Bumble Bees (Bombus terrestris) Use Time-Memory to Associate Reward with Color and Time of Day



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bees
2.2. Time Training
2.3. Statistical Analysis
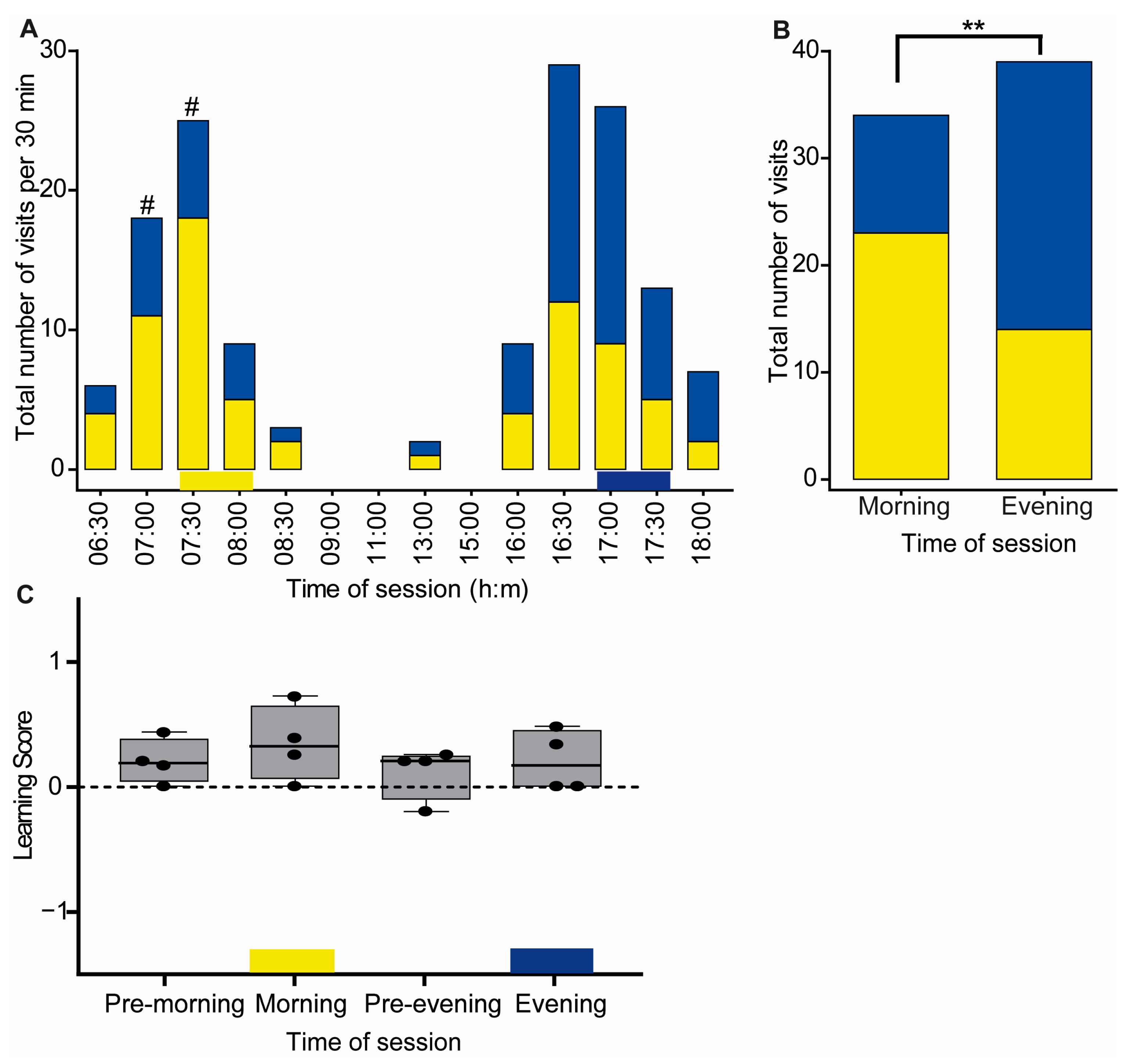
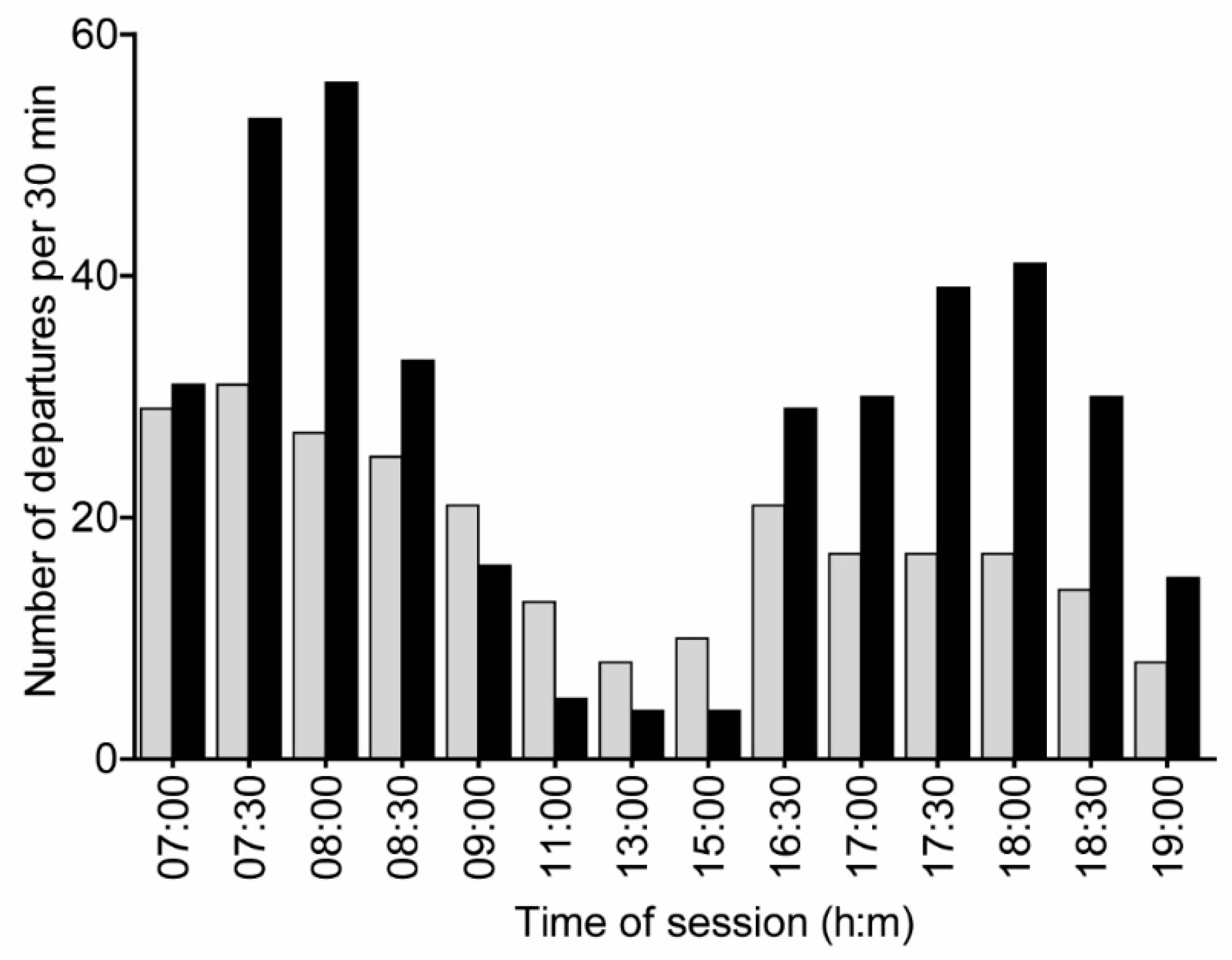
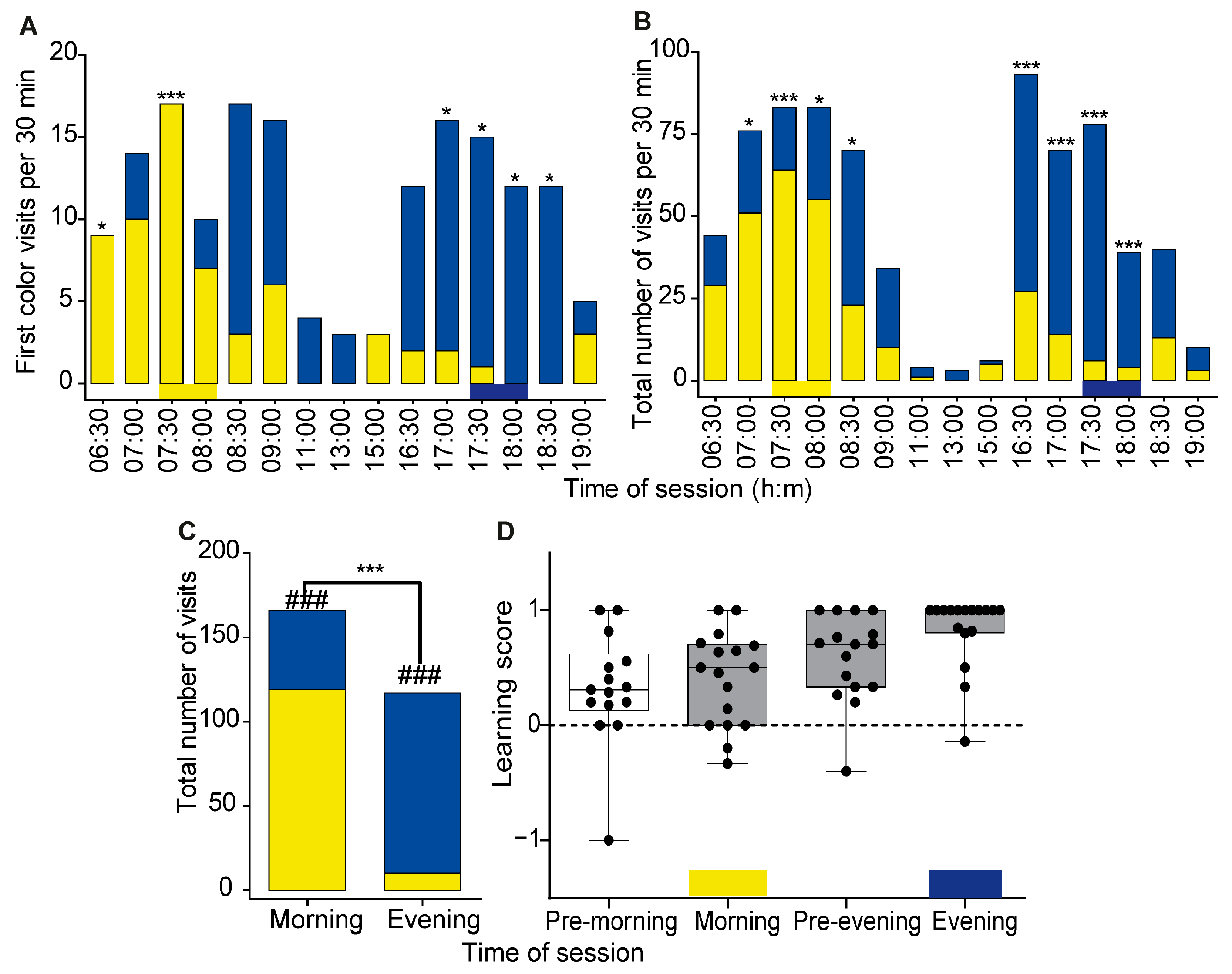
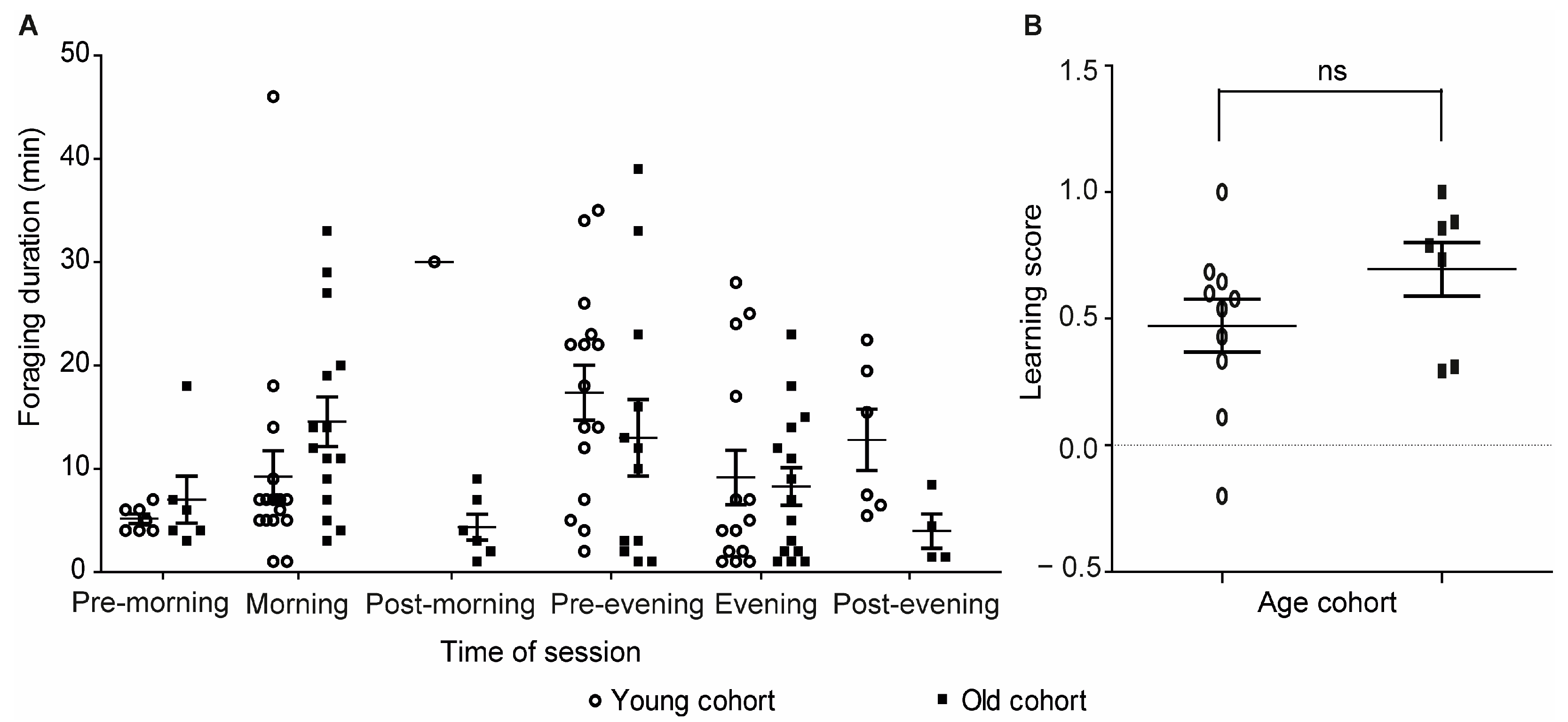
3. Results
3.1. Experiment 1
3.2. Experiment 2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benirschke, K. Chronobiology: Biological timekeeping. J. Hered. 2004, 95, 91–92. [Google Scholar] [CrossRef]
- Helm, B.; Visser, M.E.; Schwartz, W.; Kronfeld-Schor, N.; Gerkema, M.; Piersma, T.; Bloch, G. Two sides of a coin: Ecological and chronobiological perspectives of timing in the wild. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160246. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.K. Adaptive significance of circadian clocks. Chronobiol. Int. 2003, 20, 901–919. [Google Scholar] [CrossRef] [PubMed]
- Yerushalmi, S.; Green, R.M. Evidence for the adaptive significance of circadian rhythms. Ecol. Lett. 2009, 12, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Allada, R.; Chung, B.Y. Circadian organization of behavior and physiology in Drosophila. Annu. Rev. Physiol. 