

Anatolian Short-Horned Grasshoppers Unveiled: Integrating Biogeography and Pest Potential


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
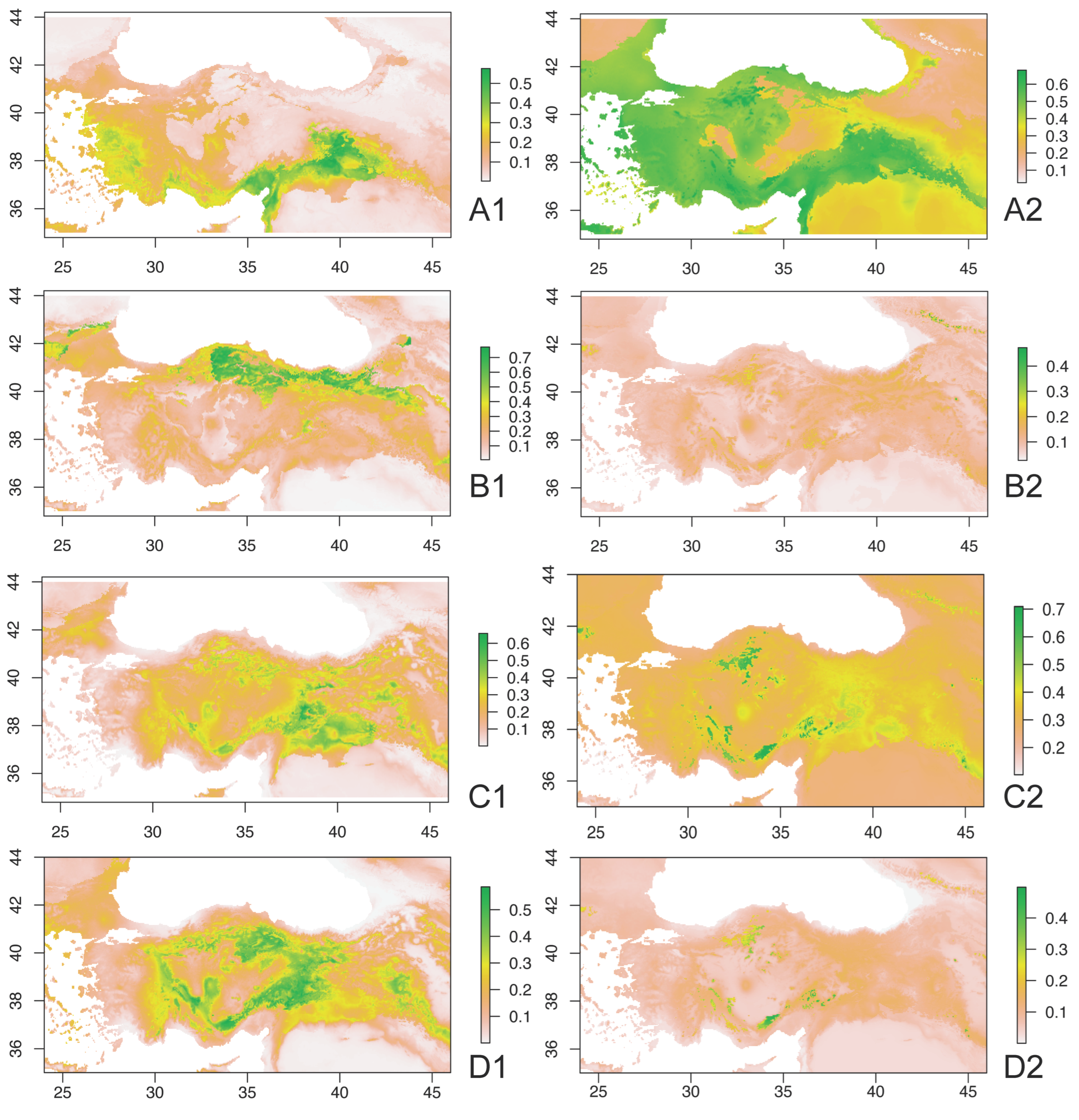
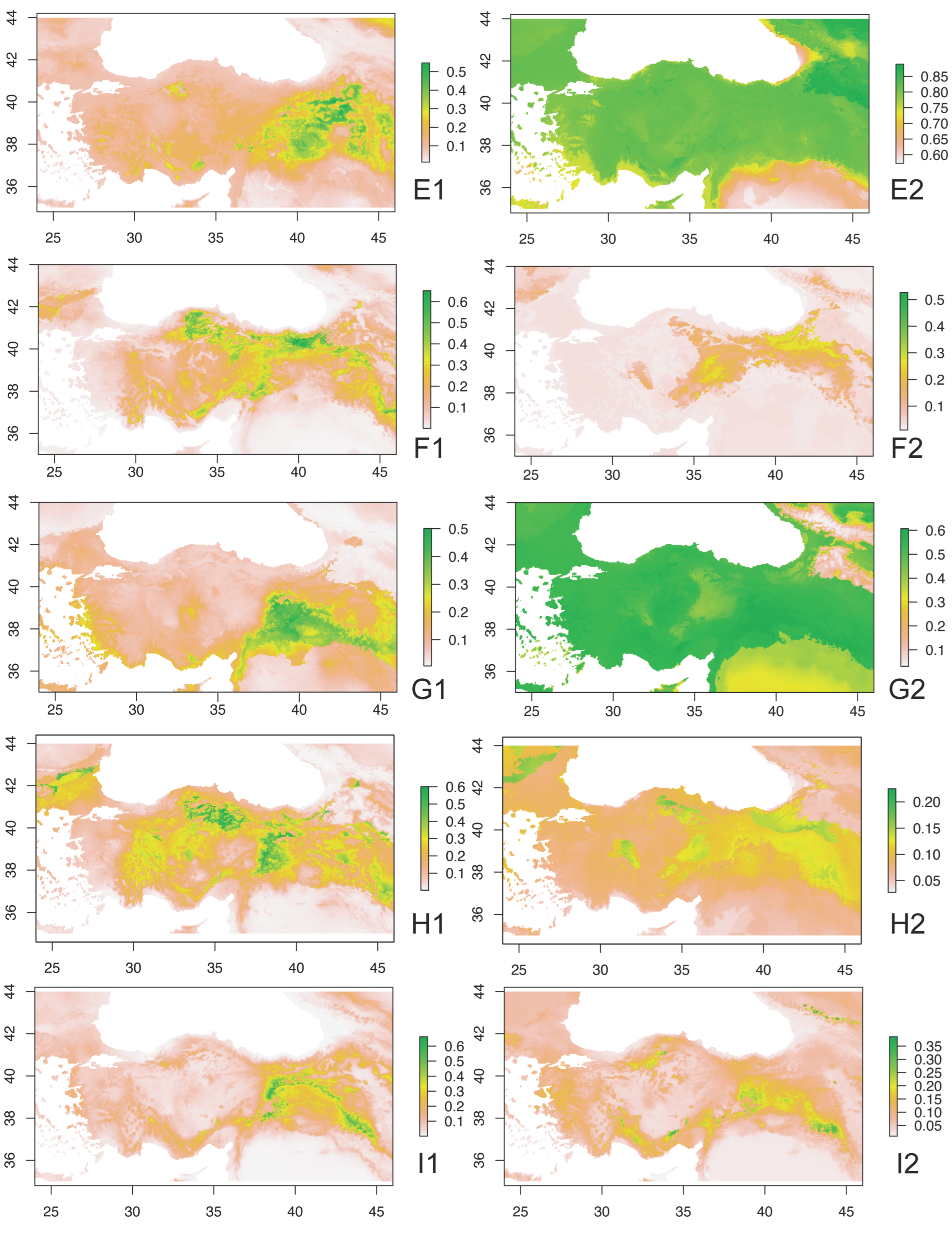
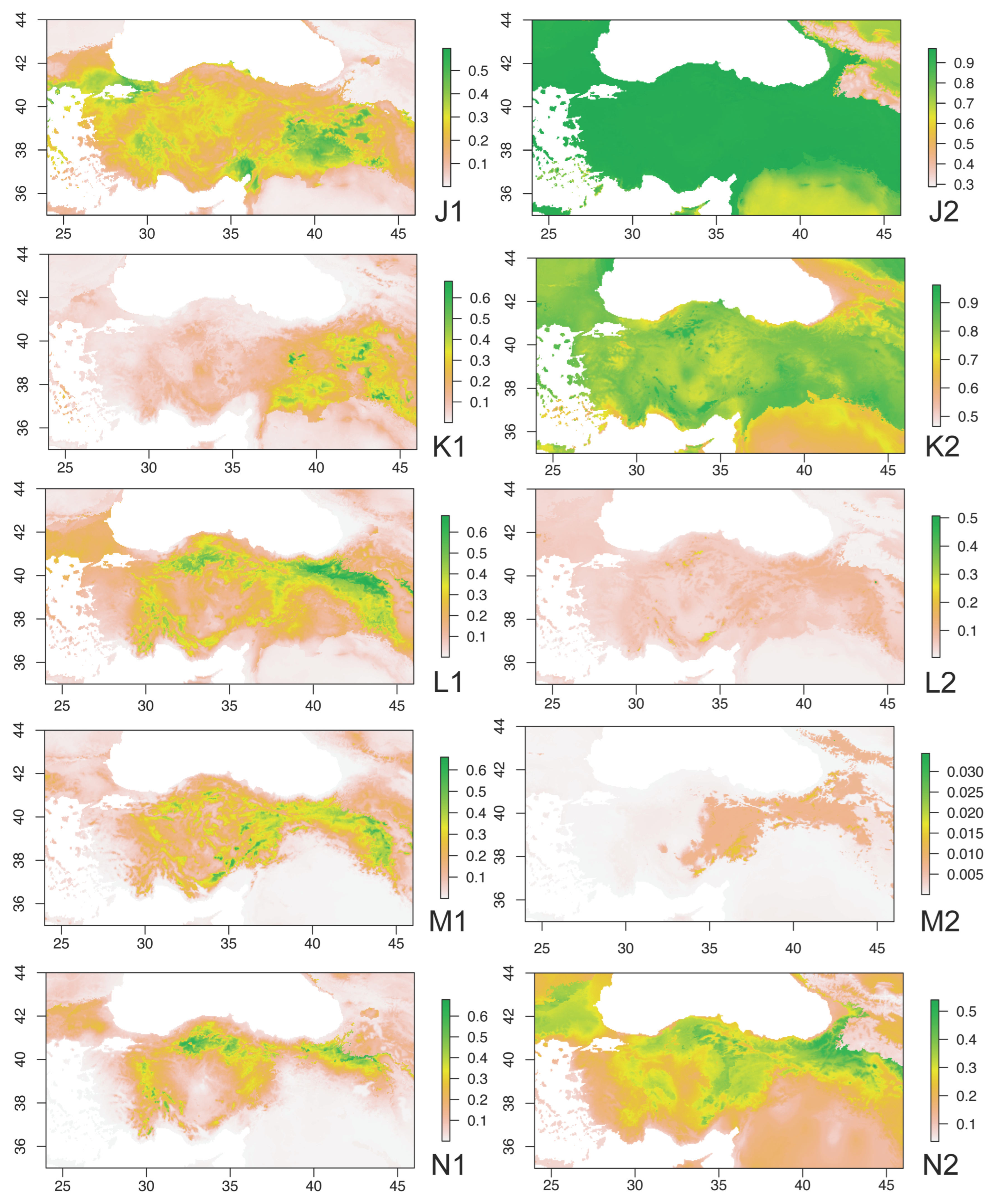
3. Results
4. Discussion and Conclusions
4.1. Faunal Composition of Anatolian Caelifera
4.2. Ecobiogeographic Classification of Anatolian Caelifera
4.3. Pest and Pullulating Species of Caelifera in Anatolia
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | ME | ES | IA | MP | EN | |
|---|---|---|---|---|---|---|
| Family Tridactylidae Brullé, 1835 | ||||||
| Subfamily Dentridactylinae Günther, 1979 | ||||||
| Bruntridactylus Günther, 1979 | ||||||
| 1 | B. irremipes (Uvarov, 1934) | * | ||||
| Subfamily Tridactylinae Brullé, 1835 | ||||||
| Asiotridactylus Günther, 1995 | ||||||
| 2 | A. fasciatus (Guérin-Méneville, 1844) | * | ||||
| Xya Latreille, 1809 | ||||||
| 3 | X. variegata (Latreille, 1809) | * | * | * | ||
| 4 | X. pfaendleri Harz, 1970 | * | ||||
| Family Tetrigidae Rambur, 1838 | ||||||
| Subfamily Tetriginae Rambur, 1838 | ||||||
| Tetrix Latreille, 1802 | ||||||
| 5 | T. depressa Brisout de Barneville, 1848 | * | * | * | * | |
| 6 | T. tenuicornis tenuicornis (Sahlberg, 1891) | * | * | |||
| 7 | T. bolivari Saulcy, 1901 | * | * | * | * | |
| 8 | T. ceperoi ceperoi (Bolívar, 1887) | * | ||||
| 9 | T. subulata (Linnaeus, 1758) | * | * | * | * | |
| 10 | T. tuerki (Krauss, 1876) | * | ||||
| Paratettix Bolivar, 1887 | ||||||
| 11 | P. meridionalis (Rambur, 1838) | * | * | * | ||
| 12 | P. iranica Uvarov, 1952 | * | ||||
| Family Pyrgomorphidae Brunner, 1874 | ||||||
| Subfamily Pyrgomorphinae Brunner, 1874 | ||||||
| Pyrgomorpha Serville, 1838 | ||||||
| 13 | Pyrgomorpha (P.) conica (Olivier, 1791) | * | * | |||
| 14 | Pyrgomorpha (P.) cognata Krauss, 1877 | * | * | |||
| 15 | Pyrgomorpha (P.) guentheri Burr, 1899 | * | * | * | ||
| Family Pamphagidae Burmeister, 1840 | ||||||
| Subfamily Thrinchinae Stål, 1876 | ||||||
| Eremopeza Saussure, 1888 | ||||||
| 16 | E. gibbera gibbera (Stål, 1876) | * | * | * | ||
| 17 | E. gibbera lata (Uvarov, 1934) | * | ||||
| 18 | E. festiva (Bolivar, 1884) | * | ||||
| 19 | E. saussurei saussurei (Uvarov, 1918) | * | ||||
| Prionotropis Fieber, 1853 | ||||||
| 20 | P. maculinervis (Stal, 1876) | * | * | * | ||
| Tmethis Fieber, 1853 | ||||||
| 21 | T. pulchripennis asiaticus Uvarov, 1943 | * | ||||
| Asiotmethis Uvarov, 1943 | ||||||
| 22 | A. limbatus (Charpentier, 1845) | * | ||||
| 23 | A. turritus (Fischer, 1833) | * | ||||
| Glyphotmethis Bei-Bienko, 1951 | ||||||
| 24 | G. holtzi brachypterus Ünal, 2007 | * | ||||
| 25 | G. holtzi holtzi (Werner, 1901) | * | * | * | ||
| 26 | G. holtzi pulchripes (Uvarov, 1943) | * | * | |||
| 27 | G. holtzi turcicus Ünal, 2007 | * | * | |||
| 28 | G. dimorphus dimorphus (Uvarov, 1934) | * | * | * | ||
| 29 | G. dimorphus armenus (Ramme, 1951) | * | * | |||
| 30 | G. ovipennis (Uvarov, 1934) | * | * | * | ||
| 31 | G. efe Ünal, 2007 | * | * | |||
| 32 | G. escherichi escherichi (Krauss, 1896) | * | * | * | * | |
| 33 | G. escherichi eliator (Ramme, 1951) | * | * | * | ||
| 34 | G. sevketi (Ramme, 1951) | * | * | |||
| 35 | G. escherichi inermis (Uvarov, 1934) | * | * | |||
| 36 | G. adaliae (Uvarov, 1928) | * | * | |||
| Subfamily Pamphaginae Burmeister, 1840 | ||||||
| Ebnerodes Ramme, 1951 | ||||||
| 37 | E. toelgi (Ebner, 1919) | * | * | |||
| Paranocarodes Bolivar, 1916 | ||||||
| 38 | P. anatoliensis anamas Ünal, 2016 | * | * | |||
| 39 | P. anatoliensis anatoliensis Demirsoy, 1973 | * | * | |||
| 40 | P. brevipes Ramme, 1951 | * | * | * | ||
| 41 | P. beieri (Ramme, 1951) | * | * | |||
| 42 | P. karabagi (Demirsoy, 1973) | * | * | |||
| 43 | P. lubricus Mistshenko, 1951 | * | * | |||
| 44 | P. straubei (Fieber, 1853) | * | * | |||
| 45 | P. tolunayi paphlagonicus Ramme, 1951 | * | * | * | ||
| 46 | P. tolunayi tolunayi Ramme, 1949 | * | * | * | ||
| 47 | P. turkmen Ünal, 2014 | * | * | |||
| 48 | P. fieberi (Brunner von Wattenwyl, 1882) | * | * | * | ||
| Ocnerosthenus Massa, 1995 | ||||||
| 49 | O. brunnerianus (Saussure, 1887) | * | ||||
| Eunothrotes Adelung, 1907 | ||||||
| 50 | E. derjugini Adelung, 1907 | * | ||||
| Paranothrotes Mistshenko, 1951 | ||||||
| 51 | P. sulcatus (Bolivar, 1912) | * | ||||
| 52 | P. asulcatus Demirsoy, 1973 | * | * | |||
| 53 | P. buzuldagi Ünal, 2016 | * | * | |||
| 54 | P. dentatus Ünal, 2016 | * | * | |||
| 55 | P. eximius bitlis Ünal, 2016 | * | * | |||
| 56 | P. eximius eximius Mistshenko, 1951 | * | ||||
| 57 | P. nigrolobus (Demirsoy, 1973) | * | * | |||
| 58 | P. kosswigi Demirsoy, 1973 | * | ||||
| 59 | P. gotvendicus Bolivar, 1912 | * | ||||
