
Effects of Temperature on Blood Feeding and Activity Levels in the Tiger Mosquito, Aedes albopictus

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Propensity to Blood Feed and Blood Meal Size across Different Temperatures
2.2. Blood Meal Size
2.3. Propensity to Take a Blood Meal
2.4. Mosquito Activity Levels across Different Temperatures
3. Results
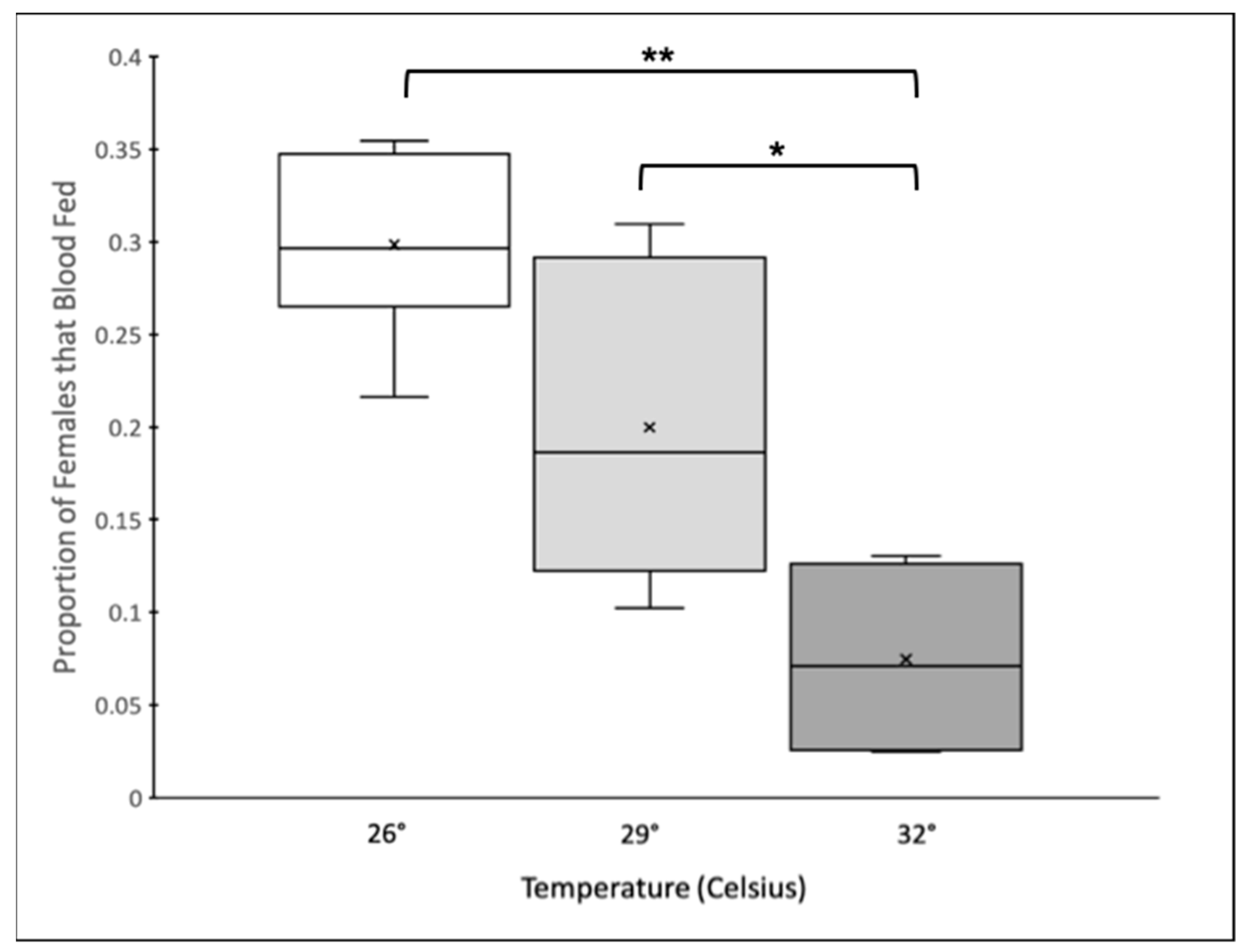
3.1. Propensity to Blood Feed and Blood Meal Size across Different Temperatures
3.2. Mosquito Activity Levels across Different Temperatures
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Gutiérrez, D.; Wilson, R.J. Intra-and Interspecific Variation in the Responses of Insect Phenology to Climate. J. Anim. Ecol. 2021, 90, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, J.; Huey, R. Size, Temperature, and Fitness: Three Rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Forrest, J.R.K. Complex Responses of Insect Phenology to Climate Change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Boukal, D.S.; Bideault, A.; Carreira, B.M.; Sentis, A. Species Interactions under Climate Change: Connecting Kinetic Effects of Temperature on Individuals to Community Dynamics. Curr. Opin. Insect Sci. 2019, 35, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The Effect of Temperature on Life History Traits of Culex Mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Padmanabha, H.; Correa, F.; Legros, M.; Nijhout, H.F.; Lord, C.; Lounibos, L.P. An Eco-Physiological Model of the Impact of Temperature on Aedes aegypti Life History Traits. J. Insect Physiol. 2012, 58, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Macoris, M.; Galvani, K.C.; Andrighetti, M.T.M.; Wanderley, D.M.V. Assessing the Effects of Temperature on the Population of Aedes aegypti, the Vector of Dengue. Epidemiol. Infect. 2009, 137, 1188–1202. [Google Scholar] [CrossRef]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects 2018, 9, 158. [Google Scholar] [CrossRef]
- Teng, H.-J.; Apperson, C.S. Development and Survival of Immature Aedes albopictus and Aedes triseriatus (Diptera: Culicidae) in the Laboratory: Effects of Density, Food, and Competition on Response to Temperature. J. Med. Entomol. 2000, 37, 40–52. [Google Scholar] [CrossRef]
- Grigaltchik, V.S.; Webb, C.; Seebacher, F. Temperature Modulates the Effects of Predation and Competition on Mosquito Larvae. Ecol. Entomol. 2016, 41, 668–675. [Google Scholar] [CrossRef]
- Muturi, E.J.; Blackshear, M., Jr.; Montgomery, A. Temperature and Density-Dependent Effects of Larval Environment on Aedes aegypti Competence for an Alphavirus. J. Vector Ecol. 2012, 37, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Bettinardi, D. Temperature and Dengue Virus Infection in Mosquitoes: Independent Effects on the Immature and Adult Stages. Am. J. Trop. Med. Hyg. 2013, 88, 497. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.L.M.; Whitehead, S.A.; Thomas, M.B. Quantifying the Effects of Temperature on Mosquito and Parasite Traits That Determine the Transmission Potential of Human Malaria. PLoS Biol. 2017, 15, e2003489. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.R.; Ryan, S.J.; Savage, V.; Shocket, M.S.; et al. Thermal Biology of Mosquito-Borne Disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The Global Distribution of the Arbovirus Vectors Aedes aegypti and Ae. Albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Lounibos, L.P. Invasions by Insect Vectors of Human Disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A.; Philip Lounibos, L. Ecology of Invasive Mosquitoes: Effects on Resident Species and on Human Health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef] [PubMed]
