
Colonization Resistance of Symbionts in Their Insect Hosts

Abstract
:Simple Summary
Abstract
1. Introduction
2. Mechanisms of Colonization Resistance
2.1. Upregulation of the Expression of Host Immune-Related Genes
2.2. Symbiont-Mediated Insect Immune Priming
2.3. Production of Antimicrobial Substances
2.4. Competitive Exclusion of Pathogens
3. Balance of Host Immune Response
3.1. Degeneration of the Host Immune System
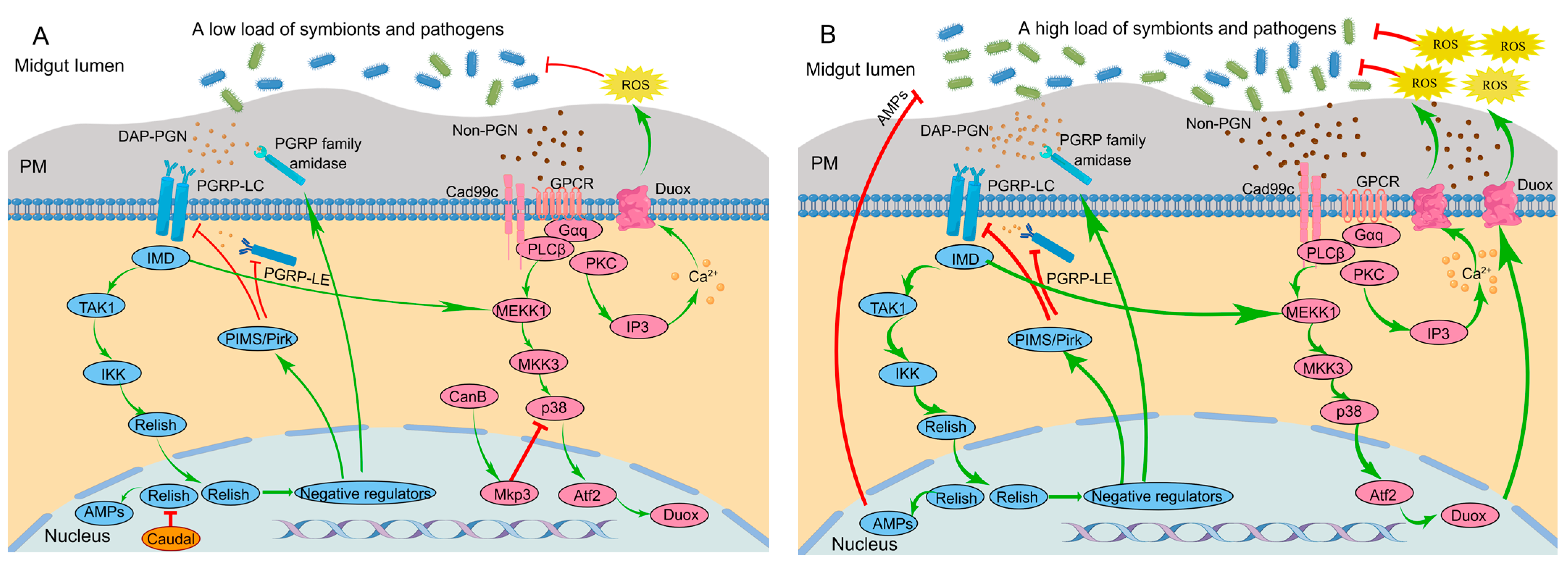
3.2. Fine-Tuned Regulation of Immune Responses by Negative Regulators
3.2.1. IMD-AMP Pathway
3.2.2. DUOX-ROS
3.3. Compartmentalization of Symbionts
4. Adaptation of Symbionts to Host Immune
5. Coevolution of Defensive Symbiosis
6. Summary and Prospect
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Medzhitov, R.; Janeway, C.A., Jr. Innate immunity: The virtues of a nonclonal system of recognition. Cell 1997, 91, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Muller, U.; Vogel, P.; Alber, G.; Schaub, G.A. The innate immune system of mammals and insects. Contrib. Microbiol. 2008, 15, 21–44. [Google Scholar] [CrossRef]
- Seufi, A.M.; Assar, A.A.; Aboelmahasen, M.M.; Mahmoud, S.H. Identification and phylogenetic analyses of two isoforms of the antibacterial gene diptericin from the larval tissue of Musca domestica (Diptera: Muscidae). Erciyes Med. J. 2017, 39, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Liu, L.; Zhao, Y.; Yang, L.; Chen, C.; Li, Z.; Lu, Z. JNK pathway plays a key role in the immune system of the pea aphid and is regulated by microRNA-184. PLoS Pathog. 2020, 16, e1008627. [Google Scholar] [CrossRef] [PubMed]
- Sulek, M.; Kordaczuk, J.; Wojda, I. Current understanding of immune priming phenomena in insects. J. Invertebr. Pathol. 2021, 185, 107656. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.; Yu, L.; Fan, H.; Wang, Z.; Xu, H.; Xi, Y.; Zhu, Z.; Zhou, W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genome Biol. 2014, 15, 521. [Google Scholar] [CrossRef] [Green Version]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Tan, M.; Li, Y.; Zheng, L.; Xu, J.; Jiang, D. The immunotoxicity of Cd exposure to gypsy moth larvae: An integrated analysis of cellular immunity and humoral immunity. Ecotoxicol. Environ. Saf. 2022, 235, 113434. [Google Scholar] [CrossRef]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Zhang, W.; Heryanto, C.; Mohamed, A.; Contreras, G.; Tettamanti, G.; Wink, M.; Bassal, T. Diversity of insect antimicrobial peptides and proteins—A functional perspective: A review. Int. J. Biol. Macromol. 2021, 191, 277–287. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, W.; Lu, Y. Biodegradation of insecticides by gut bacteria isolated from stored grain beetles and its implication in host insecticide resistance. J. Stored Prod. Res. 2022, 96, 101943. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut microbiota mediate insecticide resistance in the diamondback moth, Plutella xylostella (L.). Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A.; et al. Gut microbiome dysbiosis and honeybee health. J. Appl. Entomol. 2011, 135, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, K.L.; King, K.C. Symbiont-mediated immune priming in animals through an evolutionary lens. Microbiology 2022, 168, 001181. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florez, L.V.; Biedermann, P.H.W.; Engl, T.; Kaltenpoth, M. Defensive symbioses of animals with prokaryotic and eukaryotic microorganisms. Nat. Prod. Rep. 2015, 32, 904–936. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wang, J.; Liu, C.; Wang, C.; Walker, T.; Wang, Y. Differentially expressed profiles in the larval testes of Wolbachia infected and uninfected Drosophila. BMC Genom. 2011, 12, 595. [Google Scholar] [CrossRef] [Green Version]