| 60 | P. opacus opacus (Brunner von Wattenwyl, 1882) | * | ||||
| 61 | P. opacus rectus (Mistshenko, 1951) | * | ||||
| 62 | P. siirt Ünal, 2016 | * | ||||
| Pseudonothrotes Mistshenko, 1951 | ||||||
| 63 | P. levis Mistshenko, 1951 | * | * | |||
| Nocarodes Fischer von Waldheim, 1846 | ||||||
| 64 | N. aserbus Mistshenko, 1951 | * | * | |||
| 65 | N. nodosus Mistshenko, 1951 | * | * | |||
| 66 | N. serricollis Fischer von Waldheim, 1846 | * | * | |||
| Nocaracris Uvarov, 1928 | ||||||
| 67 | N. acinosus (Mistshenko, 1951) | * | * | * | ||
| 68 | N. bicoloripes (Uvarov, 1949) | * | * | * | ||
| 69 | N. bodenheimeri (Uvarov, 1940) | * | * | |||
| 70 | N. burri (Uvarov, 1949) | * | * | * | ||
| 71 | N. cejchani Ünal, 2016 | * | * | |||
| 72 | N. cinerascens Ramme, 1951 | * | * | |||
| 72 | N. cinerascens Ramme, 1951 | * | * | |||
| 73 | N. citripes (Uvarov, 1949) | * | * | * | ||
| 74 | N. crassipes Ünal, 2016 | * | * | |||
| 75 | N. cyanipes (Fischer von Waldheim, 1846) | * | * | * | ||
| 76 | N. demirsoyi (Ünal, 2002) | * | * | |||
| 77 | N. dilekensis Ünal, 2016 | * | * | |||
| 78 | N. elegans (Mistshenko, 1951) | * | * | |||
| 79 | N. emirdagi Ünal, 2016 | * | * | |||
| 80 | N. furvus furvus (Mistshenko, 1951) | * | * | |||
| 81 | N. furvus kazdagi Ünal, 2016 | * | * | |||
| 82 | N. goektepe Ünal, 2016 | * | * | |||
| 83 | N. idrisi (Karabağ, 1953) | * | * | |||
| 84 | N. istanbul Ünal, 2016 | * | * | |||
| 85 | N. karadagi Ünal, 2016 | * | * | |||
| 86 | N. karshitoros Ünal, 2016 | * | * | |||
| 87 | N. kosswigi (Karabağ, 1953) | * | * | |||
| 88 | N. minutus Ünal, 2016 | * | * | |||
| 89 | N. monticolus Ünal, 2016 | * | * | |||
| 90 | N. niethammeri (Ramme, 1951) | * | * | * | ||
| 91 | N. palandoken Ünal, 2016 | * | * | |||
| 92 | N. pontica Ramme, 1951 | * | * | |||
| 93 | N. rubripes (Motschulsky, 1846) | * | ||||
| 94 | N. subrubrata (Ramme, 1951) | * | ||||
| 95 | N. sureyana Ramme, 1951 | * | * | * | ||
| 96 | N. tardus Ünal, Bugrov et Jetybayev, 2016 | * | * | |||
| 97 | N. tauricola Ramme, 1951 | * | * | |||
| 98 | N. tecticollis Ramme, 1951 | * | * | |||
| 99 | N. tridentatus (Stshelkanovtzev, 1916) | * | * | * | ||
| 100 | N. tunceli Ünal, 2016 | * | * | |||
| 101 | N. van Ünal, 2016 | * | * | |||
| 102 | N. sabulosa Ramme, 1951 | * | * | |||
| Prionosthenus Bolívar, 1878 | ||||||
| 103 | P. gueleni Karabağ, 1956 | * | * | |||
| Orchamus Stål, 1876 | ||||||
| 104 | O. yersini yersini (Brunner von Wattenwyl, 1882) | * | ||||
| 105 | O. yersini davisi Uvarov, 1949 | * | ||||
| 106 | O. massai Ünal, 2016 | * | * | |||
| Anacridium Uvarov, 1923 | ||||||
| 108 | A. aegyptium (Linnaeus, 1764) | * | * | * | * | |
| Subfamily Eyprepocnemidinae Brunner von Wattenwyl, 1893 | ||||||
| Eyprepocnemis Fieber, 1853 | ||||||
| 109 | E. plorans plorans (Charpentier, 1825) | * | * | * | * | |
| Heteracris Walker, 1870 | ||||||
| 110 | H. adspersa (Redtenbacher, 1889) | * | ||||
| 111 | H. littoralis littoralis (Rambur, 1838) | * | * | * | ||
| 112 | H. pterosticha (Fischer de Waldheim, 1833) | * | * | * | ||
| Subfamily Calliptaminae Jacobson, 1905 | ||||||
| Paracaloptenus Bolívar, 1878 | ||||||
| 113 | P. caloptenoides brunneri (Stal, 1876) | * | ||||
| 114 | P. caloptenoides caloptenoides (Brunner von Wattenwyl, 1861) | * | * | |||
| Calliptamus Serville, 1831 | ||||||
| 115 | C. coelesyriensis (Giglio-Tos, 1893) | * | * | |||
| 116 | C. italicus italicus (Linnaeus, 1758) | * | * | * | * | |
| 117 | C. barbarus barbarus (Costa, 1836) | * | * | * | * | |
| 118 | C. barbarus cephalotes (Fischer de Waldheim, 1846) | * | * | |||
| 119 | C. tenuicercis Tarbinsky, 1930 | * | * | * | * | |
| Subfamily Melanoplinae Scudder, 1897 | ||||||
| Podisma Berthold, 1827 | ||||||
| 120 | P. pedestris pedestris (Linnaeus, 1758) | * | ||||
| Rammepodisma Weidner, 1969 | ||||||
| 121 | R. natoliae (Ramme, 1939) | * | * | |||
| Micropodisma Dovnar-Zapolskyi, 1933 | ||||||
| 122 | M. koenigi (Burr, 1913) | * | ||||
| Subfamily Pezotettiginae Brunner von Wattenwyl, 1893 | ||||||
| Pezotettix Burmeister, 1840 | ||||||
| 123 | P. giornae (Rossi, 1794) | * | * | |||
| 124 | P. anatolica Uvarov, 1934 | * | ||||
| 125 | P. platycerca (Stal, 1876) | * | ||||
| Sphenophyma Uvarov, 1934 | ||||||
| 126 | S. rugulosa (Stal, 1876) | * | * | * | ||
| Subfamily Egnatiinae Bey-Bienko, 1951 | ||||||
| Charora Saussure, 1888 | ||||||
| 127 | C. pentagrammica Bolívar, 1899 | * | * | * | * | |
| Subfamily Acridinae MacLeay, 1821 | ||||||
| Acrida Linnaeus, 1758 | ||||||
| 128 | A. ungarica (Herbst, 1786) | * | * | |||
| 129 | A. anatolica Dirsh, 1949 | * | * | * | * | |
| 130 | A. bicolor (Thunberg, 1815) | * | * | * | * | |
| 131 | A. oxycephala (Pallas, 1771) | * | * | * | ||