- Bargielowski, I.E.; Lounibos, L.P. Satyrization and Satyrization-Resistance in Competitive Displacements of Invasive Mosquito Species. Insect Sci. 2016, 23, 162–174. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical Review of the Vector Status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Pereira-dos-Santos, T.; Roiz, D.; Lourenço-de-Oliveira, R.; Paupy, C. A Systematic Review: Is Aedes albopictus an Efficient Bridge Vector for Zoonotic Arboviruses? Pathogens 2020, 9, 266. [Google Scholar] [CrossRef]
- Vanlandingham, D.L.; Higgs, S.; Huang, Y.-J.S. Aedes albopictus (Diptera: Culicidae) and Mosquito-Borne Viruses in the United States. J. Med. Entomol. 2016, 53, 1024–1028. [Google Scholar] [CrossRef]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of Temperature on Immature Development, Survival, Longevity, Fecundity, and Gonotrophic Cycles of Aedes albopictus, Vector of Chikungunya and Dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Juliano, S.A. Temperature Effects on the Dynamics of Aedes albopictus (Diptera: Culicidae) Populations in the Laboratory. J. Med. Entomol. 2001, 38, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling Adult Aedes aegypti and Aedes albopictus Survival at Different Temperatures in Laboratory and Field Settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate Change and Range Expansion of the Asian Tiger Mosquito (Aedes albopictus) in Northeastern USA: Implications for Public Health Practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; L’ambert, G.; Lacour, G.; Benoît, R.; Demarchi, M.; Cros, M.; Cailly, P.; Aubry-Kientz, M.; Balenghien, T.; Ezanno, P. A Rainfall-and Temperature-Driven Abundance Model for Aedes albopictus Populations. Int. J. Environ. Res. Public Health 2013, 10, 1698–1719. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global Expansion and Redistribution of Aedes-Borne Virus Transmission Risk with Climate Change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef]
- Xiao, F.-Z.; Zhang, Y.; Deng, Y.-Q.; He, S.; Xie, H.-G.; Zhou, X.-N.; Yan, Y.-S. The Effect of Temperature on the Extrinsic Incubation Period and Infection Rate of Dengue Virus Serotype 2 Infection in Aedes albopictus. Arch. Virol. 2014, 159, 3053–3057. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Chen, T.-H.; Lin, C.; Shu, P.-Y.; Su, C.-L.; Teng, H.-J. The Impact of Temperature and Wolbachia Infection on Vector Competence of Potential Dengue Vectors Aedes aegypti and Aedes albopictus in the Transmission of Dengue Virus Serotype 1 in Southern Taiwan. Parasites Vectors 2017, 10, 551. [Google Scholar] [CrossRef]
- Fikrig, K.; Harrington, L.C. Understanding and Interpreting Mosquito Blood Feeding Studies: The Case of Aedes albopictus. Trends Parasitol. 2021, 37, 959–975. [Google Scholar] [CrossRef]