- Horak, R.D.; Leonard, S.P.; Moran, N.A. Symbionts shape host innate immunity in honeybees. Proc. R. Soc. B 2020, 287, 20201184. [Google Scholar] [CrossRef]
- Bai, L.; Wang, L.; Vega-Rodriguez, J.; Wang, G.; Wang, S. A gut symbiotic bacterium Serratia marcescens renders mosquito resistance to Plasmodium infection through activation of mosquito immune responses. Front. Microbiol. 2019, 10, 1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Lee, B.L. Insect symbiosis and immunity: The bean bug-Burkholderia interaction as a case study. Adv. Insect Phys. 2017, 52, 179–197. [Google Scholar] [CrossRef]
- Weiss, B.L.; Wang, J.; Aksoy, S. Tsetse immune system maturation requires the presence of obligate symbionts in larvae. PLoS Biol. 2011, 9, e1000619. [Google Scholar] [CrossRef] [Green Version]
- Weiss, B.L.; Maltz, M.; Aksoy, S. Obligate symbionts activate immune system development in the tsetse fly. J. Immunol. 2012, 188, 3395–3403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Vigneron, A.; Broderick, N.A.; Wu, Y.; Sun, J.S.; Carlson, J.R.; Aksoy, S.; Weiss, B.L. Symbiont-induced odorant binding proteins mediate insect host hematopoiesis. ELife 2017, 6, e19535. [Google Scholar] [CrossRef]
- Prakash, A.; Khan, I. Why do insects evolve immune priming? A search for crossroads. Dev. Comp. Immunol. 2022, 126, 104246. [Google Scholar] [CrossRef] [PubMed]
- Futo, M.; Sell, M.P.; Kutzer, M.A.M.; Kurtz, J. Specificity of oral immune priming in the red flour beetle Tribolium castaneum. Biol. Lett. 2017, 13, 20170632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, J.L.; de Almeida Oliveira, G.; Calvo, E.; Dalli, J.; Colas, R.A.; Serhan, C.N.; Ribeiro, J.M.; Barillas-Mury, C. A mosquito lipoxin/lipocalin complex mediates innate immune priming in Anopheles gambiae. Nat. Commun. 2015, 6, 7403. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.; Brayner, F.A.; Alves, L.C.; Dixit, R.; Barillas-Mury, C. Hemocyte differentiation mediates innate immune memory in Anopheles gambiae mosquitoes. Science 2010, 329, 1353–1355. [Google Scholar] [CrossRef] [Green Version]
- Kaltenpoth, M. Actinobacteria as mutualists: General healthcare for insects? Trends Microbiol. 2009, 17, 529–535. [Google Scholar] [CrossRef]
- Kaltenpoth, M.; Engl, T. Defensive microbial symbionts in Hymenoptera. Funct. Ecol. 2014, 28, 315–327. [Google Scholar] [CrossRef]
- Mattoso, T.C.; Moreira, D.D.O.; Samuels, R.I. Symbiotic bacteria on the cuticle of the leaf-cutting ant Acromyrmex subterraneus subterraneus protect workers from attack by entomopathogenic fungi. Biol. Lett. 2012, 8, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Florez, L.V.; Scherlach, K.; Gaube, P.; Ross, C.; Sitte, E.; Hermes, C.; Rodrigues, A.; Hertweck, C.; Kaltenpoth, M. Antibiotic-producing symbionts dynamically transition between plant pathogenicity and insect-defensive mutualism. Nat. Commun. 2017, 8, 15172. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Chemical barriers to gut infection in the desert locust: In vivo production of antimicrobial phenols associated with the bacterium Pantoea agglomerans. J. Invertebr. Pathol. 1995, 66, 72–75. [Google Scholar] [CrossRef]
