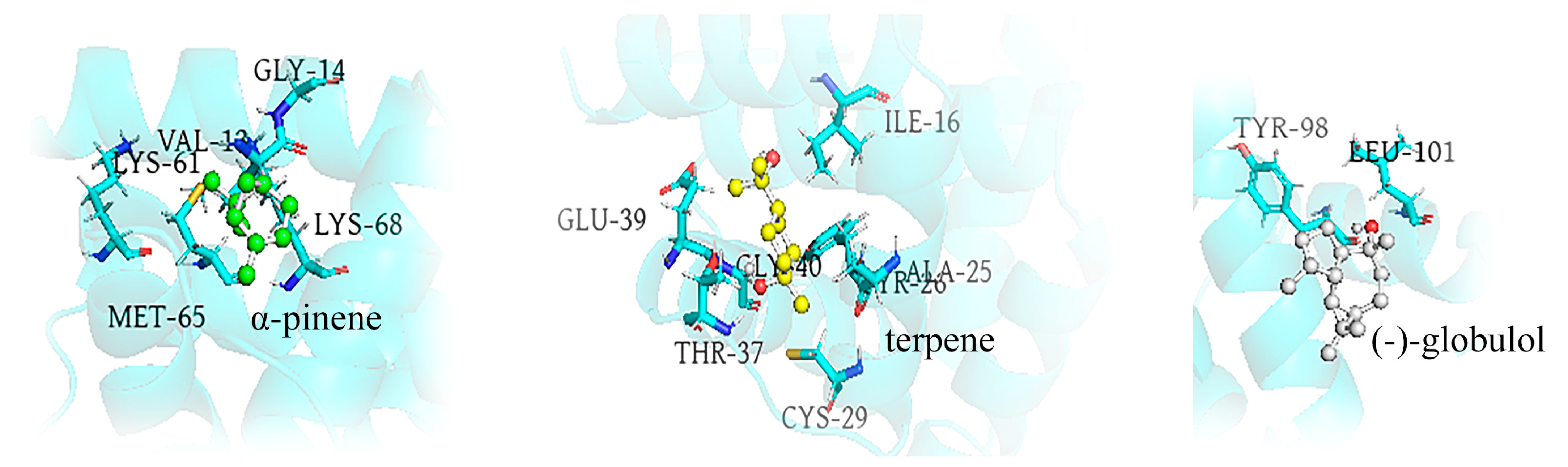
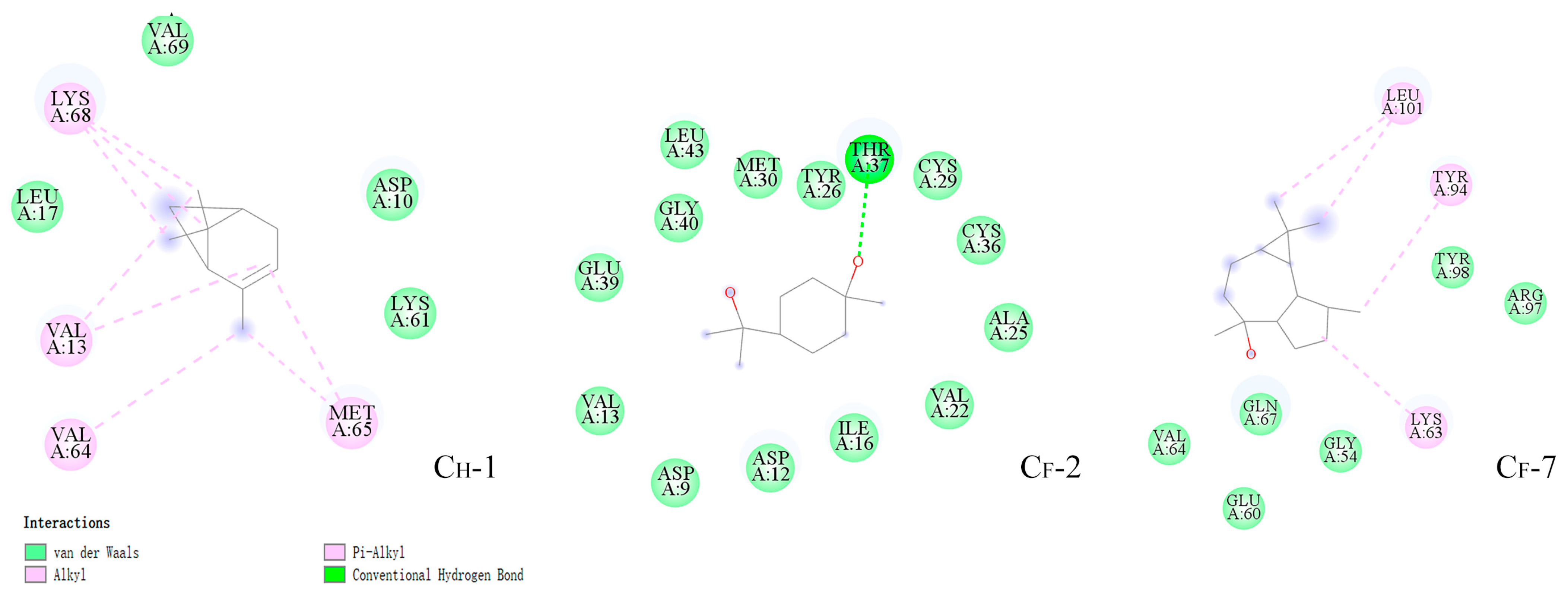
3.6. Energy Calculation and Binding Modes Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Liberles, S.D. Aversion and Attraction through Olfaction. Curr. Biol. 2015, 25, R120–R129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, D. Insect Olfaction: Deciphering System for Chemical Messages: Receptor Cells beneath a Porous Cuticle Are Highly Sensitive and Specific to Odor Molecules. Science 1969, 163, 1031–1037. [Google Scholar] [CrossRef]
- Clark, J.T.; Ray, A. Olfactory Mechanisms for Discovery of Odorants to Reduce Insect-Host Contact. J. Chem. Ecol. 2016, 42, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Getchell, T.; Margolis, F.; Getchell, M. Perireceptor and Receptor Events in Vertebrate Olfaction. Prog. Neurobiol. 1984, 23, 317–345. [Google Scholar] [CrossRef] [PubMed]
- Borrero-Echeverry, F.; Bengtsson, M.; Nakamuta, K.; Witzgall, P. Plant Odor and Sex Pheromone Are Integral Elements of Specific Mate Recognition in an Insect Herbivore. Evolution 2018, 72, 2225–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crava, C.M.; Sassù, F.; Tait, G.; Becher, P.G.; Anfora, G. Functional Transcriptome Analyses of Drosophila Suzukii Antennae Reveal Mating-Dependent Olfaction Plasticity in Females. Insect Biochem. Mol. Biol. 2019, 105, 51–59. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond Chemoreception: Diverse Tasks of Soluble Olfactory Proteins in Insects: Soluble Olfactory Proteins in Insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Falchetto, M.; Ciossani, G.; Scolari, F.; Di Cosimo, A.; Nenci, S.; Field, L.M.; Mattevi, A.; Zhou, J.-J.; Gasperi, G.; Forneris, F. Structural and Biochemical Evaluation of Ceratitis capitata Odorant-binding Protein 22 Affinity for Odorants Involved in Intersex Communication. Insect Mol. Biol. 2019, 28, 431–443. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, L.; Qiu, Y.-L.; Li, H.-L. Research progress of olfactory binding proteins in insects. Acta Entomologica Sinica. 2021, 64, 523–535. [Google Scholar] [CrossRef]
- Campanacci, V.; Lartigue, A.; Hällberg, B.M.; Jones, T.A.; Giudici-Orticoni, M.-T.; Tegoni, M.; Cambillau, C. Moth Chemosensory Protein Exhibits Drastic Conformational Changes and Cooperativity on Ligand Binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5069–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dani, F.R.; Michelucci, E.; Francese, S.; Mastrobuoni, G.; Cappellozza, S.; La Marca, G.; Niccolini, A.; Felicioli, A.; Moneti, G.; Pelosi, P. Odorant-Binding Proteins and Chemosensory Proteins in Pheromone Detection and Release in the Silkmoth Bombyx Mori. Chem. Senses 2011, 36, 335–344. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Kan, Y.; Antoniw, J.; Pickett, J.A.; Field, L.M. Genome and EST Analyses and Expression of a Gene Family with Putative Functions in Insect Chemoreception. Chem. Senses 2006, 31, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Briand, L.; Swasdipan, N.; Nespoulous, C.; Bézirard, V.; Blon, F.; Huet, J.-C.; Ebert, P.; Pernollet, J.-C. Characterization of a Chemosensory Protein (ASP3c) from Honeybee (Apis mellifera L.) as a Brood Pheromone Carrier: Brood Pheromone Binding by Bee Chemosensory Protein. Eur. J. Biochem. 2002, 269, 4586–4596. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.P.; Hekmat-Scafe, D.S.; Gaines, P.; Carlson, J.R. Putative Drosophila Pheromone-Binding Proteins Expressed in a Subregion of the Olfactory System. J. Biol. Chem. 1994, 269, 16340–16347. [Google Scholar] [CrossRef] [PubMed]
- Wanner, K.W.; Willis, L.G.; Theilmann, D.A.; Isman, M.B.; Feng, Q.; Plettner, E. Analysis of the Insect OS-D-Like Gene Family. J. Chem. Ecol. 2004, 30, 889–911. [Google Scholar] [CrossRef]
- Pikielny, C.W.; Hasan, G.; Rouyer, F.; Rosbash, M. Members of a Family of Drosophila Putative Odorant-Binding Proteins Are Expressed in Different Subsets of Olfactory Hairs. Neuron 1994, 12, 35–49. [Google Scholar] [CrossRef]
- Maleszka, R.; Stange, G. Molecular Cloning, by a Novel Approach, of a CDNA Encoding a Putative Olfactory Protein in the Labial Palps of the Moth Cactoblastis Cactorum. Gene 1997, 202, 39–43. [Google Scholar] [CrossRef]
- Nomura Kitabayashi, A.; Arai, T.; Kubo, T.; Natori, S. Molecular Cloning of cDNA for P10, a Novel Protein That Increases in the Regenerating Legs of Periplaneta Americana (American cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Biessmann, H.; Walter, M.F.; Dimitratos, S.; Woods, D. Isolation of cDNA Clones Encoding Putative Odourant Binding Proteins from the Antennae of the Malaria-Transmitting Mosquito, Anopheles gambiae. Insect Mol. Biol. 2002, 11, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Biessmann, H.; Nguyen, Q.K.; Le, D.; Walter, M.F. Microarray-Based Survey of a Subset of Putative Olfactory Genes in the Mosquito Anopheles gambiae: Gene Expression in Antennae of Anopheles gambiae. Insect Mol. Biol. 2005, 14, 575–589. [Google Scholar] [CrossRef]
- Angeli, S.; Ceron, F.; Scaloni, A.; Monti, M.; Monteforti, G.; Minnocci, A.; Petacchi, R.; Pelosi, P. Purification, Structural Characterization, Cloning and Immunocytochemical Localization of Chemoreception Proteins from Schistocerca gregaria. Eur. J. Biochem. 1999, 262, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Jacquin-Joly, E.; Vogt, R.G.; François, M.C.; Nagnan-Le Meillour, P. Functional and Expression Pattern Analysis of Chemosensory Proteins Expressed in Antennae and Pheromonal Gland of Mamestra Brassicae. Chem. Senses 2001, 26, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gu, T.; Chen, C.; Huang, K.; Tian, S.; Zhao, X.; Hao, D. cDNA Cloning, Sequence Analysis and Expression Profie of a Chemosensory Protoin from the Clostera Restitura (Lepidoptera: Notodontidae). Sci. Silvae Sin. 2018, 67–75. [Google Scholar] [CrossRef]