| Truxalis Fabricius, 1775 | ||||||
| 132 | T. eximia eximia Eichwald, 1830 | * | * | * | ||
| 133 | T. robusta robusta (Uvarov, 1916) | * | * | * | ||
| Duroniella Bolívar, 1908 | ||||||
| 134 | D. fracta (Krauss, 1890) | * | * | * | * | |
| 135 | D. laticornis (Krauss, 1909) | * | * | |||
| Subfamily Oedipodinae Walker, 1871 | ||||||
| Demirsoyus Sirin & Çiplak, 2004 | ||||||
| 136 | D. salmani Şirin & Çıplak, 2004 | * | * | |||
| Paracinema Fischer, 1853 | ||||||
| 137 | P. tricolor bisignatum (Charpentier, 1825) | * | * | * | ||
| Mecostethus Fieber, 1852 | ||||||
| 138 | M. parapleurus parapleurus (Hagenbach, 1822) | * | * | |||
| Morphacris Walker, 1870 | ||||||
| 139 | M. fasciata (Thunberg, 1815) | * | ||||
| Stethophyma Fischer, 1853 | ||||||
| 140 | S. grossum (Linnaeus, 1758) | * | ||||
| Aiolopus Fieber, 1853 | ||||||
| 141 | A. simulatrix simulatrix (Walker, 1870) | * | * | |||
| 142 | A. strepens (Latreille, 1804) | * | * | * | * | |
| 143 | A. thalassinus thalassinus (Fabricius, 1781) | * | * | * | * | |
| Locusta Linnaeus, 1758 | ||||||
| 144 | L. migratoria migratoria (Linnaeus, 1758) | * | * | * | * | |
| Pyrgodera Fischer von Waldheim, 1846 | ||||||
| 145 | P. armata (Fischer von Waldheim, 1820) | * | * | * | ||
| Oedaleus Fieber, 1853 | ||||||
| 146 | O. decorus (Germar, 1825) | * | * | * | * | |
| Scintharista Sausseure, 1884 | ||||||
| 147 | S. notabilis miramae Uvarov, 1941 | * | * | |||
| Psophus Fieber, 1853 | ||||||
| 148 | P. stridulus (Linnaeus, 1758) | * | * | |||
| Brunnerella Saussure, 1888 | ||||||
| 149 | B. mirabilis mirabilis Saussure, 1888 | * | ||||
| Celes Saussure, 1884 | ||||||
| 150 | C. variabilis variabilis (Pallas, 1771) | * | ||||
| 151 | C. variabilis curtipennis Ramme, 1939 | * | * | |||
| 152 | C. variabilis carbonaria Uvarov, 1917 | * | * | * | ||
| Sphingonotus Fieber, 1852 | ||||||
| 153 | S. (S.) theodori theodori Uvarov, 1923 | * | * | * | ||
| 154 | S. (S.) pilosus Saussure, 1884 | * | * | * | * | |
| 155 | S. (S.) rubescens rubescens (Walker, 1870) | * | * | * | ||
| 156 | S. (S.) caerulans caerulans (Linnaeus, 1767) | * | * | |||
| 157 | S. (S.) coerulipes coerulipes Uvarov, 1922 | * | * | * | ||
| 158 | S. (S.) coerulipes djakanovi Mistshenko, 1937 | * | * | * | ||
| 159 | S. (S.) octofasciata (Serville, 1838) | * | ||||
| 160 | S. (S.) nebulosus persa Saussure, 1884 | * | ||||
| 161 | S. (S.) nebulosus discolor Uvarov, 1933 | * | ||||
| 162 | S. (S.) nebulosus anatolicus Uvarov, 1930 | * | * | |||
| 163 | S. (S.) turcicus turcicus Uvarov, 1930 | * | * | * | ||
| 164 | S. (S.) turcicus kocaki Demirsoy, 1977 | * | * | |||
| Sphingoderus Bei-Bienko, 1950 | ||||||
| 165 | S. carinatus (Saussure, 1888) | * | * | |||
| Asphingoderus Bei-Bienko, 1950 | ||||||
| 166 | A. uvarovites uvarovites (Mistshenko, 1937) | * | * | * | ||
| 167 | A. uvarovites similis Bey-Bienko, 1951 | * | * | |||
| 168 | A. elazigi Demirsoy, 1979 | * | * | |||
| Mioscirtus Saussure, 1888 | ||||||
| 169 | M. wagneri wagneri (Eversmann, 1859) | * | ||||
| 170 | M. wagneri rogenhoferi (Saussure, 1888) | * | ||||
| Oedipoda Latreille, 1829 | ||||||
| 171 | O. caerulescens caerulescens (Linnaeus, 1758) | * | * | * | * | |
| 172 | O. discessa Steinmann, 1965 | * | ||||
| 173 | O. schochii schochii Br.- Wattenwyl, 1884 | * | * | * | ||
| 174 | O. schochii caucasica Saussure, 1884 | * | * | |||
| 175 | O. schochi monotona Steinmann, 1965 | * | ||||
| 176 | O. aurea Uvarov, 1923 | * | * | * | * | |
| 177 | O. miniata miniata (Pallas, 1771) | * | * | * | * | |
| 178 | O. germanica germanica (Latreille, 1804) | * | * | |||
| 179 | O. meridionalis Ramme, 1913 | * | * | |||
| Acrotylus Fieber, 1853 | ||||||
| 180 | A. longipes longipes (Charpentier, 1845) | * | * | |||
| 181 | A. patruelis (Herrich-Schäffer, 1838) | * | * | |||
| 182 | A. insubricus insubricus (Scopoli, 1786) | * | * | * | ||
| Pseudoceles Bolivar, 1899 | ||||||
| 183 | P. ledereri ledereri (Brunner von Wattenwyl, 1884) | * | ||||
| 184 | P. oedipodioides Bolívar, 1899 | * | * | |||
| 185 | P. obscrus lateritius Karabağ 1957 | * | * | * | ||
| 186 | P. karadagi (Demirsoy, 1977) | * | * | |||
| Heliopteryx Uvarov, 1914 | ||||||
| 187 | H. humeralis (Kuthy, 1907) | * | * | * | * | |
| Leptopternis Saussure, 1884 | ||||||
| 188 | L. gracilis (Eversmann, 1848) | * | ||||
| Subfamily Gomphocerinae Fieber, 1853 | ||||||
| Ptygippus Mistshenko, 1951 | ||||||
| 189 | P. brachiopterus Mistshenko, 1951 | * | * | |||
| Xerohippus Uvarov, 1942 | ||||||
| 190 | X. alkani Karabag, 1953 | * | * | |||
| 191 | X. anatolicus Ramme, 1951 | * | * | |||
| Ochrilidia Stal, 1873 | ||||||
| 192 | O. pruinosa Brunner von Wattenwyl, 1882 | * | ||||