- Wang, S.; Dos-Santos, A.L.A.; Huang, W.; Liu, K.C.; Oshaghi, M.A.; Wei, G.; Agre, P.; Jacobs-Lorena, M. Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science 2017, 357, 1399–1402. [Google Scholar] [CrossRef] [Green Version]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C.J. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Mao, Q.; Chen, Y.; Liu, Y.; Chen, Q.; Wu, W.; Zhang, X.; Chen, H.; Li, Y.; Wei, T. Insect symbiotic bacteria harbour viral pathogens for transovarial transmission. Nat. Microbiol. 2017, 2, 17025. [Google Scholar] [CrossRef] [PubMed]
- Morella, N.M.; Koskella, B. The value of a comparative approach to understand the complex interplay between microbiota and host immunity. Front. Immunol. 2017, 8, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Martinez, P.; Naseri, B.; Navarro-Cerrillo, G.; Escriche, B.; Ferre, J.; Herrero, S. Increase in midgut microbiota load induces an apparent immune priming and increases tolerance to Bacillus thuringiensis. Environ. Microbiol. 2010, 12, 2730–2737. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Buckling, A.; Charnley, A.K. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Benyacoub, J.; Bosco, N.; Blanchard, C.; Demont, A.; Philippe, D.; Castiel-Higounenc, I.; Gueniche, A. Immune modulation property of Lactobacillus paracasei NCC2461 (ST11) strain and impact on skin defences. Benef. Microbes 2014, 5, 129–136. [Google Scholar] [CrossRef]
- Ferrarini, M.G.; Dell’Aglio, E.; Vallier, A.; Balmand, S.; Vincent-Monegat, C.; Hughes, S.; Gillet, B.; Parisot, N.; Zaidman-Remy, A.; Vieira, C.; et al. Efficient compartmentalization in insect bacteriomes protects symbiotic bacteria from host immune system. Microbiome 2022, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Wang, P.; Shi, H.; Si, F.; Hao, Y.; Chen, B. Fas-associated factor 1 plays a negative regulatory role in the antibacterial immunity of Locusta migratoria. Insect Mol. Biol. 2013, 22, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Zug, R.; Hammerstein, P. Wolbachia and the insect immune system: What reactive oxygen species can tell us about the mechanisms of Wolbachia-host interactions. Front. Microbiol. 2015, 6, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laughton, A.M.; Garcia, J.R.; Altincicek, B.; Strand, M.R.; Gerardo, N.M. Characterisation of immune responses in the pea aphid, Acyrthosiphon pisum. J. Insect Physiol. 2011, 57, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Zaidman-Remy, A.; Vigneron, A.; Weiss, B.L.; Heddi, A. What can a weevil teach a fly, and reciprocally? Interaction of host immune systems with endosymbionts in Glossina and Sitophilus. BMC Microbiol. 2018, 18, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Gerardo, N.M.; Hoang, K.L.; Stoy, K.S. Evolution of animal immunity in the light of beneficial symbioses. Philos. Trans. R. Soc. B 2020, 375, 20190601. [Google Scholar] [CrossRef]
