

How Biodiversity, Climate and Landscape Drive Functional Redundancy of British Butterflies

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data and Sampling
2.2. Environmental Data
2.3. Diversity and Redundancy Metrics
2.4. Statistical Analysis
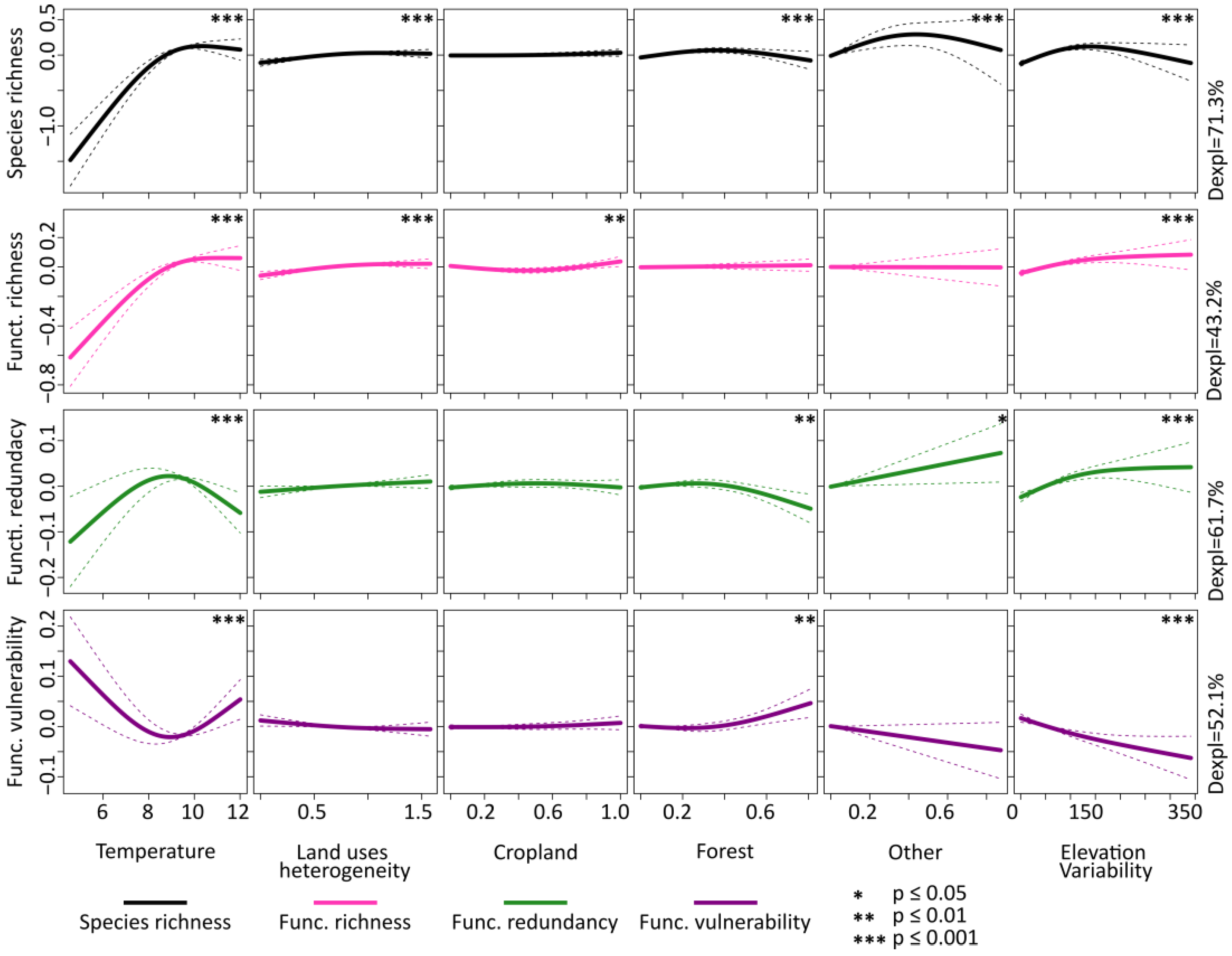
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiedmann, T.; Lenzen, M.; Keyßer, L.T.; Steinberger, J.K. Scientists’ warning on affluence. Nat. Commun. 2020, 11, 3107. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M. Climate change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; Volume 2. [Google Scholar]
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.-O.; Roberts, D.; Skea, J.; Shukla, P.R. Global Warming of 1.5 C: IPCC Special Report on Impacts of Global Warming of 1.5 C above Pre-Industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S. The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2019. [Google Scholar]
- Millhauser, J.K.; Earle, T.K. Biodiversity and the human past: Lessons for conservation biology. Biol. Conserv. 2022, 272, 109599. [Google Scholar] [CrossRef]
- van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef] [PubMed]
- Memtsas, G.; Lazarina, M.; Sgardelis, S.; Petanidou, T.; Kallimanis, A. What plant–pollinator network structure tells us about the mechanisms underlying the bidirectional biodiversity productivity relationship? Basic Appl. Ecol. 2022, 63, 49–58. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Mammola, S.; Carmona, C.P.; Guillerme, T.; Cardoso, P. Concepts and applications in functional diversity. Funct. Ecol. 2021, 35, 1869–1885. [Google Scholar] [CrossRef]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J.; Pither, J.; Kerkhoff, A.J.; Boyle, B.; Weiser, M.D.; Elser, J.J.; Fagan, W.F.; Forero-Montaña, J.; Fyllas, N. The biogeography and filtering of woody plant functional diversity in North and South America. Glob. Ecol. Biogeogr. 2012, 21, 798–808. [Google Scholar] [CrossRef]
- Mazel, F.; Guilhaumon, F.; Mouquet, N.; Devictor, V.; Gravel, D.; Renaud, J.; Cianciaruso, M.V.; Loyola, R.; Diniz-Filho, J.A.F.; Mouillot, D. Multifaceted diversity–area relationships reveal global hotspots of mammalian species, trait and lineage diversity. Glob. Ecol. Biogeogr. 2014, 23, 836–847. [Google Scholar] [CrossRef]
- Tsianou, M.A.; Kallimanis, A.S. Different species traits produce diverse spatial functional diversity patterns of amphibians. Biodivers. Conserv. 2016, 25, 117–132. [Google Scholar] [CrossRef]