- Onyango, M.G.; Bialosuknia, S.M.; Payne, A.F.; Mathias, N.; Kuo, L.; Vigneron, A.; DeGennaro, M.; Ciota, A.T.; Kramer, L.D. Increased Temperatures Reduce the Vectorial Capacity of Aedes Mosquitoes for Zika Virus. Emerg. Microbes Infect. 2020, 9, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Dye, C. Vectorial Capacity: Must We Measure All Its Components? Parasitol. Today 1986, 2, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ciota, A.T. Dissecting Vectorial Capacity for Mosquito-Borne Viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef]
- Klowden, M.J.; Lea, A.O. Blood Meal Size as a Factor Affecting Continued Host-Seeking by Aedes aegypti (L.). Am. J. Trop. Med. Hyg. 1978, 27, 827–831. [Google Scholar] [CrossRef]
- May, M.L. Insect Thermoregulation. Annu. Rev. Entomol. 1979, 24, 313–349. [Google Scholar] [CrossRef]
- Kearney, M.; Shine, R.; Porter, W.P. The Potential for Behavioral Thermoregulation to Buffer “Cold-Blooded” Animals against Climate Warming. Proc. Natl. Acad. Sci. USA 2009, 106, 3835–3840. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, R.; Blanckenhorn, W.U.; Mathis, A.; Verhulst, N.O. Video Analysis of the Locomotory Behaviour of Aedes aegypti and Ae. japonicus Mosquitoes under Different Temperature Regimes in a Laboratory Setting. J. Therm. Biol. 2022, 105, 103205. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lazzari, C.R.; Denlinger, D.L.; Lahondère, C. Thermoprotective Adaptations Are Critical for Arthropods Feeding on Warm-Blooded Hosts. Curr. Opin. Insect Sci. 2019, 34, 7–11. [Google Scholar] [CrossRef]
- Lahondère, C.; Lazzari, C.R. Mosquitoes Cool down during Blood Feeding to Avoid Overheating. Curr. Biol. 2012, 22, 40–45. [Google Scholar] [CrossRef]
- de Freitas, R.M. A Review on the Ecological Determinants of Aedes aegypti (Diptera: Culicidae) Vectorial Capacity. Oecologia Aust. 2010, 14, 726–736. [Google Scholar] [CrossRef]
- Briegel, H. Metabolic Relationship between Female Body Size, Reserves, and Fecundity of Aedes aegypti. J. Insect Physiol. 1990, 36, 165–172. [Google Scholar] [CrossRef]
- Briegel, H. Mosquito Reproduction: Incomplete Utilization of the Blood Meal Protein for Oögenesis. J. Insect Physiol. 1985, 31, 15–21. [Google Scholar] [CrossRef]
- Foster, W.A. Mosquito Sugar Feeding and Reproductive Energetics. Annu. Rev. Entomol. 1995, 40, 443–474. [Google Scholar] [CrossRef]
- Timmermann, S.E.; Briegel, H. Larval Growth and Biosynthesis of Reserves in Mosquitoes. J. Insect Physiol. 1999, 45, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Briegel, H.; Timmermann, S.E. Aedes albopictus (Diptera: Culicidae): Physiological Aspects of Development and Reproduction. J. Med. Entomol. 2001, 38, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Reiskind, M.H.; Zarrabi, A.A. Is Bigger Really Bigger? Differential Responses to Temperature in Measures of Body Size of the Mosquito, Aedes albopictus. J. Insect Physiol. 2012, 58, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Klowden, M.J.; Blackmer, J.L.; Chambers, G.M. Effects of Larval Nutrition on the Host-Seeking Behavior of Adult Aedes aegypti Mosquitoes. J. Am. Mosq. Control Assoc. 1988, 4, 73–75. [Google Scholar]
- Van Handel, E. The Obese Mosquito. J. Physiol. 1965, 181, 478. [Google Scholar] [CrossRef]