- Iovinella, I.; Bozza, F.; Caputo, B.; della Torre, A.; Pelosi, P. Ligand-Binding Study of Anopheles gambiae Chemosensory Proteins. Chem. Senses 2013, 38, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.-H.; Wang, S.-Y.; Zhang, X.-Y.; Ji, P.; Liu, J.-T.; Wang, G.-R.; Wu, M.; Guo, Y.-Y.; Zhou, J.-J.; Zhang, Y.-J. Functional Characterizations of Chemosensory Proteins of the Alfalfa Plant Bug Adelphocoris Lineolatus Indicate Their Involvement in Host Recognition. PLoS ONE 2012, 7, e42871. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Ye, Z.-F.; Yang, K.; Dong, S.-L. Antenna-Predominant and Male-Biased CSP19 of Sesamia inferens Is Able to Bind the Female Sex Pheromones and Host Plant Volatiles. Gene 2014, 536, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.R.; Huang, X.Y.; Zhou, S.Z.; Wu, J.; Zhang, P. Economic Sawfly Fauna of China. (Hymenoptera, Symphyta); Tianze Eldonejo: Beijing, China, 1991; pp. 37–42. [Google Scholar]
- Wang, M.; Wang, L.; Fu, N.; Gao, C.; Ao, T.; Ren, L.; Luo, Y. Comparison of Wing, Ovipositor, and Cornus Morphologies between Sirex noctilio and Sirex Nitobei Using Geometric Morphometrics. Insects 2020, 11, 84. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Xu, Q.; Liu, Y.; Zhao, J.; Shi, J. Predicting the Potential Geographic Distribution of Sirex Nitobei in China under Climate Change Using Maximum Entropy Model. Forests 2021, 12, 151. [Google Scholar] [CrossRef]
- Guo, B.; Hao, E.; Qiao, H.; Wang, J.; Wu, W.; Zhou, J.; Lu, P. Antennal Transcriptome Analysis of Olfactory Genes and Characterizations of Odorant Binding Proteins in Two Woodwasps, Sirex noctilio and Sirex Nitobei (Hymenoptera: Siricidae). BMC Genom. 2021, 22, 172. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein Structure Homology Modeling Using SWISS-MODEL Workspace. Nat. Protoc. 2009, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Luthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of Protein Models with Three-Dimensional Profiles. Nature 1992, 356, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Moss, D.S.; Thornton, J.M. Main-Chain Bond Lengths and Bond Angles in Protein Structures. J. Mol. Biol. 1993, 231, 1049–1067. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of Protein Structures: Patterns of Nonbonded Atomic Interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Sousa da Silva, A.W.; Vranken, W.F. ACPYPE—AnteChamber PYthon Parser InterfacE. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Yuan, X.; Li, J.; Lin, S.; Yang, B.; Chen, C.; Zhao, J.; Zheng, W.; Liao, H.; Yang, Z.; et al. Assessing the Performance of the G_mmpbsa Tools to Simulate the Inhibition of Oseltamivir to Influenza Virus Neuraminidase by Molecular Mechanics Poisson–Boltzmann Surface Area Methods. J. Chin. Chem. Soc. 2020, 67, 46–53. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Specht, C.A.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Chitin Synthases Are Required for Survival, Fecundity and Egg Hatch in the Red Flour Beetle, Tribolium Castaneum. Insect Biochem. Mol. Biol. 2008, 38, 959–962. [Google Scholar] [CrossRef]
- Li, C.; Sun, K.; Li, D.; Liu, D. Functional Characterization of Chemosensory Protein AmalCSP5 from Apple Buprestid Beetle, Agrilus Mali (Coleoptera: Buprestidae). J. Econ. Entomol. 2021, 114, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, Y.-J.; Su, H.-H.; Gao, X.-W.; Guo, Y.-Y. Identification and Expression Pattern of Putative Odorant-Binding Proteins and Chemosensory Proteins in Antennae of the Microplitis Mediator (Hymenoptera: Braconidae). Chem. Senses 2009, 34, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, F.; Zhang, X.; Zhang, S.; Guo, S.; Zhu, G.; Liu, Q.; Li, M. Transcriptome and Expression Patterns of Chemosensory Genes in Antennae of the Parasitoid Wasp Chouioia Cunea. PLoS ONE 2016, 11, e0148159. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Wang, S.-N.; Li, K.-M.; Liu, J.-T.; Zheng, Y.; Shan, S.; Yang, Y.-Q.; Li, R.-J.; Zhang, Y.-J.; Guo, Y.-Y. Identification of Odorant Binding Proteins and Chemosensory Proteins in Microplitis Mediator as Well as Functional Characterization of Chemosensory Protein 3. PLoS ONE 2017, 12, e0180775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waris, M.I.; Younas, A.; Ameen, A.; Rasool, F.; Wang, M.-Q. Expression Profiles and Biochemical Analysis of Chemosensory Protein 3 from Nilaparvata lugens (Hemiptera: Delphacidae). J. Chem. Ecol. 2020, 46, 363–377. [Google Scholar] [CrossRef]
- Zhou, X.-H.; Ban, L.-P.; Iovinella, I.; Zhao, L.-J.; Gao, Q.; Felicioli, A.; Sagona, S.; Pieraccini, G.; Pelosi, P.; Zhang, L.; et al. Diversity, Abundance, and Sex-Specific Expression of Chemosensory Proteins in the Reproductive Organs of the Locust Locusta migratoria manilensis. Biol. Chem. 2013, 394, 43–54. [Google Scholar] [CrossRef]
- Nomura, A.; Kawasaki, K.; Kubo, T.; Natori, S. Purification and Localization of P10, a Novel Protein That Increases in Nymphal Regenerating Legs of Periplaneta americana (American Cockroach). Int. J. Dev. Biol. 1992, 36, 391–398. [Google Scholar] [PubMed]
