
Barcoding Fails to Delimit Species in Mongolian Oedipodinae (Orthoptera, Acrididae)


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Material
2.2. Preparation of DNA for Sequencing
2.3. Sequence Analyses
3. Results
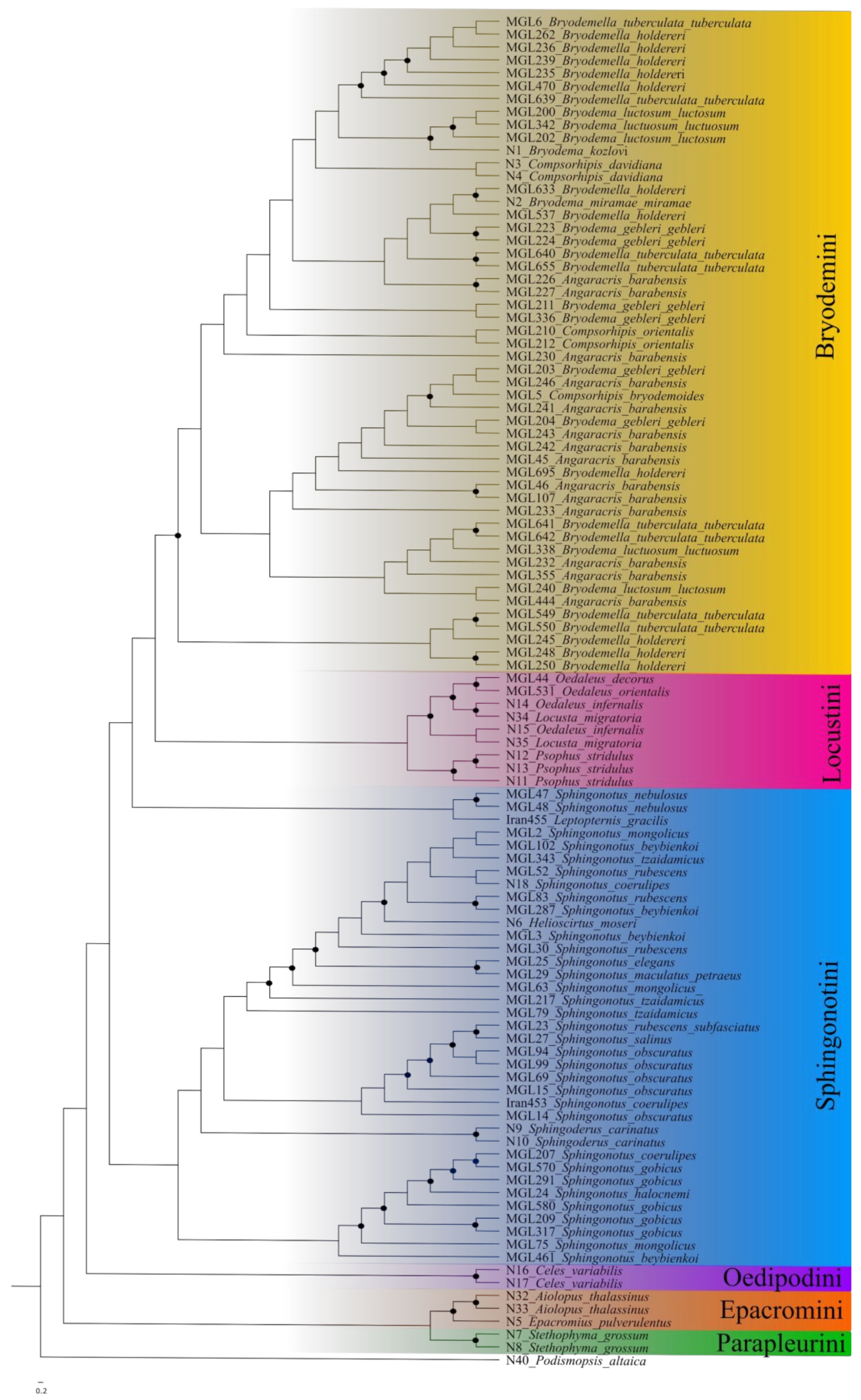
3.1. Maximum Likelihood
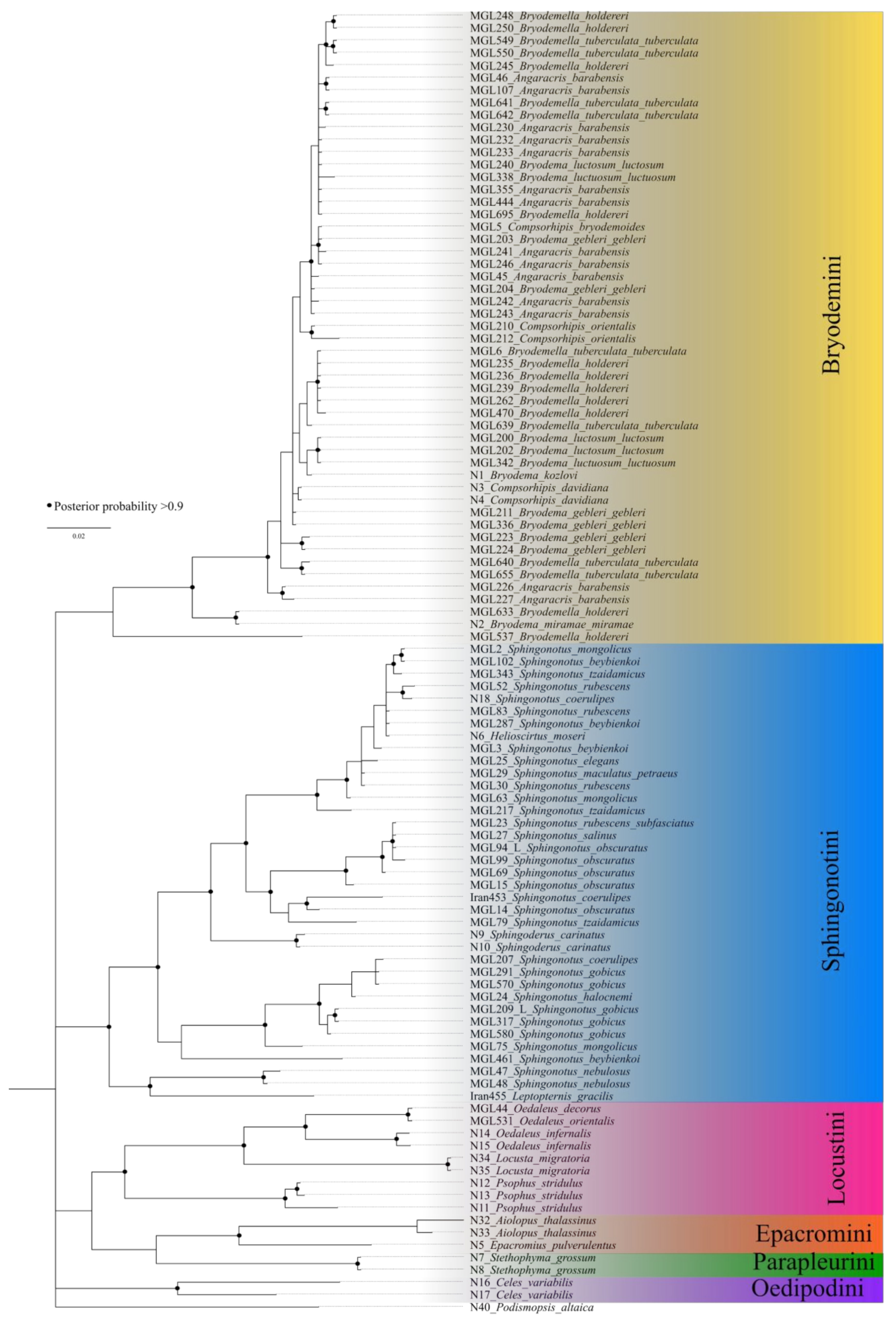
3.2. Bayesian Inference Tree
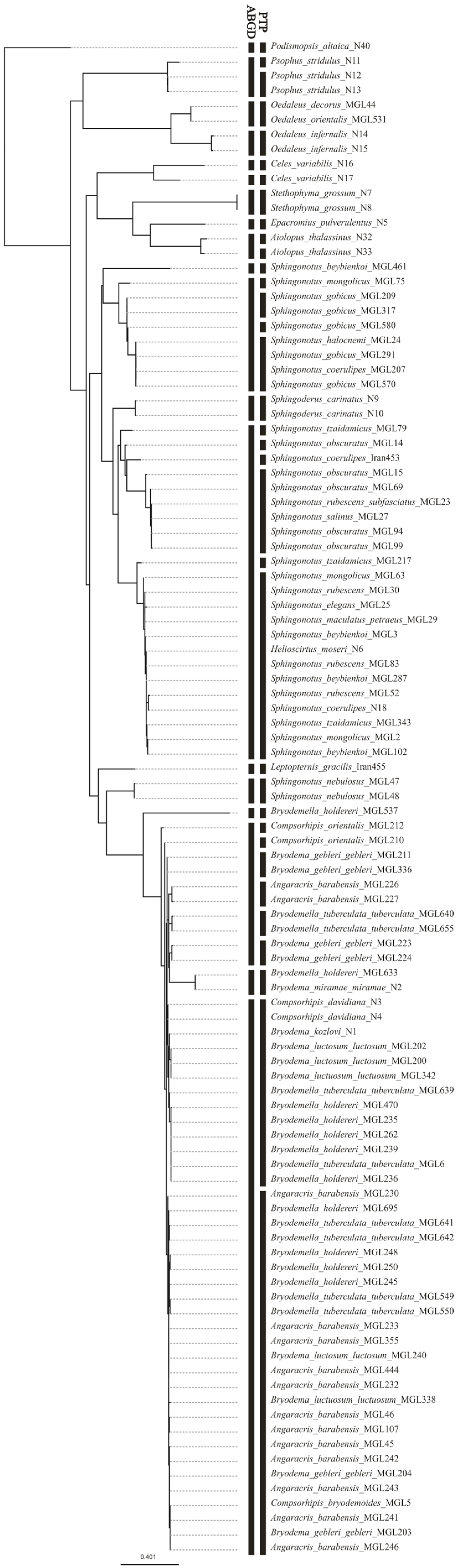
3.3. Species Delimitation
4. Discussion
4.1. Basal Genera with Little Diversity
4.2. Bryodemini and Sphingonotini
4.3. Barcoding as a Solution for Species Delimitation?
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| ID | Species | Location | BOLD ID |
|---|---|---|---|
| Iran453 | Sphingonotus coerulipes | Iran, 36°31.811′ N, 054°50.438′ E | BMGL001-24 |
| Iran455 | Leptopternis gracilis | Iran, 35°21.106′ N, 51°59.297′ E | BMGL002-24 |
| MGL2 | Sphingonotus mongolicus | Mongolia, 44°24′27.6″ N, 105°21′22.0″ E | BMGL003-24 |
| MGL3 | Sphingonotus beybienkoi | Mongolia, 44°24′27.6″ N, 105°21′22.0″ E | BMGL004-24 |
| MGL5 | Compsorhipis bryodemoides | Mongolia, 45°43′07.0″ N, 107°17′30.3″ E | BMGL005-24 |
| MGL6 | Bryodemella tuberculata tuberculata | Mongolia, 47°41′33.0″ N, 105°48′42.6″ E | BMGL006-24 |
| MGL14 | Sphingonotus obscuratus | Mongolia, Echingol | BMGL007-24 |
| MGL15 | Sphingonotus obscuratus | Mongolia, Echingol | BMGL008-24 |
| MGL23 | Sphingonotus rubescens subfasciatus | Mongolia, 44°19′133″ N, 105°22′40.3″ E | BMGL009-24 |
| MGL24 | Sphingonotus halocnemi | Mongolia, Shar Bulag | BMGL010-24 |
| MGL25 | Sphingonotus elegans | Mongolia, Bayan Bulag | BMGL011-24 |
| MGL27 | Sphingonotus salinus | Mongolia, Archangai Aimag | BMGL012-24 |
| MGL29 | Sphingonotus maculatus petraeus | Mongolia, 42°56′47″ N, 107°46′52.2″ E | BMGL013-24 |
| MGL30 | Sphingonotus rubescens | Mongolia, 43°5′103″ N, 107°29′65.9″ E | BMGL014-24 |