- Nasci, R.S. Influence of Larval and Adult Nutrition on Biting Persistence in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 1991, 28, 522–526. [Google Scholar] [CrossRef]
- Terlau, J.F.; Brose, U.; Eisenhauer, N.; Amyntas, A.; Boy, T.; Dyer, A.; Gebler, A.; Hof, C.; Liu, T.; Scherber, C.; et al. Microhabitat Conditions Remedy Heat Stress Effects on Insect Activity. Glob. Chang. Biol. 2023, 29, 3747–3758. [Google Scholar] [CrossRef]
- Kamimura, K.; Matsuse, I.T.; Takahashi, H.; Komukai, J.; Fukuda, T.; Suzuki, K.; Aratani, M.; Shirai, Y.; Mogi, M. Effect of Temperature on the Development of Aedes aegypti and Aedes albopictus. Med. Entomol. Zool. 2002, 53, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.M.; Shaw, R.; Lahondère, C. Beat the Heat: Culex quinquefasciatus Regulates Its Body Temperature during Blood Feeding. J. Therm. Biol. 2021, 96, 102826. [Google Scholar] [CrossRef] [PubMed]
- Vail, S.G.; Smith, G. Vertical Movement and Posture of Blacklegged Tick (Acari: Ixodidae) Nymphs as a Function of Temperature and Relative Humidity in Laboratory Experiments. J. Med. Entomol. 2002, 39, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Zhu, W.; Fu, Y.; Zhou, S. Influence of High-Temperature Exposure on the Mating, Oviposition and Thermotaxis of Bactrocera cucurbitae (Coquillet)(Diptera: Tephritidae). PLoS ONE 2018, 13, e0204065. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Mulla, M.S. Effects of Temperature on Development, Mortality, Mating and Blood Feeding Behavior of Culiseta incidens (Diptera: Culicidae). J. Vector Ecol. 2001, 26, 83–92. [Google Scholar]
- Gunathilaka, N.; Ranathunge, T.; Udayanga, L.; Abeyewickreme, W. Efficacy of Blood Sources and Artificial Blood Feeding Methods in Rearing of Aedes aegypti (Diptera: Culicidae) for Sterile Insect Technique and Incompatible Insect Technique Approaches in Sri Lanka. BioMed Res. Int. 2017, 2017, 3196924. [Google Scholar] [CrossRef] [PubMed]
- Seck, F.; Cailleau, A.; Diallo, M.; Dia, I. Comparison of the Efficiency and Performance of Two Systems and Three Membranes for Blood Feeding Mosquitoes. BMC Res. Notes 2021, 14, 388. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T. Multi-Cue Integration: How Female Mosquitoes Locate a Human Host. Curr. Biol. 2015, 25, R793–R795. [Google Scholar] [CrossRef]
- Corfas, R.A.; Vosshall, L.B. The Cation Channel TRPA1 Tunes Mosquito Thermotaxis to Host Temperatures. eLife 2015, 4, e11750. [Google Scholar] [CrossRef]
- Clements, A.N. The Biology of Mosquitoes. Volume 2: Sensory Reception and Behaviour; CABI Publishing: Wallingford, UK, 1999. [Google Scholar]
- Boyer, S.; Maillot, L.; Gouagna, L.C.; Fontenille, D.; Chadee, D.D.; Lempérière, G. Diel Activity Patterns of Male Aedes albopictus in the Laboratory. J. Am. Mosq. Control Assoc. 2013, 29, 74–77. [Google Scholar] [CrossRef]
- Stevenson, R.D. The Relative Importance of Behavioral and Physiological Adjustments Controlling Body Temperature in Terrestrial Ectotherms. Am. Nat. 1985, 126, 362–386. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-Safety Margins and the Necessity of Thermoregulatory Behavior across Latitude and Elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Ma, C.-S. Effect of Acclimation on Heat-Escape Temperatures of Two Aphid Species: Implications for Estimating Behavioral Response of Insects to Climate Warming. J. Insect Physiol. 2012, 58, 303–309. [Google Scholar] [CrossRef]