2010, 72, 605–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubowy, C.; Sehgal, A. Circadian rhythms and sleep in Drosophila melanogaster. Genetics 2017, 205, 1373–1397. [Google Scholar] [CrossRef] [Green Version]
- Manjunatha, T.; Dass, H.; Sharma, S.K. Egg-laying rhythm in Drosophila melanogaster. J. Genet. 2008, 87, 495–504. [Google Scholar]
- Saunders, D.S.; Steel, C.G.H.; Vafopoulou, X.; Lewis, R.D. Insect Clocks; Elsevier: Amsterdam, The Netherlands, 2002; pp. 449–472. [Google Scholar] [CrossRef]
- Tataroglu, O.; Emery, P. Studying circadian rhythms in Drosophila melanogaster. Methods 2014, 68, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Bloch, G. Plasticity in the circadian clock and the temporal organization of insect societies. In Organization of Insect Societies; Harvard University Press: Cambridge, MA, USA, 2009; pp. 402–432. [Google Scholar] [CrossRef]
- Moore, D. Mini-Review Honey bee circadian clocks: Behavioral control from individual workers to whole-colony rhythms. J. Insect Physiol. 2001, 47, 843–857. [Google Scholar] [CrossRef]
- Shpigler, H.Y.; Yaniv, A.; Gernat, T.; Robinson, G.E.; Bloch, G. The influences of illumination regime on egg-laying rhythms of honey bee queens. J. Biol. Rhythm. 2022, 37, 609–619. [Google Scholar] [CrossRef]