- Zhang, Y.-N.; Jin, J.-Y.; Jin, R.; Xia, Y.-H.; Zhou, J.-J.; Deng, J.-Y.; Dong, S.-L. Differential Expression Patterns in Chemosensory and Non-Chemosensory Tissues of Putative Chemosensory Genes Identified by Transcriptome Analysis of Insect Pest the Purple Stem Borer Sesamia inferens (Walker). PLoS ONE 2013, 8, e69715. [Google Scholar] [CrossRef] [Green Version]
- González-González, A.; Rubio-Meléndez, M.E.; Ballesteros, G.I.; Ramírez, C.C.; Palma-Millanao, R. Sex- and Tissue-Specific Expression of Odorant-Binding Proteins and Chemosensory Proteins in Adults of the Scarab Beetle Hylamorpha Elegans (Burmeister) (Coleoptera: Scarabaeidae). PeerJ 2019, 7, e7054. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Cui, S.; Tian, Z.; Zhang, Y.; Chen, G.; Gao, X.; Tian, Z.; Chen, H.; Guo, J.; Zhou, Z. OcomCSP12, a Chemosensory Protein Expressed Specifically by Ovary, Mediates Reproduction in Ophraella Communa (Coleoptera: Chrysomelidae). Front. Physiol. 2019, 10, 1290. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.-L.; Luo, Z.-X.; Li, J.-L.; Cai, X.-M.; Bian, L.; Xiu, C.-L.; Li, Z.-Q.; Chen, Z.-M.; Zhang, L.-W. Identification of Cytochrome P450, Odorant-Binding Protein, and Chemosensory Protein Genes Involved in Type II Sex Pheromone Biosynthesis and Transportation in the Tea Pest, Scopula subpunctaria. Pestic. Biochem. Physiol. 2020, 169, 104650. [Google Scholar] [CrossRef] [PubMed]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory Proteins in the Honey Bee: Insights from the Annotated Genome, Comparative Analyses and Expressional Profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Maleszka, J.; Forêt, S.; Saint, R.B.; Maleszka, R. RNAi-Induced Phenotypes Suggest a Novel Role for a Chemosensory Protein CSP5 in the Development of Embryonic Integument in the Honeybee (Apis mellifera). Dev. Genes Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef] [PubMed]
- PD, K.J.; Kempraj, V.; Aurade, R.M.; Kumar Roy, T.; KS, S.; Verghese, A. Computational Reverse Chemical Ecology: Virtual Screening and Predicting Behaviorally Active Semiochemicals for Bactrocera dorsalis. BMC Genom. 2014, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-L.; Pan, Y.-F.; Ma, Y.-F.; Wang, J.; He, M.; He, P. Binding Affinity Characterization of an Antennae-Enriched Chemosensory Protein from the White-Backed Planthopper, Sogatella furcifera (Horváth), with Host Plant Volatiles. Pestic. Biochem. Physiol. 2018, 152, 1–7. [Google Scholar] [CrossRef]
- Tian, Z.; Liu, J.; Zhang, Y. Structural Insights into Cydia Pomonella Pheromone Binding Protein 2 Mediated Prediction of Potentially Active Semiochemicals. Sci. Rep. 2016, 6, 22336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venthur, H.; Machuca, J.; Godoy, R.; Palma-Millanao, R.; Zhou, J.; Larama, G.; Bardehle, L.; Quiroz, A.; Ceballos, R.; Mutis, A. Structural Investigation of Selective Binding Dynamics for the Pheromone-binding Protein 1 of the Grapevine Moth, Lobesia botrana. Arch. Insect Biochem. Physiol. 2019, 101, e21557. [Google Scholar] [CrossRef] [PubMed]