- Mason-Romo, E.D.; Farías, A.A.; Ceballos, G. Two decades of climate driving the dynamics of functional and taxonomic diversity of a tropical small mammal community in western Mexico. PLoS ONE 2017, 12, e0189104. [Google Scholar] [CrossRef] [PubMed]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Naeem, S. Species redundancy and ecosystem reliability. Conserv. Biol. 1998, 12, 39–45. [Google Scholar] [CrossRef]
- Biggs, C.R.; Yeager, L.A.; Bolser, D.G.; Bonsell, C.; Dichiera, A.M.; Hou, Z.; Keyser, S.R.; Khursigara, A.J.; Lu, K.; Muth, A.F. Does functional redundancy affect ecological stability and resilience? A review and meta-analysis. Ecosphere 2020, 11, e03184. [Google Scholar] [CrossRef]
- Zobel, M. The relative of species pools in determining plant species richness: An alternative explanation of species coexistence? Trends Ecol. Evol. 1997, 12, 266–269. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Swenson, N.G.; Anglada-Cordero, P.; Barone, J.A. Deterministic tropical tree community turnover: Evidence from patterns of functional beta diversity along an elevational gradient. Proc. R. Soc. B Biol. Sci. 2011, 278, 877–884. [Google Scholar] [CrossRef]
- Mori, A.S.; Isbell, F.; Seidl, R. β-diversity, community assembly, and ecosystem functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef]
- WallisDeVries, M.F. Linking species assemblages to environmental change: Moving beyond the specialist-generalist dichotomy. Basic Appl. Ecol. 2014, 15, 279–287. [Google Scholar] [CrossRef]
- Bonelli, S.; Cerrato, C.; Barbero, F.; Boiani, M.V.; Buffa, G.; Casacci, L.P.; Fracastoro, L.; Provenzale, A.; Rivella, E.; Zaccagno, M. Changes in alpine butterfly communities during the last 40 years. Insects 2021, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Basset, Y.; Lamarre, G.P. Toward a world that values insects. Science 2019, 364, 1230–1231. [Google Scholar] [CrossRef] [PubMed]
- Warren, M.S.; Maes, D.; van Swaay, C.A.; Goffart, P.; Van Dyck, H.; Bourn, N.A.; Wynhoff, I.; Hoare, D.; Ellis, S. The decline of butterflies in Europe: Problems, significance, and possible solutions. Proc. Natl. Acad. Sci. USA 2021, 118, e2002551117. [Google Scholar] [CrossRef] [PubMed]
- National Biodiversity Network Gateway. Available online: https://nbn.org.uk/the-national-biodiversity-network/archive-information/nbn-gateway/ (accessed on 12 December 2012).