| MGL44 | Oedaleus decorus | Mongolia, 47°39′30.3″ N, 106°42′29.0″ E | BMGL015-24 |
| MGL45 | Angaracris barabensis | Mongolia, 47°39′30.3″ N, 106°42′29.0″ E | BMGL016-24 |
| MGL46 | Angaracris barabensis | Mongolia, 47°39′30.3″ N, 106°42′29.0″ E | BMGL017-24 |
| MGL47 | Sphingonotus nebulosus | Mongolia, 42°38′30.8″ N, 105°20′39.3″ E | BMGL018-24 |
| MGL48 | Sphingonotus nebulosus | Mongolia, 42°38′30.8″ N, 105°20′39.3″ E | BMGL019-24 |
| MGL52 | Sphingonotus rubescens | Mongolia, 42°38′30.8″ N, 105°20′39.3″ E | BMGL020-24 |
| MGL63 | Sphingonotus mongolicus | Mongolia, 44°24′27.6″ N, 105°21′22.0″ E | BMGL021-24 |
| MGL69 | Sphingonotus obscuratus | Mongolia, 42°36′23.6″ N, 106°55′53.2″ E | BMGL022-24 |
| MGL75 | Sphingonotus mongolicus | Mongolia, 43°31′11.6″ N, 104°07′06.3″ E | BMGL023-24 |
| MGL79 | Sphingonotus tzaidamicus | Mongolia, 42°45′31.04″ N, 106°59′40.04″ E | BMGL024-24 |
| MGL83 | Sphingonotus rubescens | Mongolia, 43°53′49.5″ N, 107°39′43.5″ E | BMGL025-24 |
| MGL94_L | Sphingonotus obscuratus | Mongolia, 42°36′23.6″ N, 106°55′53.2″ E | BMGL026-24 |
| MGL99 | Sphingonotus obscuratus | Mongolia, 42°35′40.5″ N, 106°46’ 47.4″ E | BMGL027-24 |
| MGL102 | Sphingonotus beybienkoi | Mongolia, 42°35′40.5″ N, 106°46′47.4″ E | BMGL028-24 |
| MGL107 | Angaracris barabensis | Mongolia, no detailed record given | BMGL029-24 |
| MGL200 | Bryodema luctosum luctosum | Mongolia, 46°24′55.4868″ N, 102°50′10.212″ E | BMGL030-24 |
| MGL202 | Bryodema luctosum luctosum | Mongolia, 46°24′55.4868″ N, 102°50′10.212″ E | BMGL031-24 |
| MGL203 | Bryodema gebleri gebleri | Mongolia, 46°8′11.1588″ N, 99°10′39.018″ E | BMGL032-24 |
| MGL204 | Bryodema gebleri gebleri | Mongolia, 46°8′11.1588″ N, 99°10′39.018″ E | BMGL033-24 |
| MGL207 | Sphingonotus coerulipes | Mongolia, 46°16′5.7936″ N, 97°26′48.534″ E | BMGL034-24 |