- Zylstra, K.E.; Dodds, K.J.; Francese, J.A.; Mastro, V. Sirex noctilio in North America: The Effect of Stem-Injection Timing on the Attractiveness and Suitability of Trap Trees. Agric. For. Entomol. 2010, 12, 243–250. [Google Scholar] [CrossRef]
- Wang, M.; Bao, M.; Ao, T.; Ren, L.; Luo, Y. Population Distribution Patterns and Ecological Niches of Two Sirex Species Damaging Pinus sylvestris Var. Mongolica. Chin. J. Appl. Entomol. 2017, 54, 924–932. [Google Scholar]
- Xu, Q.; Sun, X.-T.; Lu, P.-F.; Luo, Y.-Q.; Shi, J. Volatile Profiles of Three Tree Species in the Northeastern China and Associated Effects on Sirex noctilio Activity. J. Plant Interact. 2019, 14, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Bashford, R. The Development of Static Trapping Systems to Monitor for Wood-Boring Insects in Forestry Plantations. Aust. For. 2008, 71, 236–241. [Google Scholar] [CrossRef]
- Talbot, P.H.B. The Sirex-Amylostereum-Pinus Association. Annu. Rev. Phytopathol. 1977, 15, 41–54. [Google Scholar] [CrossRef]
- Coutts, M. The Mechanism of Pathogenicity of Sirex noctilio on Pinus Radiata I. Effects of the Symbiotic Fungus Amylostereum Sp. (Thelophoraceae). Aust. Jnl. Bio. Sci. 1969, 22, 915. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.M.; Bodart, J.; McEwen, C.; Gruner, D.S. Adaptations for Symbiont-Mediated External Digestion in Sirex noctilio (Hymenoptera: Siricidae). Ann. Entomol. Soc. Am. 2014, 107, 453–460. [Google Scholar] [CrossRef]
- Wang, L.-X.; Ren, L.-L.; Liu, X.-B.; Shi, J.; Wang, J.-Z.; Luo, Y.-Q. Effects of Endophytic Fungi in Mongolian Pine on the Selection Behavior of Woodwasp (Sirex noctilio) and the Growth of Its Fungal Symbiont. Pest. Manag. Sci. 2019, 75, 492–505. [Google Scholar] [CrossRef]
- Tomaselli, S.; Crescenzi, O.; Sanfelice, D.; Ab, E.; Wechselberger, R.; Angeli, S.; Scaloni, A.; Boelens, R.; Tancredi, T.; Pelosi, P.; et al. Solution Structure of a Chemosensory Protein from the Desert Locust Schistocerca gregaria. Biochemistry 2006, 45, 10606–10613. [Google Scholar] [CrossRef] [PubMed]
- Lartigue, A.; Campanacci, V.; Roussel, A.; Larsson, A.M.; Jones, T.A.; Tegoni, M.; Cambillau, C. X-ray Structure and Ligand Binding Study of a Moth Chemosensory Protein. J. Biol. Chem. 2002, 277, 32094–32098. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primers Sequence (5′→3′) | Annealing Temperature, °C | Segment Length, bp |
|---|---|---|---|
| SnitCSP2 | F: GTGACGACAGTTATGGCCATTG R: TGCAGCGAATATAGGCAGTG | 59.0 | 106 |