- Mouillot, D.; Villéger, S.; Parravicini, V.; Kulbicki, M.; Arias-González, J.E.; Bender, M.; Chabanet, P.; Floeter, S.R.; Friedlander, A.; Vigliola, L. Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc. Natl. Acad. Sci. USA 2014, 111, 13757–13762. [Google Scholar] [CrossRef]
- Gaston, K.J.; Quinn, R.M.; Wood, S.; Arnold, H.R. Measures of geographic range size: The effects of sample size. Ecography 1996, 19, 259–268. [Google Scholar] [CrossRef]
- Warren, M.S.; Barnett, L.K.; Gibbons, D.W.; Avery, M.I. Assessing national conservation priorities: An improved red list of British butterflies. Biol. Conserv. 1997, 82, 317–328. [Google Scholar] [CrossRef]
- Gaston, K.J.; Quinn, R.M.; Blackburn, T.M.; Eversham, B.C. Species-range size distributions in Britain. Ecography 1998, 21, 361–370. [Google Scholar] [CrossRef]
- Asher, J.; Warren, M.; Fox, R.; Harding, P.; Jeffcoate, G.; Jeffcoate, S. The Millennium Atlas of Butterflies in Britain and Ireland; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Brereton, T.; Roy, D.; Middlebrook, I.; Botham, M.; Warren, M. The development of butterfly indicators in the United Kingdom and assessments in 2010. J. Insect Conserv. 2011, 15, 139–151. [Google Scholar] [CrossRef]
- Met Office. Available online: https://catalogue.ceda.ac.uk/uuid/4dc8450d889a491ebb20e724debe2dfb (accessed on 3 May 2023).
- Fuchs, R.; Herold, M.; Verburg, P.H.; Clevers, J.G. A high-resolution and harmonized model approach for reconstructing and analysing historic land changes in Europe. Biogeosciences 2013, 10, 1543–1559. [Google Scholar] [CrossRef]
- Global Human Settlement Layer. Available online: https://ghsl.jrc.ec.europa.eu/download.php?ds=pop (accessed on 16 May 2023).
- Copernicus Land Monitoring Services. Available online: https://land.copernicus.eu/imagery-in-situ/eu-dem/eu-dem-v1.1 (accessed on 17 May 2023).
- Aguirre-Gutiérrez, J.; Kissling, W.D.; Carvalheiro, L.G.; WallisDeVries, M.F.; Franzén, M.; Biesmeijer, J.C. Functional traits help to explain half-century long shifts in pollinator distributions. Sci. Rep. 2016, 6, 24451. [Google Scholar] [CrossRef]
- Middleton-Welling, J.; Dapporto, L.; García-Barros, E.; Wiemers, M.; Nowicki, P.; Plazio, E.; Bonelli, S.; Zaccagno, M.; Šašić, M.; Liparova, J. A new comprehensive trait database of European and Maghreb butterflies, Papilionoidea. Sci. Data 2020, 7, 351. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology, R package version 1.0–12. 2014.
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The vegan package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Baselga, M.A. Package ‘Betapart’: Partitioning Beta Diversity into Turnover and Nestedness Components, R package version 1.5.4. 2018.
- Wood, S.; Wood, M.S. Package ‘mgcv’. R Package Version 2015, 1, 729. [Google Scholar]
- Naimi, B. USDM: Uncertainty Analysis for Species Distribution Models, R package version 1.1–15; R Documentation, 2015.
- Ferrier, S.; Manion, G.; Elith, J.; Richardson, K. Using generalized dissimilarity modelling to analyse and predict patterns of beta diversity in regional biodiversity assessment. Divers. Distrib. 2007, 13, 252–264. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Sanders, N.J.; Normand, S.; Svenning, J.-C.; Ferrier, S.; Gove, A.D.; Dunn, R.R. Environmental and historical imprints on beta diversity: Insights from variation in rates of species turnover along gradients. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131201. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 1 May 2023).