- Price, D.R.G.; Duncan, R.P.; Shigenobu, S.; Wilson, A.C.C. Genome expansion and differential expression of amino acid transporters at the aphid/Buchnera symbiotic interface. Mol. Biol. Evol. 2011, 28, 3113–3126. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Desneux, N.; Barta, R.J.; Hoelmer, K.A.; Hopper, K.R.; Heimpel, G.E. Multifaceted determinants of host specificity in an aphid parasitoid. Oecologia 2009, 160, 387–398. [Google Scholar] [CrossRef]
- Li, S.; de Mandal, S.; Xu, X.; Jin, F. The tripartite interaction of host immunity-Bacillus thuringiensis infection-gut microbiota. Toxins 2020, 12, 514. [Google Scholar] [CrossRef]
- Bai, S.; Yao, Z.; Raza, M.F.; Cai, Z.; Zhang, H. Regulatory mechanisms of microbial homeostasis in insect gut. Insect Sci. 2021, 28, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Lee, K.A.; Hyeon, D.Y.; Kyung, M.; Jun, K.Y.; Seo, S.H.; Hwang, D.; Kwon, Y.; Lee, W.J. Bacterial nucleoside catabolism controls quorum sensing and commensal-to-pathogen transition in the Drosophila gut. Cell Host Microbe 2020, 27, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial world: Insights from Drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, Y.; Zhou, J.; Zhang, Y.; Ma, Y.; Wang, D.; Jiang, Y.; Shi, S.; Qin, L. Peptidoglycan recognition proteins regulate immune response of Antheraea pernyi in different ways. J. Invertebr. Pathol. 2019, 166, 107204. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Kim, S.H.; Kim, E.K.; Ha, E.M.; You, H.; Kim, B.; Kim, M.J.; Kwon, Y.; Ryu, J.H.; Lee, W.J. Bacterial-derived uracil as a modulator of mucosal immunity and gut-microbe homeostasis in Drosophila. Cell 2013, 153, 797–811. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Su, F.; Li, Q.; Zhang, J.; Li, Y.; Tang, T.; Hu, Q.; Yu, X. Pattern recognition receptors in Drosophila immune responses. Dev. Comp. Immunol. 2020, 102, 103468. [Google Scholar] [CrossRef]
- Bischoff, V.; Vignal, C.; Duvic, B.; Boneca, I.G.; Hoffmann, J.A.; Royet, J. Downregulation of the Drosophila immune response by peptidoglycan-recognition proteins SC1 and SC2. PLoS Pathog. 2006, 2, e14. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.M.; Cirimotich, C.M.; Dong, Y.; Dimopoulos, G. Caudal is a negative regulator of the Anopheles IMD pathway that controls resistance to Plasmodium falciparum infection. Dev. Comp. Immunol. 2013, 39, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lee, W.J. Role of DUOX in gut inflammation: Lessons from Drosophila model of gut-microbiota interactions. Front. Cell Infect. Microbiol. 2014, 3, 116. [Google Scholar] [CrossRef] [Green Version]
- Ha, E.M.; Lee, K.A.; Seo, Y.Y.; Kim, S.H.; Lim, J.H.; Oh, B.H.; Kim, J.; Lee, W.J. Coordination of multiple dual oxidase-regulatory pathways in responses to commensal and infectious microbes in Drosophila gut. Nat. Immunol. 2009, 10, 949–957. [Google Scholar] [CrossRef]
- Ha, E.M.; Oh, C.T.; Bae, Y.S.; Lee, W.J. A direct role for dual oxidase in Drosophila gut immunity. Science 2005, 310, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S.; Choi, M.K.; Lee, W.J. Dual oxidase in mucosal immunity and host-microbe homeostasis. Trends Immunol. 