- Beekman, M.; Ratnieks, F.L.W. Long-range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Von Frisch, K. The Dance Language and Orientation of Bees; Harvard University Press: Cambridge, MA, USA, 1967. [Google Scholar] [CrossRef]
- Biebach, H.; Gordijn, M.; Krebst, J.R. Time-and-place learning by Garden Warblers, Sylvia borin. Anim. Behav. 1989, 37, 353–360. [Google Scholar] [CrossRef]
- Chouhan, N.S.; Wolf, R.; Helfrich-Förster, C.; Heisenberg, M. Flies remember the time of day. Curr. Biol. 2015, 25, 1619–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.M.; Breed, M.D. Temporal learning in the giant tropical ant, Paraponera clavata. Physiol. Entomol. 1987, 12, 317–320. [Google Scholar] [CrossRef]
- Reebs, S.G. Time-place learning in Golden Shiners (Pisces: Cyprinidae). Behav. Process. 1996, 36, 253–262. [Google Scholar] [CrossRef]
- Van der Zee, E.A.; Havekes, R.; Barf, R.P.; Hut, R.A.; Nijholt, I.M.; Jacobs, E.H.; Gerkema, M.P. Circadian time-place learning in mice depends on Cry genes. Curr. Biol. 2008, 18, 844–848. [Google Scholar] [CrossRef] [Green Version]
- Beling, I. Uber Das Zeitgedachtnis Der Bienen. J. Comp. Physiol. 1929, 9, 259–338. [Google Scholar]