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Suárez-Castro, A.F.; Raymundo, M.; Bimler, M.; Mayfield, M.M. Using multi-scale spatially explicit frameworks to understand the relationship between functional diversity and species richness. Ecography 2022, 2022, e05844. [Google Scholar] [CrossRef]
- Ibarra, J.T.; Martin, K. Biotic homogenization: Loss of avian functional richness and habitat specialists in disturbed Andean temperate forests. Biol. Conserv. 2015, 192, 418–427. [Google Scholar] [CrossRef]
- Cooke, R.S.; Bates, A.E.; Eigenbrod, F. Global trade-offs of functional redundancy and functional dispersion for birds and mammals. Glob. Ecol. Biogeogr. 2019, 28, 484–495. [Google Scholar] [CrossRef]
- Lanari, M.; Horta, P.A.; da Silva Copertino, M. Functional redundancy and stability in a subtidal macroalgal community in the Southwestern Atlantic coast. Mar. Environ. Res. 2022, 173, 105519. [Google Scholar] [CrossRef]
- Cumming, G.S.; Child, M.F. Contrasting spatial patterns of taxonomic and functional richness offer insights into potential loss of ecosystem services. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Luck, G.W.; Carter, A.; Smallbone, L. Changes in bird functional diversity across multiple land uses: Interpretations of functional redundancy depend on functional group identity. PLoS ONE 2013, 8, e63671. [Google Scholar] [CrossRef]
- Pecuchet, L.; Törnroos, A.; Lindegren, M. Patterns and drivers of fish community assembly in a large marine ecosystem. Mar. Ecol. Prog. Ser. 2016, 546, 239–248. [Google Scholar] [CrossRef]
- Guillemot, N.; Kulbicki, M.; Chabanet, P.; Vigliola, L. Functional redundancy patterns reveal non-random assembly rules in a species-rich marine assemblage. PLoS ONE 2011, 6, e26735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zang, R. Tropical forests are vulnerable in terms of functional redundancy. Biol. Conserv. 2021, 262, 109326. [Google Scholar] [CrossRef]
- Zhang, S.; Zang, R.; Sheil, D. Rare and common species contribute disproportionately to the functional variation within tropical forests. J. Environ. Manag. 2022, 304, 114332. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, N.S.; Brook, B.W.; Bradshaw, C.J. Causes and consequences of species extinctions. Princet. Guide Ecol. 2009, 1, 514–520. [Google Scholar]
- Violle, C.; Thuiller, W.; Mouquet, N.; Munoz, F.; Kraft, N.J.; Cadotte, M.W.; Livingstone, S.W.; Mouillot, D. Functional rarity: The ecology of outliers. Trends Ecol. Evol. 2017, 32, 356–367. [Google Scholar] [CrossRef]
- Winfree, R.; Reilly, J.R.; Bartomeus, I.; Cariveau, D.P.; Williams, N.M.; Gibbs, J. Species turnover promotes the importance of bee diversity for crop pollination at regional scales. Science 2018, 359, 791–793. [Google Scholar] [CrossRef]
- Genung, M.A.; Reilly, J.; Williams, N.M.; Buderi, A.; Gardner, J.; Winfree, R. Rare and declining bee species are key to consistent pollination of wildflowers and crops across large spatial scales. Ecology 2023, 104, e3899. [Google Scholar] [CrossRef]
- Turner, J.R.; Gatehouse, C.M.; Corey, C.A. Does solar energy control organic diversity? Butterflies, moths and the British climate. Oikos 1987, 48, 195–205. [Google Scholar] [CrossRef]
- Menéndez, R.; González-Megías, A.; Collingham, Y.; Fox, R.; Roy, D.B.; Ohlemüller, R.; Thomas, C.D. Direct and indirect effects of climate and habitat factors on butterfly diversity. Ecology 2007, 88, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Warren, M.; Hill, J.; Thomas, J.; Asher, J.; Fox, R.; Huntley, B.; Roy, D.; Telfer, M.; Jeffcoate, S.; Harding, P. Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 2001, 414, 65–69. [Google Scholar] [CrossRef]