| 193 | O. gracilis gracilis (Krauss, 1902) | * | ||||
| 194 | O. tibialis (Fieber, 1853) | * | ||||
| Ramburiella Bolivar, 1906 | ||||||
| 195 | R. turcomana (Fischer de Waldheim, 1833) | * | * | |||
| 196 | R. bolivari (Kuthy, 1907) | * | * | |||
| Arcyptera Serville, 1839 | ||||||
| 197 | A. (A.) fusca (Pallas, 1773) | * | * | |||
| 198 | A. (Paracyptera) labiata (Brulle, 1832) | * | * | * | ||
| 199 | A. (P.) microptera microptera (Fischer de Waldheim, 1833) | * | ||||
| 200 | A. (P.) microptera karadagi Karabağ, 1956 | * | * | |||
| 201 | A. (P.) microptera transcaucasica Uvarov, 1917 | * | ||||
| Eremippus Uvarov, 1926 | ||||||
| 202 | E. angulatus Uvarov, 1934 | * | * | |||
| 203 | E. gracilis Uvarov, 1934 | * | * | |||
| 204 | E. turcicus Ramme, 1951 | * | * | |||
| 205 | E. simplex simplex (Eversmann, 1859) | * | ||||
| 206 | E. zeybekoglui Mol, 2012 | * | * | |||
| Stenobothrus Fischer, 1853 | ||||||
| 207 | S. stigmaticus stigmaticus (Rambur, 1839) | * | ||||
| 208 | S. zubowskyi Bolivar, 1899 | * | * | * | ||
| 209 | S. burri Karabağ, 1953 | * | * | * | ||
| 210 | S. lineatus lineatus (Panzer, 1796) | * | * | |||
| 211 | S. fischeri fischeri (Eversman, 1848) | * | * | * | ||
| 212 | S. nigromaculatus nigromaculatus (Herrich-Schäffer, 1840) | * | * | |||
| 213 | S. nigromaculatus transcaucasicus Ramme, 1933 | * | * | |||
| 214 | S. werneri werneri Adelung, 1907 | * | * | |||
| 215 | S. sviridenkoi Ramme, 1930 | * | ||||
| 216 | S. bozcuki Çıplak, 1994 | * | * | |||
| 217 | S. graecus malatyensis Çıplak, 1994 | * | * | |||
| 218 | S. derrai Harz, 1988 | * | * | |||
| 219 | S. eurasius eurasius Zubovski, 1898 | * | ||||
| 220 | S. miramae Dirsh, 1931 | * | * | |||
| 221 | S. selmae Ünal,1999 | * | * | |||
| 222 | S. weidneri Demirsoy, 1977 | * | * | |||
| Stauroderus Bolívar, 1897 | ||||||
| 223 | S. scalaris scalaris (Fischer-Waldheim, 1846) | * | * | |||
| 224 | S. scalaris znojkoi (Miram, 1938) | * | ||||
| Omocestus Bolivar, 1878 | ||||||
| 225 | O. nanus Uvarov, 1934 | * | * | |||
| 226 | O. viridulus (Linnaeus, 1758) | * | * | |||
| 227 | O. rufipes (Zetterstedt, 1821) | * | ||||
| 228 | O. haemorrhoidalis haemorrhoidalis (Charpentier, 1825) | * | * | |||
| 229 | O. haemorrhoidalis ciscaucasicus Mistshenko, 1951 | * | * | |||
| 230 | O. petraeus (Brisout de Barneville, 1856) | * | * | |||
| 231 | O. minutus (Brullé, 1832) | * | * | |||
| Euchorthippus Tarbinsky, 1926 | ||||||
| 232 | E. declivus (Brisout de Barneville, 1848) | * | * | |||
| 233 | E. pulvinatus (Fischer de Waldheim, 1849) | * | * | |||
| 234 | E. transcaucasicus Tarbinski, 1930 | * | ||||
| Chorthippus Fieber, 1852 | ||||||
| 235 | C. (C.) dichrous (Eversmann, 1859) | * | * | * | * | |
| 236 | C. (C.) labaumei Ramme, 1926 | * | * | * | ||
| 237 | C. (C.) karelini karelini (Uvarov, 1910) | * | ||||
| 238 | C. (C.) albomarginatus hakkaricus Demirsoy, 1977 | * | * | |||
| 239 | C. (C.) loratus (Fischer de Waldheim, 1846) | * | * | * | * | |
| 240 | C. (Glyptobothrus) bozdaghi Uvarov, 1934 | * | * | |||
| 241 | C. (G.) demokidovi (Ramme, 1930) | * | * | |||
| 242 | C. (G.) helverseni Mol, Çiplak & Sirin, 2003 | * | * | |||
| 243 | C. (G.) ilkazi Uvarov, 1934 | * | * | * | ||
| 244 | C. (G.) kazdaghensis Mol & Çiplak, 2005 | * | * | |||
| 245 | C. (G.) macrocerus macrocerus (Fischer de Waldheim, 1846) | * | * | * | ||
| 246 | C. (G.) apricarius apricarius (Linnaeus, 1758) | * | * | |||
| 247 | C. (G.) apricarius major (Pylnov, 1914) | * | * | |||
| 248 | C. (G.) vagans vagans (Eversman, 1848) | * | * | * | ||
| 249 | C. (G.) vagans dissimilis Willemse, Helversen et Odé, 2009 | * | ||||
| 250 | C. (G.) brunneus brunneus (Thunberg, 1815) | * | * | * | * | |
| 251 | C. (G.) biguttulus biguttulus (Linnaeaus, 1758) | * | * | * | * | |
| 252 | C. (G.) biguttulus euhedickei (Helversen, 1989) | * | ||||
| 253 | C. (G.) mollis mollis (Charpentier, 1825) | * | * | * | * | |
| 254 | C. (G.) relicticus Sirin, Helversen & Çiplak, 2010 | * | ||||
| 255 | C. (G.) taurensis Şirin & Çıplak, 2005 | * | * | |||
| 256 | C. (G.) aktaci Ünal, 2010 | * | * | |||
| 257 | C. (G.) antecessor Şirin & Çıplak, 2010 | * | * | |||
| Pseudochorthippus Defaut, 2012 | ||||||
| 258 | P. parallelus parallelus (Zetterstedt, 1821) | * | * | * | ||
| Euthystira Fieber, 1852 | ||||||
| 259 | E. brachyptera brachyptera (Ocskay, 1826) | * | ||||
| Rammeihippus Woznessenskij, 1996 | ||||||
| 260 | R. turcicus (Ramme, 1939) | * | * | * | ||
| Myrmeleotettix Bolivar, 1914 | ||||||
| 261 | M. maculatus maculatus (Thunberg, 1815) | * | * | * | ||
| 262 | M. ethicus Şirin & Çıplak, 2011 | * | * | |||
| Dasyhippus Uvarov, 1930 | ||||||
| 263 | D. escalerai (Bolivar, 1899) | * | * | |||
| 264 | D. uvarovi Karabağ, 1953 | * | * | |||