2010, 31, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Kim, B.; Bhin, J.; Kim, D.H.; You, H.; Kim, E.K.; Kim, S.H.; Ryu, J.H.; Hwang, D.; Lee, W.J. Bacterial uracil modulates Drosophila DUOX-dependent gut immunity via Hedgehog-induced signaling endosomes. Cell Host Microbe 2015, 17, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, K.L.; Stepien, T.A.; Blum, J.E.; Holt, J.F.; Labbe, N.H.; Rush, J.S.; Raffa, K.F.; Handelsman, J. From commensal to pathogen: Translocation of Enterococcus faecalis from the midgut to the hemocoel of Manduca sexta. mBio 2011, 2, e00065-11. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Housing microbial symbionts: Evolutionary origins and diversification of symbiotic organs in animals. Philos. Trans. R. Soc. B 2020, 375, 20190603. [Google Scholar] [CrossRef]
- Braendle, C.; Miura, T.; Bickel, R.; Shingleton, A.W.; Kambhampati, S.; Stern, D.L. Developmental origin and evolution of bacteriocytes in the aphid-Buchnera symbiosis. PLoS Biol. 2003, 1, e21. [Google Scholar] [CrossRef] [Green Version]
- Michalik, A.; Franco, D.C.; Deng, J.; Szklarzewicz, T.; Stroinski, A.; Kobialka, M.; Lukasik, P. Variable organization of symbiont-containing tissue across planthoppers hosting different heritable endosymbionts. Front. Physiol. 2023, 14, 1135346. [Google Scholar] [CrossRef]
- Wang, D.; Huang, Z.; Billen, J.; Zhang, G.; He, H.; Wei, C. Structural diversity of symbionts and related cellular mechanisms underlying vertical symbiont transmission in cicadas. Environ. Microbiol. 2021, 23, 6603–6621. [Google Scholar] [CrossRef]
- Kucuk, R.A. Gut bacteria in the Holometabola: A review of obligate and facultative symbionts. J. Insect Sci. 2020, 20, 22. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Y.; Yang, G.; Aksoy, S. Interactions between mutualist Wigglesworthia and tsetse peptidoglycan recognition protein (PGRP-LB) influence trypanosome transmission. Proc. Natl. Acad. Sci. USA 2009, 106, 12133–12138. [Google Scholar] [CrossRef] [Green Version]
- Login, F.H.; Heddi, A. Insect immune system maintains long-term resident bacteria through a local response. J. Insect Physiol. 2013, 59, 232–239. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Ma, Q.; Wood, T.K. OmpA influences Escherichia coli biofilm formation by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 2009, 11, 2735–2746. [Google Scholar] [CrossRef]
- Kint, G.; de Coster, D.; Marchal, K.; Vanderleyden, J.; de Keersmaecker, S.C.J. The small regulatory RNA molecule MicA is involved in Salmonella enterica serovar Typhimurium biofilm formation. BMC Microbiol. 2010, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect. Immun. 2009, 77, 3150–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Leger, R.J.S. A collagenous protective coat enables Metarhizium anisopliae to evade insect immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 6647–6652. [Google Scholar] [CrossRef] [Green Version]
- Moser, C.; Jensen, P.O.; Thomsen, K.; Kolpen, M.; Rybtke, M.; Lauland, A.S.; Trostrup, H.; Tolker-Nielsen, T. Immune responses to Pseudomonas aeruginosa biofilm infections. Front. Immunol. 2021, 12, 625597. [Google Scholar] [CrossRef]
- Mulcahy, H.; Sibley, C.D.; Surette, M.G.; Lewenza, S. Drosophila melanogaster as an animal model for the study of Pseudomonas aeruginosa biofilm infections in vivo. PLoS Pathog. 2011, 7, e1002299. [Google Scholar] [CrossRef] [Green Version]