- Fox, R.; Oliver, T.H.; Harrower, C.; Parsons, M.S.; Thomas, C.D.; Roy, D.B. Long-term changes to the frequency of occurrence of British moths are consistent with opposing and synergistic effects of climate and land-use changes. J. Appl. Ecol. 2014, 51, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Ridding, L.E.; Redhead, J.W.; Pywell, R.F. Fate of semi-natural grassland in England between 1960 and 2013: A test of national conservation policy. Glob. Ecol. Conserv. 2015, 4, 516–525. [Google Scholar] [CrossRef]
- Fox, R.D.E.; Purdy, K.M.; Middlebrook, I.; Roy, D.B.; Noble, D.G.; Botham, M.S.; Bourn, N.A.D. The State of the UK’s Butterflies 2022; Butterfly Conservation: Wareham, UK, 2023. [Google Scholar]
- Dapporto, L.; Dennis, R.L. The generalist–specialist continuum: Testing predictions for distribution and trends in British butterflies. Biol. Conserv. 2013, 157, 229–236. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Chang. Biol. 2000, 4, 407–416. [Google Scholar] [CrossRef]
- Diamond, S.E.; Frame, A.M.; Martin, R.A.; Buckley, L.B. Species’ traits predict phenological responses to climate change in butterflies. Ecology 2011, 92, 1005–1012. [Google Scholar] [CrossRef]
- Forrest, J.R. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate change effects on animal ecology: Butterflies and moths as a case study. Biol. Rev. 2021, 96, 2113–2126. [Google Scholar] [CrossRef]
- Mason, S.C.; Palmer, G.; Fox, R.; Gillings, S.; Hill, J.K.; Thomas, C.D.; Oliver, T.H. Geographical range margins of many taxonomic groups continue to shift polewards. Biol. J. Linn. Soc. 2015, 115, 586–597. [Google Scholar] [CrossRef]
- Menéndez, R.; Megías, A.G.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species richness changes lag behind climate change. Proc. R. Soc. B Biol. Sci. 2006, 273, 1465–1470. [Google Scholar] [CrossRef]
- Slancarova, J.; Benes, J.; Kristynek, M.; Kepka, P.; Konvicka, M. Does the surrounding landscape heterogeneity affect the butterflies of insular grassland reserves? A contrast between composition and configuration. J. Insect Conserv. 2014, 18, 1–12. [Google Scholar] [CrossRef]
- Oliver, T.; Roy, D.B.; Hill, J.K.; Brereton, T.; Thomas, C.D. Heterogeneous landscapes promote population stability. Ecol. Lett. 2010, 13, 473–484. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Shreeve, T.G.; Dennis, R.L.H.; Roy, D.B.; Moss, D. An ecological classification of British butterflies: Ecological attributes and biotope occupancy. J. Insect Conserv. 2001, 5, 145–161. [Google Scholar] [CrossRef]
- Laliberte, E.; Wells, J.A.; DeClerck, F.; Metcalfe, D.J.; Catterall, C.P.; Queiroz, C.; Aubin, I.; Bonser, S.P.; Ding, Y.; Fraterrigo, J.M. Land-use intensification reduces functional redundancy and response diversity in plant communities. Ecol. Lett. 2010, 13, 76–86. [Google Scholar] [CrossRef]
- Carmona, C.P.; Guerrero, I.; Peco, B.; Morales, M.B.; Oñate, J.J.; Pärt, T.; Tscharntke, T.; Liira, J.; Aavik, T.; Emmerson, M. Agriculture intensification reduces plant taxonomic and functional diversity across European arable systems. Funct. Ecol. 2020, 34, 1448–1460. [Google Scholar] [CrossRef]
- Zellweger, F.; Roth, T.; Bugmann, H.; Bollmann, K. Beta diversity of plants, birds and butterflies is closely associated with climate and habitat structure. Glob. Ecol. Biogeogr. 2017, 26, 898–906. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, K.; Messyasz, B.; Xu, Y.; Gao, M.; Li, Y.; Wu, N. Functional and taxonomic beta diversity of butterfly assemblages in an archipelago: Relative importance of island characteristics, climate, and spatial factors. Ecol. Indic. 2022, 142, 109191. [Google Scholar] [CrossRef]