| Gomphocerus Thunberg, 1815 | ||||||
| 265 | G. armeniacus dimorphus Karabağ, 1953 | * | * | |||
| 266 | G. transcaucasicus Mistshenko, 1951 | * | * | |||
| 267 | G. sibiricus sibiricus (Linnaeus, 1767) | * | * | * | ||
| 268 | G. sibiricus acutus Karabağ, 1957 | * | * | |||
| 269 | G. sibiricus hemipterus Karabağ, 1953 | * | * | |||
| Aeropedellus Hebard, 1935 | ||||||
| 270 | A. turcicus Karabağ, 1959 | * | * | |||
| Dociostaurus Fieber, 1853 | ||||||
| 271 | D. (D.) maroccanus (Thunberg, 1815) | * | * | * | ||
| 272 | D. (D.) salmani Demirsoy, 1979 | * | * | |||
| 273 | D. (Kazakia) brevicollis (Eversman, 1848) | * | * | * | ||
| 274 | D. (K.) icconium Sirin & Mol, 2013 | * | * | |||
| 275 | D. (K.) tartarus Stshelkanovtzev, 1921 | * | ||||
| 276 | D. (K.) jagoi jagoi Soltani, 1978 | * | * | * | ||
| 277 | D. (Stauronotulus) hauensteini hauensteini (Bolívar, 1893) | * | * | * | ||
| 278 | D. (S.) hauensteini cappadocicus (Azam, 1913) | * | * | * | ||
| Notostaurus Bey-Bienko, 1933 | ||||||
| 279 | N. anatolicus (Krauss, 1896) | * | * | * | * | |
| Subfamily Tropidopolinae Jacobson, 1905 | ||||||
| Tropidopola Stal, 1873 | ||||||
| 280 | T. longicornis longicornis (Fieber, 1853) | * | ||||
| 281 | T. graeca graeca Uvarov, 1926 | * | * | * | ||
| Family Dericorythidae Jacobson &Bianchi, 1905 | ||||||
| Subfamily Dericorythinae Jacobson & Bianchi, 1905 | ||||||
| Dericorys Serville, 1838 | ||||||
| 282 | D. tibialis (Pallas, 1773) | * | * | |||
| 283 | D. albidula Serville, 1838 | * | * | |||
| 284 | D. uvarovi uvarovi Ramme, 1930 | * | ||||
References
- Bolivar, I. Orthopteres du voyage de M. escalera dans l’asie Mineure. Ann. Soc. Ent. Belg. 1899, 43, 583–606. [Google Scholar]
- Adelung, N. Beitrage zur Kenntnis der Orthopterafauna Transcaucasiens. Hor. Ent. Ross. 1907, 38, 32–82. [Google Scholar]
- Werner, F. Die Dermapteren- und Orthopterenfauna Kleinasiens. Sitz. Ber Akad. Wiss. Wien. 1901, 110, 259–305. [Google Scholar]
- Werner, F. Neue Locustiden aust Westasien. Zool. Anz. 1903, 26, 528–531. [Google Scholar]
- Werner, F. Ergebnisse einer naturwissenschaftlichen Reise zum Erdschias-Dag Kleinasiens, Dermapteren- und Orthopterenfauna. Ann. K. K. Nat. Hist. Hofmus 1905, 20, 168–170. [Google Scholar]
- Werner, F. Ergebnisse einer zoologischen Studien- und Sammelreise nach den Inseln des aegaeischen Meeres. II. Orthopteren. Sitz. Ber Akad. Wiss. Wien 1933, 142, 185–204. [Google Scholar]
- Uvarov, B.P. The geographical distribution of orthopterous insects in the Caucasus and in Western Asia. Proc. Zool. Soc. London 1921, 31, 447–472. [Google Scholar]
- Uvarov, B.P. Orthoptera Caucasia nova a cl. dom. K. A. Satunin lecta. Bul. Mus. Caucase 1916, 10, 1–9, (In Russian, descriptions in Latinish). [Google Scholar]
- Uvarov, B.P. Materialia ad cognitionem Orthopterorum Caucasi et cornium, II. Bul. Mus. Caucase 1917, 11, 281–298. [Google Scholar]
- Uvarov, B.P. Studies in the Orthoptera of Turkey, Iraq and Syria. EOS Rev. Esp. Entomol. 1934, 10, 21–119. [Google Scholar]
- Ebner, R. Wissenschaftliche Ergebnisse der Expedition nach Mesopotarnien Orthoptera I. Mantoidea und Tettigonioidea (=Locustodea). Ann. K. K. Natür. Hofmus. 1912, 26, 441–449. [Google Scholar]
- Ebner, R. VI. Orthopteren aus Kleinasien. Arch. Naturgesch. 1919, 8, 148–176. [Google Scholar]
- Ramme, W. Zur systematik, faunistik und biologie der Orthopteren von Sudost Europa und Vorderasien. Mitt. Zool. Mus. Berl. 1951, 27, 1–421. [Google Scholar]
- Ramme, W. Neue Orthopteren aus Westasien (Acrid. et Tett.). Mitt. Zool. Mus. Berl. 1930, 16, 394–396. [Google Scholar]
- Ramme, W. Beitrage zur Kenntnis der palaearktischen Orthopteren fauna (Tettig. et. Acrid). Mitt. Zool. Mus. Berl. 1931, 17, 165–200. [Google Scholar]
- Ramme, W. Beitrage zur palaearktischen Orthopteren fauna (Tettigon. et. Acrid). Mitt. Zool. Mus. Berl. 1933, 18, 416–434. [Google Scholar] [CrossRef]
- Ramme, W. Beitrage zur Kenntnis der palaearktischen Orthopterenfauna III (Tett. et. Acrid). Mitt. Zool. Mus. Berl. 1939, 22, 41–150. [Google Scholar]
- Bei-Bienko, G.J.; Mistshenko, L.L. The Grasshopper of the Fauna of the USSR and Adjacent Countries; Akademii Nauk: Moskow, Russia, 1951; Volume 2, pp. 1–400.
- Karabağ, T. Orthoptera Fauna of Turkey: A Synonymic and Distributional Catalogue of Turkish Orthoptera; Ankara Üniversitesi Fen Fakültesi Yayınları: Ankara, Turkey, 1958; p. 198.
- Weidner, H. Beitrage zur kenntnis der feldheuschenrecken (Caelifera) Anatoliens. Mitt. Zool. Mus. Berl. 1969, 66, 145–226. [Google Scholar]
- Demirsoy, A. Türkiye Caelifera (Insecta, Orthoptera) Faunasının Tespiti ve Taksonomik Incelenmesi; Atatürk Üniversitesi Yayınları: Erzurum, Turkey, 1977; p. 122.