| MGL209_L | Sphingonotus gobicus | Mongolia, 46°16′5.7936″ N, 97°26′48.534″ E | BMGL035-24 |
| MGL210 | Compsorhipis orientalis | Mongolia, 46°16′5.7936″ N, 97°26′48.534″ E | BMGL036-24 |
| MGL211 | Bryodema gebleri gebleri | Mongolia, 46°23′59.0856″ N, 96°23′10.32″ E | BMGL037-24 |
| MGL212 | Compsorhipis orientalis | Mongolia, 46°22′54.5412″ N, 96°16′16.968″ E | BMGL038-24 |
| MGL217 | Sphingonotus tzaidamicus | Mongolia, 48°19′59.628″ N, 92°49′5.808″ E | BMGL039-24 |
| MGL223 | Bryodema gebleri gebleri | Mongolia, 47°41′41.5464″ N, 96°31′7.968″ E | BMGL040-24 |
| MGL224 | Bryodema gebleri gebleri | Mongolia, 47°41′41.5464″ N, 96°31′7.968″ E | BMGL041-24 |
| MGL226 | Angaracris barabensis | Mongolia, 47°41′41.5464″ N, 96°31′7.968″ E | BMGL042-24 |
| MGL227 | Angaracris barabensis | Mongolia, 47°43′37.02″ N, 96°14′9.24″ E | BMGL043-24 |
| MGL230 | Angaracris barabensis | Mongolia, 48°16′6.8664″ N, 99°45′52.434″ E | BMGL044-24 |
| MGL232 | Angaracris barabensis | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL045-24 |
| MGL233 | Angaracris barabensis | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL046-24 |
| MGL235 | Bryodemella holdereri | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL047-24 |
| MGL236 | Bryodemella holdereri | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL048-24 |
| MGL239 | Bryodemella holdereri | Mongolia, 47°56′20.562″ N, 100°36′49.896″ E | BMGL049-24 |
| MGL240 | Bryodema luctosum luctosum | Mongolia, 47°56′20.562″ N, 100°36′49.896″ E | BMGL050-24 |
| MGL241 | Angaracris barabensis | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL051-24 |
| MGL242 | Angaracris barabensis | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL052-24 |
| MGL243 | Angaracris barabensis | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL053-24 |
| MGL245 | Bryodemella holdereri | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL054-24 |