- Maltz, M.A.; Weiss, B.L.; O’Neill, M.; Wu, Y.; Aksoy, S. OmpA-mediated biofilm formation is essential for the commensal bacterium Sodalis glossinidius to colonize the tsetse fly gut. Appl. Environ. Microbiol. 2012, 78, 7760–7768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, H.; Weiss, B.L.; Perkin, S.A.H.; Yamashita, A.; Oshima, K.; Hattori, M.; Aksoy, S. Massive genome erosion and functional adaptations provide insights into the symbiotic lifestyle of Sodalis glossinidius in the tsetse host. Genome Res. 2006, 16, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, B.L.; Wu, Y.; Schwank, J.J.; Tolwinski, N.S.; Aksoy, S. An insect symbiosis is influenced by bacterium-specific polymorphisms in outer-membrane protein A. Proc. Natl. Acad. Sci. USA 2008, 105, 15088–15093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, M.J.; Kim, J.E.; Chung, Y.H.; Lee, W.B.; Shin, Y.K.; Lee, J.S.; Kim, D.; Park, Y.M. Dendritic cells stimulated with outer membrane protein A (OmpA) of Salmonella typhimurium generate effective anti-tumor immunity. Vaccine 2011, 29, 2400–2410. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Chen, M.; Yue, L.; Xing, K.; Li, T.; Kang, K.; Liang, Z.; Yuan, L.; Zhang, W. A distinct strain of Arsenophonus symbiont decreases insecticide resistance in its insect host. PLoS Genet. 2018, 14, e1007725. [Google Scholar] [CrossRef]
- Feng, H.; Edwards, N.; Anderson, C.M.H.; Althaus, M.; Duncan, R.P.; Hsu, Y.C.; Luetje, C.W.; Price, D.R.G.; Wilson, A.C.C.; Thwaites, D.T. Trading amino acids at the aphid-Buchnera symbiotic interface. Proc. Natl. Acad. Sci. USA 2019, 116, 16003–16011. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Nair, S. Dynamics of insect-microbiome interaction influence host and microbial symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef]
- Latorre, A.; Manzano-Marín, A. Dissecting genome reduction and trait loss in insect endosymbionts. Ann. N. Y. Acad. Sci. 2017, 1389, 52–75. [Google Scholar] [CrossRef]
- Kim, J.K.; Son, D.W.; Kim, C.H.; Cho, J.H.; Marchetti, R.; Silipo, A.; Sturiale, L.; Park, H.Y.; Huh, Y.R.; Nakayama, H.; et al. Insect gut symbiont susceptibility to host antimicrobial peptides caused by alteration of the bacterial cell envelope. J. Biol. Chem. 2015, 290, 21042–21053. [Google Scholar] [CrossRef] [Green Version]
- Chong, R.A.; Moran, N.A. Intraspecific genetic variation in hosts affects regulation of obligate heritable symbionts. Proc. Natl. Acad. Sci. USA 2016, 113, 13114–13119. [Google Scholar] [CrossRef] [Green Version]
- Chrostek, E.; Teixeira, L. Within host selection for faster replicating bacterial symbionts. PLoS ONE 2018, 13, e0191530. [Google Scholar] [CrossRef] [Green Version]
- Parker, B.J.; Hrcek, J.; McLean, A.H.C.; Brisson, J.A.; Godfray, H.C.J. Intraspecific variation in symbiont density in an insect-microbe symbiosis. Mol. Ecol. 2021, 30, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Nichols, H.L.; Goldstein, E.B.; Ziabari, O.S.; Parker, B.J. Intraspecific variation in immune gene expression and heritable symbiont density. PLoS Pathog. 2021, 17, e1009552. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, H. Importance of endosymbionts Wolbachia and Rickettsia in insect resistance development. Curr. Opin. Insect Sci. 2019, 33, 84–90. [Google Scholar] [CrossRef]