- Villéger, S.; Grenouillet, G.; Brosse, S. Decomposing functional β-diversity reveals that low functional β-diversity is driven by low functional turnover in E uropean fish assemblages. Glob. Ecol. Biogeogr. 2013, 22, 671–681. [Google Scholar] [CrossRef]
- Leao-Pires, T.A.; Luiz, A.M.; Sawaya, R.J. The complex roles of space and environment in structuring functional, taxonomic and phylogenetic beta diversity of frogs in the Atlantic Forest. PLoS ONE 2018, 13, e0196066. [Google Scholar] [CrossRef] [PubMed]
- Sandal, L.; Grøtan, V.; Sæther, B.E.; Freckleton, R.P.; Noble, D.G.; Ovaskainen, O. Effects of density, species interactions, and environmental stochasticity on the dynamics of British bird communities. Ecology 2022, 103, e3731. [Google Scholar] [CrossRef]
- Lazarina, M.; Sgardelis, S.P.; Michailidou, D.E.; Tsianou, M.; Andrikou-Charitidou, A.; Touloumis, K.; Kallimanis, A.S. Replacement drives native β-diversity of British avifauna, while richness differences shape alien β-diversity. Divers. Distrib. 2023, 29, 61–74. [Google Scholar] [CrossRef]
- Lewthwaite, J.M.; Debinski, D.M.; Kerr, J.T. High community turnover and dispersal limitation relative to rapid climate change. Glob. Ecol. Biogeogr. 2017, 26, 459–471. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Bini, L.M.; Altermatt, F.; Al-Shami, S.A.; Angeler, D.G.; Bonada, N.; Brand, C.; Callisto, M.; Cottenie, K. A comparative analysis reveals weak relationships between ecological factors and beta diversity of stream insect metacommunities at two spatial levels. Ecol. Evol. 2015, 5, 1235–1248. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Malumbres-Olarte, J.; Arnedo, M.A.; Crespo, L.C.; Domenech, M.; Cardoso, P. Taxonomic divergence and functional convergence in Iberian spider forest communities: Insights from beta diversity partitioning. J. Biogeogr. 2020, 47, 288–300. [Google Scholar] [CrossRef]
- Wayman, J.P.; Sadler, J.P.; Pugh, T.A.; Martin, T.E.; Tobias, J.A.; Matthews, T.J. Identifying the drivers of spatial taxonomic and functional beta-diversity of British breeding birds. Front. Ecol. Evol. 2021, 9, 620062. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic β-Diversity | Functional β-Diversity | |
|---|---|---|
| Deviance Explained | 49.13% | 25.40% |
| Variable Importance | ||
| Geographic Distance | 0.76 | 0.54 |
| Temperature | 1.13 | 1.59 |
| Cropland | 0.02 | 0.01 |
| Forest | 0.18 | 0.20 |
| Other | 0.01 | 0.18 |
| Settlement | 0.01 | 0.02 |
| Water | 0.00 | 0.02 |
| Human Population | 0.23 | 0.31 |
| Elevation | 0.00 | 0.00 |
| Elevation Variability | 0.43 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarina, M.; Michailidou, D.-E.; Tsianou, M.; Kallimanis, A.S. How Biodiversity, Climate and Landscape Drive Functional Redundancy of British Butterflies. Insects 2023, 14, 722. https://doi.org/10.3390/insects14090722
Lazarina M, Michailidou D-E, Tsianou M, Kallimanis AS. How Biodiversity, Climate and Landscape Drive Functional Redundancy of British Butterflies. Insects. 2023; 14(9):722. https://doi.org/10.3390/insects14090722
Chicago/Turabian StyleLazarina, Maria, Danai-Eleni Michailidou, Mariana Tsianou, and Athanasios S. Kallimanis. 2023. "How Biodiversity, Climate and Landscape Drive Functional Redundancy of British Butterflies" Insects 14, no. 9: 722. https://doi.org/10.3390/insects14090722
APA StyleLazarina, M., Michailidou, D. -E., Tsianou, M., & Kallimanis, A. S. (2023). How Biodiversity, Climate and Landscape Drive Functional Redundancy of British Butterflies. Insects, 14(9), 722. https://doi.org/10.3390/insects14090722