| MGL246 | Angaracris barabensis | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL055-24 |
| MGL248 | Bryodemella holdereri | Mongolia, 47°20′48.9264″ N, 104°17′6.1224″ E | BMGL056-24 |
| MGL250 | Bryodemella holdereri | Mongolia, 47°43′0.624″ N, 101°37′15.276″ E | BMGL057-24 |
| MGL262 | Bryodemella holdereri | Mongolia, 47°28′16.3992″ N, 101°37′15.276″ E | BMGL058-24 |
| MGL287 | Sphingonotus beybienkoi | Mongolia, 46°21′28.6812″ N, 95°24′34.236″ E | BMGL059-24 |
| MGL291 | Sphingonotus gobicus | Mongolia, 46°21′28.6812″ N, 95°24′34.236″ E | BMGL060-24 |
| MGL317 | Sphingonotus gobicus | Mongolia, 48°17′39.552″ N, 93°29′19.14″ E | BMGL061-24 |
| MGL336 | Bryodema gebleri gebleri | Mongolia, 46°16′5.7936″ N, 97°26′48.534″ E | BMGL062-24 |
| MGL338 | Bryodema luctuosum luctuosum | Mongolia, 46°23′59.0856″ N, 96°23′10.32″ E | BMGL063-24 |
| MGL342 | Bryodema luctuosum luctuosum | Mongolia, 46°24′55.4868″ N, 102°50′10.212″ E | BMGL064-24 |
| MGL343 | Sphingonotus tzaidamicus | Mongolia, 48°54′3.42″ N, 93°19′18.336″ E | BMGL065-24 |
| MGL355 | Angaracris barabensis | Mongolia, 48°46′41.016″ N, 97°14′21.354″ E | BMGL066-24 |
| MGL444 | Angaracris barabensis | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL067-24 |
| MGL461 | Sphingonotus beybienkoi | Mongolia, 42°53.249’ N, 98°54.708’ E | BMGL068-24 |
| MGL470 | Bryodemella holdereri | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL069-24 |
| MGL531 | Oedaleus orientalis | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL070-24 |
| MGL537 | Bryodemella holdereri | Mongolia, 48°8′42.702″ N, 100°12′18.144″ E | BMGL071-24 |
| MGL549 | Bryodemella tuberculata tuberculata | Mongolia, 42°52.930′ N, 98°51.818′ E | BMGL072-24 |
| MGL550 | Bryodemella tuberculata tuberculata | Mongolia, no detailed record given | BMGL073-24 |