- Wahl, O. Beitrag zur frage der biologischen bedeutung des Zeitgedächtnisses der bienen. Z. Vgl. Physiol. 1932, 18, 709–717. [Google Scholar] [CrossRef]
- Woodard, S.H.; Lozier, J.D.; Goulson, D.; Williams, P.H.; Strange, J.P.; Jha, S. Molecular tools and bumble bees: Revealing hidden details of ecology and evolution in a model system. Mol. Ecol. 2015, 24, 2916–2936. [Google Scholar] [CrossRef]
- Cueva Del Castillo, R.; Sanabria-Urb An, S.; Serrano-Meneses, M.A.; Serrano-Meneses, A. Trade-offs in the evolution of bumblebee colony and body size: A comparative analysis. Ecol. Evol. 2015, 5, 3914–3926. [Google Scholar] [CrossRef]
- Walther-Hellwig, K.; Frankl, R. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Wolf, S.; Moritz, R.F.A. Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 2008, 39, 419–427. [Google Scholar] [CrossRef]
- Dornhaus, A.; Chittka, L. Information flow and regulation of foraging activity in bumble bees (Bombus spp.). Apidologie 2004, 35, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Dornhaus, A.; Chittka, L. Bumble bees (Bombus terrestris) store both food and information in honeypots. Behav. Ecol. 2005, 16, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Renner, M.A.; Nieh, J.C. Bumble bee olfactory information flow and contact-based foraging activation. Insectes Sociaux 2008, 55, 417–424. [Google Scholar] [CrossRef]