- Engl, T.; Kaltenpoth, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Huang, X.; Yin, Y. Beyond immunity: The Imd pathway as a coordinator of host defense, organismal physiology and behavior. Dev. Comp. Immunol. 2018, 83, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Dutra, H.L.C.; Sucupira, P.H.F.; Ferreira, A.G.A.; Moreira, L.A. Wolbachia as translational science: Controlling mosquito-borne pathogens. Trends Parasitol. 2021, 37, 1050–1067. [Google Scholar] [CrossRef]
- Thomas, S.; Verma, J.; Woolfit, M.; OʼNeill, S.L. Wolbachia-mediated virus blocking in mosquito cells is dependent on XRN1-mediated viral RNA degradation and influenced by viral replication rate. PLoS Pathog. 2018, 14, e1006879. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.P.; Powell, J.E.; Perutka, J.; Geng, P.; Heckmann, L.C.; Horak, R.D.; Davies, B.W.; Ellington, A.D.; Barrick, J.E.; Moran, N.A. Engineered symbionts activate honey bee immunity and limit pathogens. Science 2020, 367, 573–576. [Google Scholar] [CrossRef]
- Schmidt, K.; Engel, P. Mechanisms underlying gut microbiota-host interactions in insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef]
- Xie, S.; Lan, Y.; Sun, C.; Shao, Y. Insect microbial symbionts as a novel source for biotechnology. World J. Microbiol. Biotechnol. 2019, 35, 25. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, B.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the lepidopteran gut microbiota. Cell Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Omics and the metabolic function of insect-microbial symbioses. Curr. Opin. Insect Sci. 2018, 29, 1–6. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Partner | Influence from Symbiosis | Selection in Evolution | Evolutionary Pattern | |
|---|---|---|---|---|
| Insect hosts | Positive | Improved immunity | To improve communication efficiency between symbiotic partners | Symbiosomal membranes produced by the host with selective permeability of metabolites [84] |
| Negative | Excessive energy consumption in immune responses | To maintain symbiont homeostasis | Degeneration of the host immune system [44] Fine-tuned regulation of immune responses by host negative regulators [45] Compartmentalization of the symbionts in specialized symbiotic organs [46] | |
| Symbionts | Positive | Stable ecological niche (environment/nutrition) | To allocate more energetic resources to colonization resistance | Degeneration or massive pseudogenization of the genome [45,85] |
| Negative | Damage from host immune responses | To avoid recognition by the host immune system and to escape from host immune responses | Formation of biofilms [79] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Yong, H.; Zhang, S.; Liu, Z.; Zhao, Y. Colonization Resistance of Symbionts in Their Insect Hosts. Insects 2023, 14, 594. https://doi.org/10.3390/insects14070594
Wang Z, Yong H, Zhang S, Liu Z, Zhao Y. Colonization Resistance of Symbionts in Their Insect Hosts. Insects. 2023; 14(7):594. https://doi.org/10.3390/insects14070594
Chicago/Turabian StyleWang, Zhengyan, Hanzi Yong, Shan Zhang, Zhiyuan Liu, and Yaru Zhao. 2023. "Colonization Resistance of Symbionts in Their Insect Hosts" Insects 14, no. 7: 594. https://doi.org/10.3390/insects14070594
APA StyleWang, Z., Yong, H., Zhang, S., Liu, Z., & Zhao, Y. (2023). Colonization Resistance of Symbionts in Their Insect Hosts. Insects, 14(7), 594. https://doi.org/10.3390/insects14070594