| MGL570 | Sphingonotus gobicus | Mongolia, 42°52.930′ N, 98°51.818′ E | BMGL074-24 |
| MGL580 | Sphingonotus gobicus | Mongolia, 42°52.930′ N, 98°51.818′ E | BMGL075-24 |
| MGL633 | Bryodemella holdereri | Mongolia, 45°46′46.146″ N, 107°15′8.0928″ E | BMGL076-24 |
| MGL639 | Bryodemella tuberculata tuberculata | Mongolia, 47°46′51.96″ N, 108°49′26.9832″ E | BMGL077-24 |
| MGL640 | Bryodemella tuberculata tuberculata | Mongolia, 47°46′51.96″ N, 108°49′26.9832″ E | BMGL078-24 |
| MGL641 | Bryodemella tuberculata tuberculata | Mongolia, 47°46′51.96″ N, 108°49′26.9832″ E | BMGL079-24 |
| MGL642 | Bryodemella tuberculata tuberculata | Mongolia, 47°46′51.96″ N, 108°49′26.9832″ E | BMGL080-24 |
| MGL655 | Bryodemella tuberculata tuberculata | Mongolia, 49°23′4.614″ N, 113°30′30.042″ E | BMGL081-24 |
| MGL695 | Bryodemella holdereri | Mongolia, 47°37′41.0304″ N, 105°54′2.4696″ E | BMGL082-24 |
References
- Gillon, Y. The Invertebrates of the Grass Layer. Ecosyst. World 1983, 13, 289–311. [Google Scholar]
- Gankhuyag, E.; Dorjsuren, A.; Choi, E.H.; Hwang, U.W. An Annotated Checklist of Grasshoppers (Orthoptera, Acridoidea) from Mongolia. Biodivers. Data J. 2023, 11, e96705. [Google Scholar] [CrossRef] [PubMed]
- Bayartogtokh, B. Common Insects of Mongolia; Ministry of Environment, National Institute of Biological Resources (NIBR): Incheon, Republic of Korea, 2016. [Google Scholar]
- Dey, L.-S.; Seidel, M.; Davaa, L.; Husemann, M. From the Steppe to Desert: Survey of Band-Winged Grasshoppers from Mongolia (Orthoptera: Acrididae: Oedipodinae) Based on Material from 50 Years of Expeditions. Erforsch. Biol. Ressourcen Der Mong./Explor. Into Biol. Resour. Mong. 2021, 14, 329–360. [Google Scholar]
- Cigliano, M.M.; Braun, H.; Eades, H.; Otte, D. Orthoptera Species File. Version 5.0/5.0. Available online: http://orthoptera.speciesfile.org/ (accessed on 3 January 2024).