- Pearce, R.F.; Giuggioli, L.; Rands, S.A. Bumblebees can discriminate between scent-marks deposited by conspecifics. Sci. Rep. 2017, 7, 43872. [Google Scholar] [CrossRef] [Green Version]
- Saleh, N.; Scott, A.G.; Bryning, G.P.; Chittka, L. Distinguishing signals and cues: Bumblebees use general footprints to generate adaptive behaviour at flowers and nest. Arthropod-Plant Interact. 2007, 1, 119–127. [Google Scholar] [CrossRef]
- Schmitt, U. Hydrocarbons in tarsal glands of Bombus terrestris. Experientia 1990, 46, 1080–1082. [Google Scholar]
- Sunrise and Sunset Times in Jerusalem. Available online: https://www.timeanddate.com/sun/israel/jerusalem?month=9&year=2020&as_qdr=y15 (accessed on 2 August 2022).
- Weather in Jerusalem. 2020. Available online: https://www.timeanddate.com/weather/israel/jerusalem/historic?month=9&year=2020 (accessed on 3 August 2022).
- Sunrise and Sunset Times in Jerusalem. Available online: https://www.timeanddate.com/sun/israel/jerusalem?month=7&year=2021 (accessed on 1 December 2022).
- Weather in Jerusalem. 2021. Available online: https://www.timeanddate.com/weather/israel/jerusalem/historic?month=7&year=2021 (accessed on 1 December 2022).