- Çıplak, B.; Yalım, B.; Demirsoy, A. Türkiye Orthoptera (=Düzkanatlılar=Çekirge) Faunası. In Genel Zoocoğrafya ve Türkiye Zoocoğrafyası, 2nd ed.; Demirsoy, A., Ed.; Meteksan Press: Ankara, Turkey, 1999; pp. 748–772.
- Ünal, M. Checklist of Turkish Orthoptera. Available online: http://www.orthoptera-tr.org/ (accessed on 17 January 2023).
- Çıplak, B. Biogeography of Anatolia: The marker group Orthoptera. Mem. Soc. Entomol. Ital. 2004, 82, 357–372. [Google Scholar]
- Çıplak, B. Distribution of Tettigoniinae (Orthoptera, Tettigoniidae) bush-crickets in Turkey: The importance of the Anatolian Taurus Mountains in biodiversity and implications for conservation. Biodivers. Conserv. 2003, 12, 47–64. [Google Scholar] [CrossRef]
- Çıplak, B. The analogy between interglacial and global warming for the glacial relicts in a refugium: A biogeographic perspective for conservation of Anatolian Orthoptera. In Insect Ecology and Conservation; Fattorini, S., Ed.; Research Sign Post: Kerela, India, 2008; pp. 135–163.
- Çıplak, B.; Yahyaoğlu, Ö.; Uluar, O. Revisiting Pholidopterini (Orthoptera, Tettigoniidae): Rapid radiation causes homoplasy and phylogenetic instability. Zool. Scr. 2021, 50, 225–240. [Google Scholar] [CrossRef]
- Çıplak, B.; Demirsoy, A. Türkiye’de Ensifera (Orthoptera, Insecta) alttakımının endemizm açısından değerlendirilmesi. Turk. J. Zool. 1995, 19, 213–220. [Google Scholar]
- Çıplak, B.; Demirsoy, A. Caelifera (Orthoptera, Insecta) alttakımının Türkiye’de endemizm durumu. Turk. J. Zool. 1996, 20, 241–246. [Google Scholar] [CrossRef]
- Zohary, M. Geobotanical Foundations of the Middle East. Volume 2; Gustav Fischer Verlag: Stuttgart, Germany, 1973; Volume 2, p. 738.
- Demirsoy, A. Genel ve Turkiye Zoocografyası: Hayvan Cografyası, 5th ed.; Meteksan, A.Ş. Press: Ankara, Turkey, 2002; p. 1007.
- Ünal, M. Pamphagidae (Orthoptera: Acridoidea) from the Palaearctic Region: Taxonomy, classification, keys to genera and a review of the tribe Nocarodeini I. Bolívar. Zootaxa 2016, 4206, 1–223. [Google Scholar] [CrossRef]
- Şirin, D.; von Helversen, O.; Çıplak, B. Chorthippus brunneus subgroup (Orthoptera, Gomphocerinae) in Anatolia with description of two new species: Data suggest an Anatolian origin for the lineage. Zootaxa 2010, 2410, 1–28. [Google Scholar] [CrossRef]
- Kosswig, C. Zoogeography of the Near East. Syst. Zool. 1955, 4, 49–73. [Google Scholar] [CrossRef]
- Korkmaz, E.M.; Lunt, D.H.; Çıplak, B.; Değerli, N.; Babaşıbüyük, H.H. The contribution of Anatolia to European phylogeography: The centre of origin for the meadow grasshopper Chorthippus parallelus. J. Biogeogr. 2014, 41, 1793–1805. [Google Scholar] [CrossRef]
- Bücher, H. Die Heuschreckenplage und ihre Bekämpfung. Monogr. Angew. Entomol. 1918, 3, 1–274. [Google Scholar]
- Balamir, S. Yakın doğuda çöl çekirgesi durumu. Bitki Koruma Bülteni 1952, 3, 14–26. [Google Scholar]
- Balamir, S. Türkiye’de Çöl Çekirgesi (Schistocerca Gregaria Forsk.) Salgınları; Şark Matbaası: Ankara, Turkey, 1973; pp. 1–94.
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines: 1. Biology and Behaviour; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; p. 43.
- Çiplak, B. Locust and Grasshopper Outbreaks in the Near East: Review under Global Warming Context. Agronomy 2021, 11, 111. [Google Scholar] [CrossRef]
- Uvarov, B.P. Ecological studies on the Moroccan locust in Western Anatolia. Bull. Entomol. Res. 1932, 23, 273–287. [Google Scholar] [CrossRef]
- Balamir, S. Türkiye’de Fas çekirgesi (Dociostaurus maroccanus) durumu. Bitki Koruma Bülteni 1952, 1, 11–15. [Google Scholar]
- Erkılıç, S.S. Türkiye’nin Zararlı Çekirgelerini Birbirbinden ve Benzerlerinden Ayırma Anahtarı; Orhanbey Matbaası: Istanbul, Turkey, 1945; p. 40.
- Tutkun, E. Ankara İli Dahilinde Mevcut Caliptamus Türlerinin Morfolojisi ve Biyoekolojisi Üzerine Incelemeler; Yenigün Matbaası: Ankara, Turkey, 1974; p. 136.
- Balamir, S. Türkiye’de son yıllarda zararı görülen bir çekirge türü: Thisoicetrinus pterostichus. Bitki Koruma Bülteni 1962, 2, 3–10. [Google Scholar]
- Karabağ, T. Ankara Vilayeti Dahilinde Mevcut Çekirgelerin Ekolojik, Coğrafi ve Sistematik Durumları Üzerine Araştırmalar; Ankara Üniversitesi Ziraat Fakültesi Yayınları: Ankara, Turkey, 1949; p. 121.
- Türkeş, M. Küresel iklim değişikliği nedir? Temel kavramlar, nedenleri, gözlenen ve öngörülen değişiklikler. İklim Değişikliği ve Çevre 2008, 1, 26–37. [Google Scholar]
- Cigliano, M.M.; Braun, H.; Eades, D.C.; Otte, D. Orthoptera Species File. Version 5.0/5.0. Available online: http://orthoptera.speciesfile.org (accessed on 15 January 2023).
- Kaya, Z.; Raynal, D.J. Biodiversity and conservation of Turkish forests. Biol. Conserv. 2001, 97, 131–141. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modelling. R Package Version 2.5–8. 2017. Available online: https://cran.r-project.org/package=raster (accessed on 15 October 2023).
- Naimi, B.; Araújo, M.B. SDM: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, R Core Team: Vienna, Austria, 2013; Available online: http://www.r-project.org/ (accessed on 15 October 2023).
- Çıplak, B. Malatya ve Civarı Orthoptera (Insecta) Faunasının Incelenmesi. Ph.D. Thesis, İnönü Üniversitesi, Fen Bilimleri Enstitüsü, Malatya, Turkey; 1992. p. 134.
- Çıplak, B.; Mol, A.; Sirin, D.; Zeybekoglu, Ü.; Taylan, M.S. The demokidovi-like short winged Glyptobothrus (Orthoptera, Gomphocerinae, Chorthippus) of Anatolia with description of two new species: From Balkans to Caucasus through southern Anatolia. Trans. Am. Entomol. Soc. 2005, 131, 463–489. [Google Scholar]
- Demirsoy, A. Erzurum Bölgesi Orthoptera (Insecta) Faunasının Tesbiti ve Taxonomik İncelenmesi; Atatürk Üniversitesi Yayınları: Erzurum, Turkey, 1975; p. 122, No. 347.
- Erman, O.; Salman, S. Elazig ili Orthoptera Faunası. In Proceedings of the Erzurum Biyoloji Kongresi, Zooloji Bildirileri (Proceeding of Biological Congress, Zoology Section), Erzurum, Turkey; 18–20 July 1990. pp. 109–119.
- Karabag, T.; Balamir, S.; Gumusuyu, I.; Tutkun, E. Turkiye Orthoptera faunasının tesbiti üzerinde araştırmalar. Bitki Koruma Bulteni 1971, 11, 73–100. [Google Scholar]
- Karabag, T.; Balamir, S.; Gumusuyu, I.; Tutkun, E. Turkiye Orthoptera faunasının tesbiti üzerinde araştırmalar (II). Bitki Koruma Bulteni 1981, 20, 1–25. [Google Scholar]
- Mol, A.; Şirin, D.; Taylan, M.S. Türkiye’de dağılım gösteren bazı Caelifera (Insecta: Orthoptera) türlerinin yeni lokalite kayıtları, endemizm, yaygınlık ve tarımsal zarar oluşturma açısından değerlendirilmesi. Bitki Koruma Bülteni 2014, 54, 133–170. [Google Scholar]
- Salman, S. Ağrı, Kars ve Artvin Illerinin Orthoptera (Insecta) Faunasi Üzerine Taksonomik Araştırmalar; Atatürk Üniversitesi Yayınları: Erzurum, Turkey, 1978; p. 184, No. 184.