- Song, H.; Mariño-Pérez, R.; Woller, D.A.; Cigliano, M.M. Evolution, Diversification, and Biogeography of Grasshoppers (Orthoptera: Acrididae). Insect Syst. Divers. 2018, 2, 3. [Google Scholar] [CrossRef]
- Hawlitschek, O.; Morinière, J.; Lehmann, G.U.C.; Lehmann, A.W.; Kropf, M.; Dunz, A.; Glaw, F.; Detcharoen, M.; Schmidt, S.; Hausmann, A. DNA Barcoding of Crickets, Katydids and Grasshoppers (Orthoptera) from Central Europe with Focus on Austria, Germany and Switzerland. Mol. Ecol. Resour. 2017, 17, 1037–1053. [Google Scholar] [CrossRef] [PubMed]
- Husemann, M.; Sadílek, D.; Dey, L.-S.; Hawlitschek, O.; Seidel, M. New Genome Size Estimates for Band-Winged and Slant-Faced Grasshoppers (Orthoptera: Acrididae: Oedipodinae, Gomphocerinae) Reveal the so Far Largest Measured Insect Genome. Caryologia 2020, 73, 111–120. [Google Scholar]
- Hawlitschek, O.; Sadílek, D.; Dey, L.-S.; Buchholz, K.; Noori, S.; Baez, I.L.; Wehrt, T.; Brozio, J.; Trávníček, P.; Seidel, M.; et al. New Estimates of Genome Size in Orthoptera and their Evolutionary Implications. PLoS ONE 2023, 18, e0275551. [Google Scholar] [CrossRef]
- Stubbe, M.; Dawaa, N.; Hilbig, W.; Schamsran, Z. Die Entwicklung Der Zasammenarbeit Auf Biologischem Gebiet Zwischen Der MVR Und DDR Seit 1962. Erforsch. Biol. Ressourcen Der Mong./Explor. Into Biol. Resour. Mong. 1981, 1, 13–46. [Google Scholar]
- Günther, K.K. Blattoidea-Orthopteroidea-Ausbeute 1964, Teil II. (Tetrigidae Und Acrididae). Ergebnisse Der Mongolisch-Deutschen Biologischen Expeditionen Seit 1962, Nr. 55. Mitteilungen Aus Dem Mus. Für Naturkunde Berlin. Zool. Mus. Und Inst. Für Spez. Zool. 1971, 47, 109–130. [Google Scholar] [CrossRef]
- Hebert, P.D.; Gregory, T.R. The Promise of DNA Barcoding for Taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding Animal Life: Cytochrome c Oxidase Subunit 1 Divergences among Closely Related Species. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Kumar, H.; Tyagi, K.; Chandra, K.; Kumar, V. DNA Barcoding of Selected Short-Horned Grasshoppers (Orthoptera: Acrididae) from Indian Himalayan Region. Mitochondrial DNA Part B 2020, 5, 3600–3605. [Google Scholar] [CrossRef] [PubMed]
- De Jesús-Bonilla, V.S.; Barrientos-Lozano, L.; Zaldívar-Riverón, A. Sequence-Based Species Delineation and Molecular Phylogenetics of the Transitional Nearctic–Neotropical Grasshopper Genus Taeniopoda (Orthoptera, Romaleidae). Syst. Biodivers. 2017, 15, 600–617. [Google Scholar] [CrossRef]
- Gopurenko, D.; Fletcher, M.; Locker, H.; Mitchell, A. Morphological and DNA Barcode Species Identifications of Leafhoppers, Planthoppers and Treehoppers (Hemiptera: Auchenorrhyncha) at Barrow Island. Rec. WA Mus. 2013, 83, 253–285. [Google Scholar] [CrossRef]
- Trewick, S.A. DNA Barcoding Is Not Enough: Mismatch of Taxonomy and Genealogy in New Zealand Grasshoppers (Orthoptera: Acrididae). Cladistics 2008, 24, 240–254. [Google Scholar] [CrossRef]
- Moussi, A.; Dey, L.-S.; Petit, D.; Abba, A.; Klesser, R.; Husemann, M. First Genetic Data for Band-Winged Grasshoppers (Orthoptera: Acrididae: Oedipodinae) of the Biskra Region of Algeria with New Records for the Country. Afr. Zool. 2018, 53, 31–40. [Google Scholar] [CrossRef]
- Virgilio, M.; Backeljau, T.; Nevado, B.; De Meyer, M. Comparative Performances of DNA Barcoding across Insect Orders. BMC Bioinform. 2010, 11, 1–10. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Mistshenko, L. Orthoptera Palaearctica Critica: XII. Revision of Palaearctic Species of the Genus Sphingonotus Fieber (Orth. Acrid.).(Continua). Eos Rev. Española Entomol 1936, 12, 65–192. [Google Scholar]