- Bodlah, M.A.; Iqbal, J.; Ashiq, A.; Bodlah, I.; Jiang, S.; Mudassir, M.A.; Rasheed, M.T.; Fareen, A.G.E. Insect Behavioral Restraint and Adaptation Strategies under Heat Stress: An Inclusive Review. J. Saudi Soc. Agric. Sci. 2023, in press. [Google Scholar] [CrossRef]
- Pincebourde, S.; Suppo, C. The Vulnerability of Tropical Ectotherms to Warming Is Modulated by the Microclimatic Heterogeneity. Integr. Comp. Biol. 2016, 56, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.T.; Beckerman, A.P.; Schmitz, O.J. Climate Warming Strengthens Indirect Interactions in an Old-Field Food Web. Ecology 2009, 90, 2346–2351. [Google Scholar] [CrossRef]
- Sauer, F.G.; Kiel, E.; Lühken, R. Effects of Mosquito Resting Site Temperatures on the Estimation of Pathogen Development Rates in Near-Natural Habitats in Germany. Parasites Vectors 2022, 15, 390. [Google Scholar] [CrossRef]
- Sauer, F.G.; Grave, J.; Lühken, R.; Kiel, E. Habitat and Microclimate Affect the Resting Site Selection of Mosquitoes. Med. Vet. Entomol. 2021, 35, 379–388. [Google Scholar] [CrossRef]
- Samson, D.M.; Qualls, W.A.; Roque, D.; Naranjo, D.P.; Alimi, T.; Arheart, K.L.; Müller, G.C.; Beier, J.C.; Xue, R.-D. Resting and Energy Reserves of Aedes albopictus Collected in Common Landscaping Vegetation in St. Augustine, Florida. J. Am. Mosq. Control Assoc. 2013, 29, 231–236. [Google Scholar] [CrossRef]
- Andrew, N.R.; Hart, R.A.; Jung, M.-P.; Hemmings, Z.; Terblanche, J.S. Can Temperate Insects Take the Heat? A Case Study of the Physiological and Behavioural Responses in a Common Ant, Iridomyrmex purpureus (Formicidae), with Potential Climate Change. J. Insect Physiol. 2013, 59, 870–880. [Google Scholar] [CrossRef]
- Ma, G.; Ma, C.S. Upper Critical Temperatures for Behaviors of Three Species of Cereal Aphids in Leaf Temperature Gradients. Acta Ecol. Sin. 2007, 27, 2449–2459. [Google Scholar]
- Ziegler, R.; Blanckenhorn, W.U.; Mathis, A.; Verhulst, N.O. Temperature Preference of Sugar-or Blood-Fed Aedes japonicus Mosquitoes under Semi-Natural Conditions. J. Therm. Biol. 2023, 114, 103592. [Google Scholar] [CrossRef] [PubMed]
- Blanford, S.; Read, A.F.; Thomas, M.B. Thermal Behaviour of Anopheles Stephensi in Response to Infection with Malaria and Fungal Entomopathogens. Malar. J. 2009, 8, 72. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, K.; Occhino, D. Effects of Temperature on Blood Feeding and Activity Levels in the Tiger Mosquito, Aedes albopictus. Insects 2023, 14, 752. https://doi.org/10.3390/insects14090752
Costanzo K, Occhino D. Effects of Temperature on Blood Feeding and Activity Levels in the Tiger Mosquito, Aedes albopictus. Insects. 2023; 14(9):752. https://doi.org/10.3390/insects14090752
Chicago/Turabian StyleCostanzo, Katie, and Dominic Occhino. 2023. "Effects of Temperature on Blood Feeding and Activity Levels in the Tiger Mosquito, Aedes albopictus" Insects 14, no. 9: 752. https://doi.org/10.3390/insects14090752
APA StyleCostanzo, K., & Occhino, D. (2023). Effects of Temperature on Blood Feeding and Activity Levels in the Tiger Mosquito, Aedes albopictus. Insects, 14(9), 752. https://doi.org/10.3390/insects14090752