- Satar, A.; Özbay, C. On The Orthoptera (Insecta) fauna of the Karacadağ Mountains and the Tigris Basin (Diyarbakır, Turkey). Boletín Soc. Entomológica Aragonesa 2003, 32, 115–120. [Google Scholar]
- Sevgili, H. Şanlıurfa Yöresinde Orthoptera Faunasının Sistematik Açıdan Incelenmesi. Master’s Thesis, Harran Üniversitesi, Fen Bilimleri Enstitüsü, Şanlıurfa, Turkey. 1997.
- Sevgili, H.; Ciplak, B. The Orthoptera of Şanlıurfa province from the Mesopotamian part of Turkey. Ital. J. Zool. 2000, 67, 229–240. [Google Scholar] [CrossRef]
- Sevgili, H.; Demirsoy, A.; Durmuş, Y. Orthoptera fauna of Kemaliye, Erzincan. HJBC 2012, 40, 317–335. [Google Scholar]
- Ünal, M. Nevşehir ili Avanos çevresinin Orthopter’lerinin eko-faunası üzerine araştırmalar. Priamus 1997, 9, 1–54. [Google Scholar]
- Güneş, H.V. Doğu Akdeniz Bölgesi Orthoptera (Insecta) Faunası Üzerine Taksonomik Çalışmalar. Ph.D. Thesis, Ankara Üniversitesi, Fen Bilimleri Enstitüsü, Ankara, Turkey; 1984. p. 202.
- Naskrecki, P.; Ünal, M. The Orthoptera of Hatay Province, S. Turkey. Beiträge Zur Entomol. = Contrib. Entomol. 1995, 45, 393–419. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Zhang, D.-C.; Yin, H.; Yin, X.-C. On the taxonomic system of Eurasian Pamphagidae (Orthoptera: Caelifera). Acta Entomol. Sinica 2003, 46, 218–221. [Google Scholar]
- Harz, K. The Orthoptera of Europe; Dr. W. Junk, N. V.: The Hague, The Netherlands, 1975; Volume II, p. 749.
- Jago, N.D. A review of the Gomphocerinae of the world with a key to the genera (Orthoptera: Acrididae). Proc. Acad. Nat. Sci. Phladelphia 1971, 123, 205–343. Available online: http://books.google.com/books?id=ELz9FCjoVVkC (accessed on 15 October 2023).
- Karabag, T. Revision of Drymadusa Stein and related genera (Orthoptera: Tettigoniidae). Bull. Br. Mus. (Nat. Hist.) Ent. 1961, 11, 1–41. [Google Scholar]
- De Lattin, G. Grundriß der Zoogeographie; Fischer: Stuttgart, Germany, 1967; p. 602.
- Babaroğlu, N.; Directorate of Plant Protection Central Research Institute, Ankara, Türkiye. Personal communication. 2021.







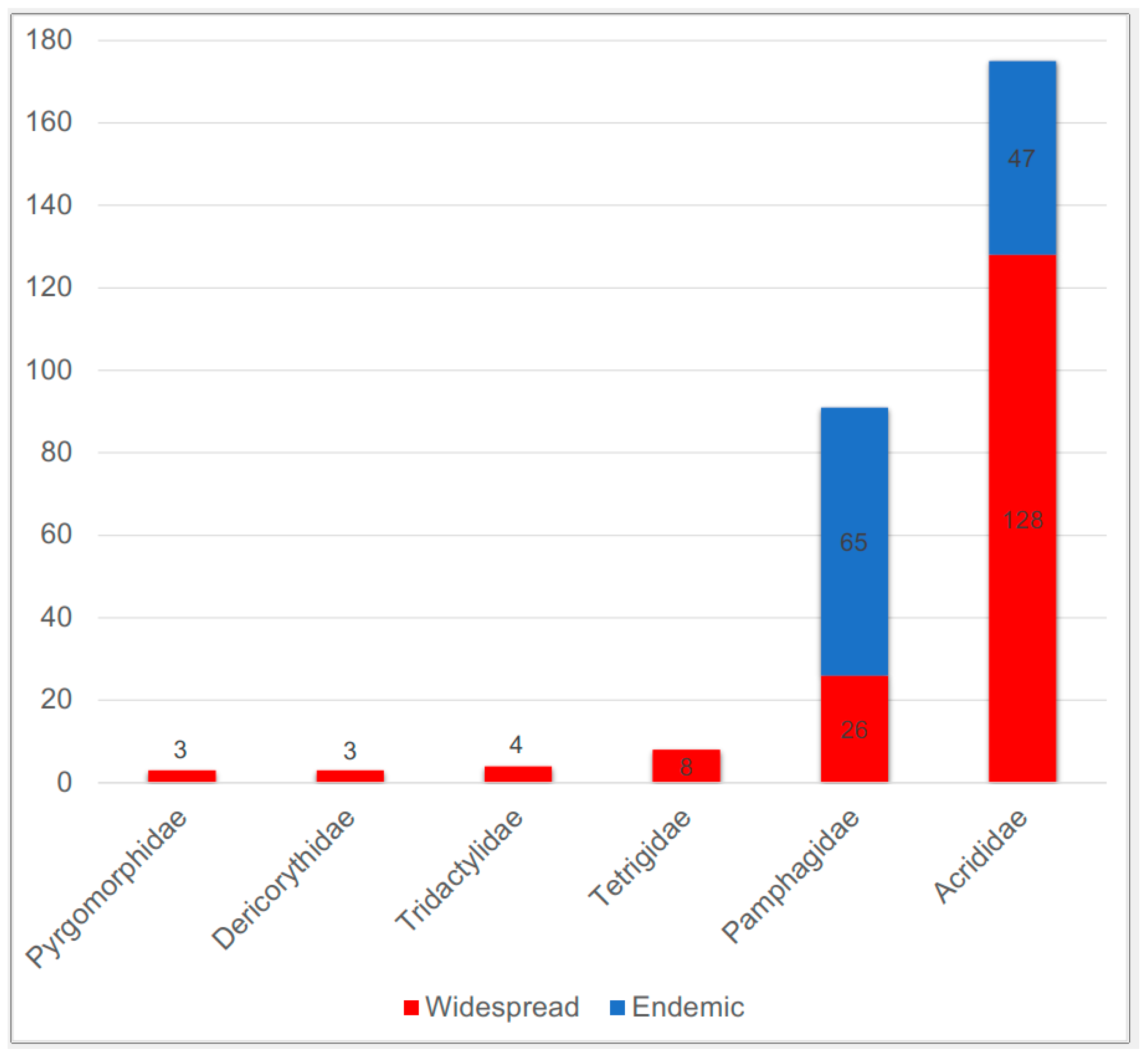
| Family | N, Widespread | N, Endemic | Total |
|---|---|---|---|
| Tridactylidae | 4 | 0 | 4 |
| Pyrgomorphidae | 3 | 0 | 3 |
| Dericorythidae | 3 | 0 | 3 |
| Tetrigidae | 8 | 0 | 8 |
| Pamphagidae | 26 | 65 | 91 |
| Acrididae | 128 | 47 | 175 |
| Total | 172 | 112 | 284 |
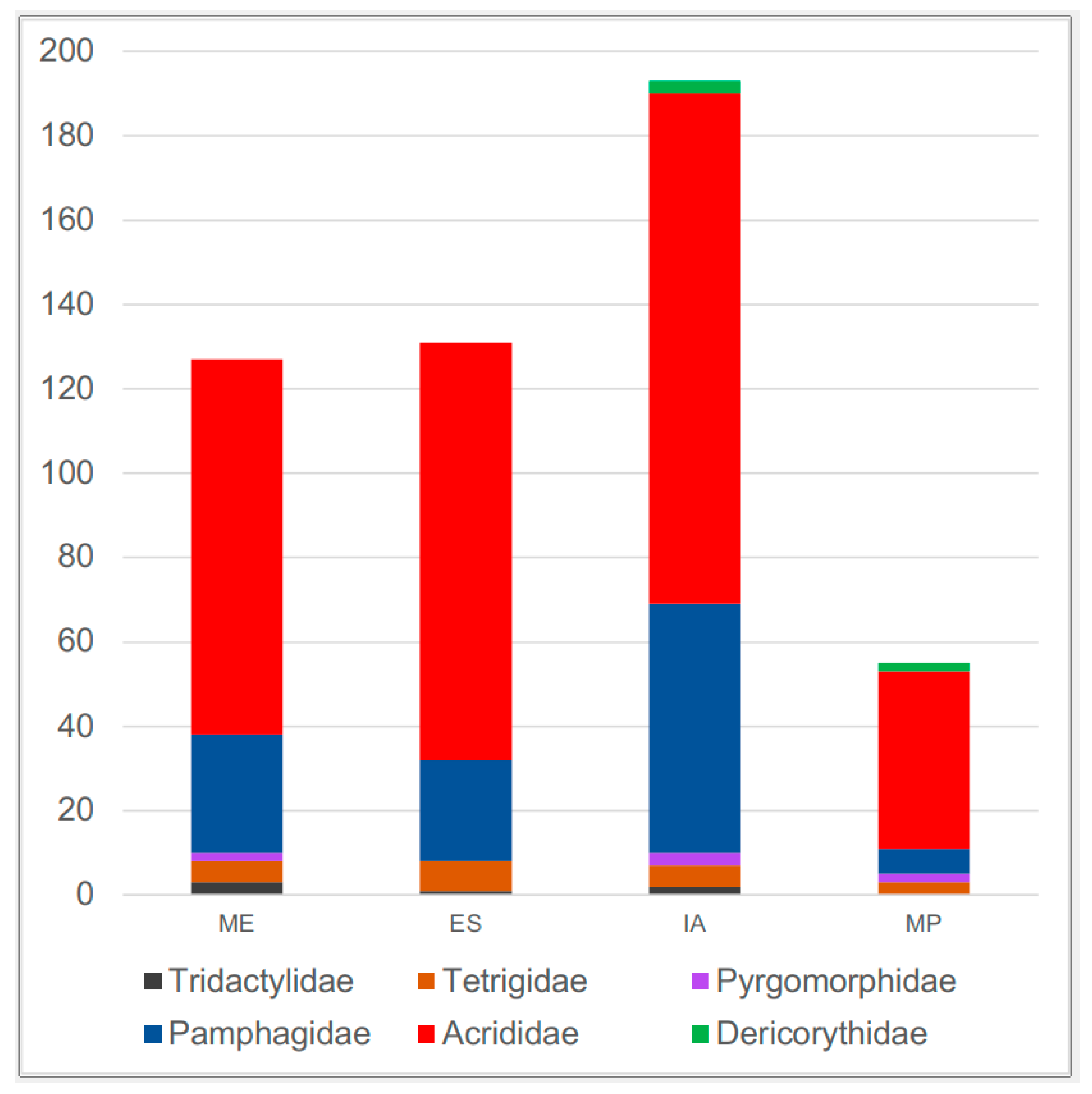
| Irano-Anatolia | Euro–Siberia | Mediterranean | Mesopotamia | |
|---|---|---|---|---|
| Dericorythidae | 3 | - | - | 2 |
| Pyrgomorphidae | 3 | - | 2 | 2 |
| Tridactylidae | 2 | 1 | 3 | - |
| Tetrigidae | 5 | 7 | 5 | 3 |
| Pamphagidae | 59 | 24 | 28 | 6 |
| Acrididae | 121 | 99 | 89 | 42 |
| Total | 193 | 131 | 127 | 55 |
| Bioclimatic Variables | Description | Bioclimatic Variables | Description |
|---|---|---|---|
| BIO1 | Annual mean temperature | BIO11 | Mean temperature of coldest quarter |
| BIO2 | Mean diurnal range (mean of monthly (max temp–min temp)) | BIO12 | Annual precipitation |
| BIO3 | Isothermality (BIO2/BIO7) (×100) | BIO13 | Precipitation in wettest month |
| BIO4 | Temperature seasonality (standard deviation ×100) | BIO14 | Precipitation in driest month |
| BIO5 | Max temperature of warmest month | BIO15 | Precipitation seasonality (coefficient of variation) |
| BIO6 | Min temperature of coldest month | BIO16 | Precipitation in wettest quarter |
| BIO7 | Temperature annual range (BIO5-BIO6) | BIO17 | Precipitation in driest quarter |
| BIO8 | Mean temperature of wettest quarter | BIO18 | Precipitation in warmest quarter |
| BIO9 | Mean temperature of driest quarter | BIO19 | Precipitation in coldest quarter |
| BIO10 | Mean temperature of warmest quarter |