- Storozhenko, S.Y.; Benediktov, A.A.; Huang, J. Revision of the Genus Angaracris Bey-Bienko, 1930 (Orthoptera: Acrididae, Oedipodinae) with Notes on the Tribe Bryodemini. Zootaxa 2017, 4244, 416. [Google Scholar] [CrossRef] [PubMed]
- Saboori, D.A.; Hodjat, S.H.; Tork, M.; Pahlow, F.; Husemann, M. A Faunistic Review of the Iranian Species of Sphingonotus (Orthoptera, Oedipodinae) with an Online Key to Species. Zootaxa 2018, 4379, 151–176. [Google Scholar]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a Medium for Simple Extraction of DNA for PCR-Based Typing from Forensic Material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef]
- Pan, C.; Hu, J.; Zhang, X.; Huang, Y. The DNA Barcoding Application of mtDNA COI Gene in Seven Species of Catantopidae (Orthoptera). Entomotaxonomia 2006, 28, 103–110. [Google Scholar]
- Huang, J.; Zhang, A.; Mao, S.; Huang, Y. DNA Barcoding and Species Boundary Delimitation of Selected Species of Chinese Acridoidea (Orthoptera: Caelifera). PLoS ONE 2013, 8, e82400. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Ze, S.; Liu, C.; Chen, B. New Insights into the Genetic Structure of the Outbreak-Prone Bamboo Grasshoppers. Front. Ecol. Evol. 2023, 11, 1062857. [Google Scholar] [CrossRef]
- Dey, L.-S.; Simões, M.V.; Hawlitschek, O.; Sergeev, M.G.; Xu, S.-Q.; Lkhagvasuren, D.; Husemann, M. Analysis of Geographic Centrality and Genetic Diversity in the Declining Grasshopper Species Bryodemella tuberculata (Orthoptera: Oedipodinae). Biodivers. Conserv. 2021, 30, 2773–2796. [Google Scholar] [CrossRef]
- Ivković, S.; Dey, L.-S.; Maria Buzzetti, F.; Puskás, G.; Warcha\lowska-Śliwa, E.; Horvat, L.; Chobanov, D.; Hochkirch, A. Strong Intraspecific Phylogenetic and Karyotypic Diversification in Isophya modestior (Orthoptera: Tettigoniidae: Phaneropterinae). Biol. J. Linn. Soc. 2023, 138, 194–203. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree (Version Version 1.4.4) Computer Software. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 8 April 2018).
- Inkscape Project Inkscape Computer Software. Available online: https://inkscape.org (accessed on 17 August 2021).
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Valdivia, P.; Rosas-Puchuri, U.; Valdivia, N.L. SPdel: A Pipeline to Compare and Visualize Species Delimitation Methods for Single-Locus Datasets. Mol. Ecol. Resour. 2023, 23, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Husemann, M.; Namkung, S.; Habel, J.C.; Danley, P.D.; Hochkirch, A. Phylogenetic Analyses of Band-winged Grasshoppers (Orthoptera, Acrididae, Oedipodinae) Reveal Convergence of Wing Morphology. Zool. Scr. 2012, 41, 515–526. [Google Scholar] [CrossRef]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many Species in One: DNA Barcoding Overestimates the Number of Species When Nuclear Mitochondrial Pseudogenes Are Coamplified. Proc. Natl. Acad. Sci. 2008, 105, 13486–13491. [Google Scholar] [CrossRef]
- Kasalo, N.; Skejo, J.; Husemann, M. DNA Barcoding of Pygmy Hoppers—The First Comprehensive Overview of the BOLD Systems’ Data Shows Promise for Species Identification. Diversity 2023, 15, 696. [Google Scholar] [CrossRef]
- Chen, L.-P.; Chang, Y.-G.; Li, J.; Wang, J.-M.; Liu, J.-L.; Zhi, Y.-C.; Li, X.-J. Application of DNA Barcoding in the Classification of Grasshoppers (Orthoptera: Acridoidea)—A Case Study of Grasshoppers from Hebei Province, China. Zootaxa 2018, 4497, 99–110. [Google Scholar] [CrossRef] [PubMed]
- DeSalle, R.; Goldstein, P. Review and Interpretation of Trends in DNA Barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef]