- Yerushalmi, S.; Bodenhaimer, S.; Bloch, G. Developmentally determined attenuation in circadian rhythms links chronobiology to social organization in bees. J. Exp. Biol. 2006, 209, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, D.J.; Levy, R.; Scheepers, C.; Tily, H.J. Random effects structure for confirmatory hypothesis testing: Keep it maximal. J. Mem. Lang. 2013, 68, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using Lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lehmann, M.; Gustav, D.; Galizia, C.G. The early bee catches the flower—Circadian rhythmicity influences learning performance in honey bees, Apis mellifera. Behav. Ecol. Sociobiol. 2011, 65, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, D.; Siegfried, D.; Wilson, R.; Rankin, M.A. The influence of time of day on the foraging behavior of the honeybee, Apis mellifera. J. Biol. Rhythm. 1989, 4, 305–325. [Google Scholar]
- Moore, D.; Rankin, M.A. Diurnal changes in the accuracy of the honey bee foraging rhythm. Biol. Bull. 1983, 164, 471–482. [Google Scholar] [CrossRef]
- Collett, T.S.; de Ibarra, N.H.; Riabinina, O.; Philippides, A. Coordinating compass-based and nest-based flight directions during bumblebee learning and return flights. J. Exp. Biol. 2013, 216 Pt 6, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Menzel, R. Serial position learning in honeybees. PLoS ONE 2009, 4, e4694. [Google Scholar]
- Moore, D.; Doherty, P. Acquisition of a time-memory in forager honey bees. J. Comp. Physiol. A 2009, 195, 741–751. [Google Scholar] [CrossRef]
- Wagner, A.E.; Van Nest, B.N.; Hobbs, C.N.; Moore, D. Persistence, reticence and the management of multiple time memories by forager honey bees. J. Exp. Biol. 2013, 216, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Chole, H.; Woodard, S.H.; Bloch, G. Body size variation in bees: Regulation, mechanisms, and relationship to social organization. Curr. Opin. Insect Sci. 2019, 35, 77–87. [Google Scholar] [CrossRef]
- Free, J.B. The division of labour within bumblebee colonies. Insectes Sociaux 1955, 2, 195–212. [Google Scholar] [CrossRef]
- Jandt, J.M.; Dornhaus, A. Spatial organization and division of labour in the bumblebee Bombus impatiens. Anim. Behav. 2009, 77, 641–651. [Google Scholar] [CrossRef]
- Spaethe, J.; Weidenmüller, A. Size variation and foraging rate in bumblebees (Bombus terrestris). Insectes Sociaux 2002, 49, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Riveros, A.J.; Gronenberg, W. Olfactory learning and memory in the bumblebee Bombus occidentalis. Naturwissenschaften 2009, 96, 851–856. [Google Scholar] [CrossRef]
- Raine, N.E.; Chittka, L. The correlation of learning speed and natural foraging success in bumble-bees. Proc. R. Soc. B Biol. Sci. 2008, 275, 803–808. [Google Scholar] [CrossRef]
- Mulder, C.; Gerkema, M.P.; Van der Zee, E.A. Circadian clocks and memory: Time-place learning. Front. Mol. Neurosci. 2013, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, G.; Bar-Shai, N.; Cytter, Y.; Green, R. Time is honey: Circadian clocks of bees and flowers and how their interactions may influence ecological communities. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160256. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonulkirmaz-Cancalar, O.; Shertzer, O.; Bloch, G. Bumble Bees (Bombus terrestris) Use Time-Memory to Associate Reward with Color and Time of Day. Insects 2023, 14, 707. https://doi.org/10.3390/insects14080707
Gonulkirmaz-Cancalar O, Shertzer O, Bloch G. Bumble Bees (Bombus terrestris) Use Time-Memory to Associate Reward with Color and Time of Day. Insects. 2023; 14(8):707. https://doi.org/10.3390/insects14080707
Chicago/Turabian StyleGonulkirmaz-Cancalar, Ozlem, Oded Shertzer, and Guy Bloch. 2023. "Bumble Bees (Bombus terrestris) Use Time-Memory to Associate Reward with Color and Time of Day" Insects 14, no. 8: 707. https://doi.org/10.3390/insects14080707
APA StyleGonulkirmaz-Cancalar, O., Shertzer, O., & Bloch, G. (2023). Bumble Bees (Bombus terrestris) Use Time-Memory to Associate Reward with Color and Time of Day. Insects, 14(8), 707. https://doi.org/10.3390/insects14080707