| Bioclimatic Variables | LM | CI | CB | CT | DM | DB | HP | NA | AL | OM | OS | ChD | ChK | EP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BIO1 | 4.405 | 3.614 | 2.061 | 1.487 | * | 2.448 | * | 2.562 | 2.986 | * | 4.217 | 2.828 | * | * |
| BIO2 | 2.069 | 1.758 | 1.959 | 2.132 | 2.917 | 1.588 | 1.178 | 1.566 | 2.514 | * | * | 2.011 | 2.112 | 2.135 |
| BIO3 | 1.840 | 1.380 | * | * | * | * | * | * | * | 2.798 | 2.629 | * | 1.996 | 1.301 |
| BIO4 | * | * | 1.382 | 1.508 | 2.682 | 1.920 | 4.716 | 1.684 | 2.791 | 3.056 | * | 1.856 | * | * |
| BIO5 | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| BIO6 | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| BIO7 | * | * | * | * | * | * | * | * | * | * | 1.793 | * | * | * |
| BIO8 | 2.510 | 1.308 | 2.236 | 2.392 | 3.224 | 3.162 | 1.326 | 2.115 | 4.324 | 1.365 | 4.000 | 1.898 | 2.439 | 2.527 |
| BIO9 | * | 4.189 | * | * | 1.677 | 2.341 | 1.908 | 2.171 | * | 3.072 | 2.975 | 2.872 | 2.814 | 4.967 |
| BIO10 | * | * | * | * | * | * | * | * | * | * | * | * | 2.826 | 3.171 |
| BIO11 | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| BIO12 | 2.133 | * | 2.061 | 7.391 | * | * | * | 8.389 | 4.256 | 3.768 | 5.918 | 7.237 | 4.313 | * |
| BIO13 | * | 1.579 | 6.978 | 5.071 | * | 3.363 | 4.929 | 6.112 | 5.129 | * | 4.947 | 4.748 | * | 4.331 |
| BIO14 | 5.790 | 2.615 | 4.331 | 2.555 | * | 3.115 | 1.905 | 2.873 | 4.391 | 6.002 | 2.954 | 3.129 | 9.876 | |
| BIO15 | 6.361 | * | * | * | * | * | * | * | * | 9.466 | * | * | 5.524 | 5.768 |
| BIO16 | * | * | * | * | 2.981 | * | * | * | * | * | * | * | * | * |
| BIO17 | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| BIO18 | * | * | * | * | 4.062 | * | * | * | * | * | * | * | * | 7.324 |
| BIO19 | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| Species | Methods | AUC | TSS | Species | Methods | AUC | TSS |
|---|---|---|---|---|---|---|---|
| Arcyptera labiata | GLM | 0.79 | 0.61 | Dociostaurus maroccanus | GLM | 0.65 | 0.49 |
| GAM | 0.84 | 0.71 | GAM | 0.65 | 0.46 | ||
| MAXENT | 0.87 | 0.71 | MAXENT | 0.77 | 0.65 | ||
| Calliptamus barbarus | GLM | 0.69 | 0.41 | Euchortippus pulvinatus | GLM | 0.78 | 0.62 |
| GAM | 0.79 | 0.54 | GAM | 0.79 | 0.64 | ||
| MAXENT | 0.8 | 0.54 | MAXENT | 0.86 | 0.72 | ||
| Calliptamus italicus | GLM | 0.74 | 0.45 | Heteracris pterosticha | GLM | 0.75 | 0.61 |
| GAM | 0.84 | 0.61 | GAM | 0.75 | 0.58 | ||
| MAXENT | 0.82 | 0.56 | MAXENT | 0.82 | 0.71 | ||
| Calliptamus tenuicercis | GLM | 0.71 | 0.47 | Oedipoda schochii | GLM | 0.79 | 0.63 |
| GAM | 0.79 | 0.55 | GAM | 0.84 | 0.69 | ||
| MAXENT | 0.81 | 0.59 | MAXENT | 0.85 | 0.72 | ||
| Chorthippus dichrous | GLM | 0.74 | 0.45 | Oedipoda miniata | GLM | 0.69 | 0.41 |
| GAM | 0.79 | 0.52 | GAM | 0.76 | 0.52 | ||
| MAXENT | 0.8 | 0.54 | MAXENT | 0.75 | 1.2 | ||
| Chorthippus karelini | GLM | 0.83 | 0.65 | Locusta migratoria | GLM | 0.78 | 0.58 |
| GAM | 0.84 | 0.66 | GAM | 0.79 | 0.6 | ||
| MAXENT | 0.84 | 0.68 | MAXENT | 0.83 | 0.67 | ||
| Dociostaurus brevicollis | GLM | 0.78 | 0.56 | Notostaurus anatolicus | GLM | 0.71 | 0.47 |
| GAM | 0.86 | 0.69 | GAM | 0.78 | 0.56 | ||
| MAXENT | 0.85 | 0.66 | MAXENT | 0.8 | 0.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çıplak, B.; Uluar, O. Anatolian Short-Horned Grasshoppers Unveiled: Integrating Biogeography and Pest Potential. Insects 2024, 15, 55. https://doi.org/10.3390/insects15010055
Çıplak B, Uluar O. Anatolian Short-Horned Grasshoppers Unveiled: Integrating Biogeography and Pest Potential. Insects. 2024; 15(1):55. https://doi.org/10.3390/insects15010055
Chicago/Turabian StyleÇıplak, Battal, and Onur Uluar. 2024. "Anatolian Short-Horned Grasshoppers Unveiled: Integrating Biogeography and Pest Potential" Insects 15, no. 1: 55. https://doi.org/10.3390/insects15010055
APA StyleÇıplak, B., & Uluar, O. (2024). Anatolian Short-Horned Grasshoppers Unveiled: Integrating Biogeography and Pest Potential. Insects, 15(1), 55. https://doi.org/10.3390/insects15010055