| β-Tubulin | F: CGTCGGTTCCGTTGATAAGTTG R: AGAATATCCCGACCGAGTGTTG | 59.0 | 122 |
| Chemical Name | PubChem CID No. | Molecular Formula | Complex Code | Chemical Name | PubChem CID No. | Molecular Formula | Complex Code |
|---|---|---|---|---|---|---|---|
| Female Pheromones | MalePheromones | ||||||
| (Z)-7-heptacosene | 56936088 | C27H54 | CP-1 | (Z)-3-dodecenol | 5364626 | C12H24O | CP-3 |
| (Z)-9-nonacosene | 14367299 | C29H58 | CP-2 | (E,E)-2,4-decadienal | 5283349 | C10H16O | CP-4 |
| Host Plant Volatiles | Symbiotic Fungi Volatiles | ||||||
| α-pinene | 6654 | C10H16 | CH-1 | 2-hexene | 19966 | C6H12 | CF-1 |
| 3-carene | 26049 | C10H16 | CH-2 | terpene | 6651 | C10H20O2 | CF-2 |
| camphene | 92221 | C10H16 | CH-3 | 6-methyl-5-hepten-2-one | 9862 | C8H14O | CF-3 |
| Trans-3-hexenyl acetate | 5352557 | C8H14O2 | CF-4 | ||||
| Linalool | 6549 | C10H18O | CF-5 | ||||
| Geraniol | 637566 | C10H18O | CF-6 | ||||
| (−)-globulol | 12304985 | C15H26O | CF-7 |
| Name | Molecular Formula | MW (ku) | pI | Arg + Lys | Asp + Glu | Aliphatic Index | Instability Index | GRAVY |
|---|---|---|---|---|---|---|---|---|
| SnitCSP2 | C625H1024N172O184S10 | 14.21 | 9.10 | 21 | 15 | 88.87 | 54.11 | −0.346 |
| The Binding Energy Components | CH-1 (kJ/mol) | CF-2 (kJ/mol) | CF-7 (kJ/mol) |
|---|---|---|---|
| ΔE VDW | −53.189 ± 0.154 | −99.712 ± 0.363 | −71.014 ± 0.342 |
| ΔE elec | −0.539 ± 0.002 | −9.687 ± 0.036 | −0.581 ± 0.003 |
| ΔG PB | 19.537 ± 0.056 | 68.209 ± 0.247 | 24.826 ± 0.119 |
| ΔG SA | −11.559 ± 0.034 | −16.699 ± 0.061 | −12.959 ± 0.062 |
| T ΔS S | −0.937 ± 0.003 | −7.443 ± 0.027 | −3.310 ± 0.016 |
| ΔG bind | −44.813 ± 0.189 | −50.446 ± 0.396 | −56.418 ± 0.368 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Hao, E.; Li, H.; Yang, X.; Lu, P.; Qiao, H. Expression Pattern and Ligand Binding Characteristics Analysis of Chemosensory Protein SnitCSP2 from Sirex nitobei. Insects 2023, 14, 583. https://doi.org/10.3390/insects14070583
Guo P, Hao E, Li H, Yang X, Lu P, Qiao H. Expression Pattern and Ligand Binding Characteristics Analysis of Chemosensory Protein SnitCSP2 from Sirex nitobei. Insects. 2023; 14(7):583. https://doi.org/10.3390/insects14070583
Chicago/Turabian StyleGuo, Pingping, Enhua Hao, Han Li, Xi Yang, Pengfei Lu, and Haili Qiao. 2023. "Expression Pattern and Ligand Binding Characteristics Analysis of Chemosensory Protein SnitCSP2 from Sirex nitobei" Insects 14, no. 7: 583. https://doi.org/10.3390/insects14070583
APA StyleGuo, P., Hao, E., Li, H., Yang, X., Lu, P., & Qiao, H. (2023). Expression Pattern and Ligand Binding Characteristics Analysis of Chemosensory Protein SnitCSP2 from Sirex nitobei. Insects, 14(7), 583. https://doi.org/10.3390/insects14070583