- O’hare, M.; Hugel, S.; Hendrickse, M.; Greyling, C.; Egan, B.; van Asch, B. Documenting the Biodiversity of Edible Grasshoppers in South Africa with Traditional Knowledge, Classic Taxonomy and Genetic Information. Biodivers. Conserv. 2023, 32, 3481–3502. [Google Scholar] [CrossRef]
- Pocco, M.E.; Minutolo, C.; Dinghi, P.A.; Lange, C.E.; Confalonieri, V.A.; Cigliano, M.M. Species Delimitation in the Andean Grasshopper Genus Orotettix Ronderos & Carbonell (Orthoptera: Melanoplinae): An Integrative Approach Combining Morphological, Molecular and Biogeographical Data. Zool. J. Linn. Soc. 2015, 174, 733–759. [Google Scholar]
- Song, H.; Moulton, M.J.; Whiting, M.F. Rampant Nuclear Insertion of mtDNA across Diverse Lineages within Orthoptera (Insecta). PLoS ONE 2014, 9, e110508. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.; Vukić, J.; Šanda, R.; Doadrio, I. Ancient Mitochondrial Capture as Factor Promoting Mitonuclear Discordance in Freshwater Fishes: A Case Study in the Genus Squalius (Actinopterygii, Cyprinidae) in Greece. PLoS ONE 2016, 11, e0166292. [Google Scholar] [CrossRef]
- Andersen, M.J.; McCullough, J.M.; Gyllenhaal, E.F.; Mapel, X.M.; Haryoko, T.; Jønsson, K.A.; Joseph, L. Complex Histories of Gene Flow and a Mitochondrial Capture Event in a Nonsister Pair of Birds. Mol. Ecol. 2021, 30, 2087–2103. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.W.; Devriese, H.; Tumbrinck, J.; Skejo, J.; Lehmann, G.U.; Hochkirch, A. The Importance of Validated Alpha Taxonomy for Phylogenetic and DNA Barcoding Studies: A Comment on Species Identification of Pygmy Grasshoppers (Orthoptera, Tetrigidae). ZooKeys 2017, 679, 139. [Google Scholar] [CrossRef] [PubMed]



| 1 | Bryodema heptapotanicum |
| 2 | Bryodema nigripennis |
| 3 | Bryodemella orientale orientale |
| 4 | Bryodemella semenovi |
| 5 | Bryodemella zaisanicum fallax |
| 6 | Celes skalozubovi |
| 7 | Epacromius tergestinus tergestinus |
| 8 | Leptopternis iliensis |
| 9 | Sphingonotus lucidus |
| 10 | Sphingonotus halophilus |
| ID | Species | Origin | Original ID |
|---|---|---|---|
| N1 | Bryodema kozlovi | NCBI | NC_052731 |
| N2 | Bryodema miramae miramae | NCBI | KP889242 |
| N3 | Compsorhipis davidiana | NCBI | NC_029408 |
| N4 | Compsorhipis davidiana | NCBI | KT157830 |
| N5 | Epacromius pulverulentus | NCBI | MT129326 |
| N6 | Helioscirtus moseri | NCBI | KR005923 |
| N7 | Stethophyma grossum | NCBI | GU706160 |
| N8 | Stethophyma grossum | NCBI | GU706136 |
| N9 | Sphingoderus carinatus | NCBI | MK251002 |
| N10 | Sphingoderus carinatus | NCBI | MK250997 |
| N11 | Psophus stridulus | NCBI | MT311126 |
| N12 | Psophus stridulus | NCBI | HQ955713 |
| N13 | Psophus stridulus | NCBI | GU706161 |
| N14 | Oedaleus infernalis | NCBI | KC297217 |
| N15 | Oedaleus infernalis | NCBI | NC_029327 |
| N16 | Celes variabilis | BOLD | GBMH5075_08 |
| N17 | Celes variabilis | BOLD | GBORT942_15 |
| N18 | Sphingonotus coerulipes | BOLD | GBMH5086_08 |
| N32 | Aiolopus thalassinus | NCBI | OQ214016.1 |
| N33 | Aiolopus thalassinus | NCBI | OQ214015.1 |
| N34 | Locusta migratoria | NCBI | OQ214122.1 |
| N35 | Locusta migratoria | NCBI | OQ214121.1 |
| N40 | Podismopsis altaica | NCBI | AY738343.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kock, L.-S.; Körs, E.; Husemann, M.; Davaa, L.; Dey, L.-S. Barcoding Fails to Delimit Species in Mongolian Oedipodinae (Orthoptera, Acrididae). Insects 2024, 15, 128. https://doi.org/10.3390/insects15020128
Kock L-S, Körs E, Husemann M, Davaa L, Dey L-S. Barcoding Fails to Delimit Species in Mongolian Oedipodinae (Orthoptera, Acrididae). Insects. 2024; 15(2):128. https://doi.org/10.3390/insects15020128
Chicago/Turabian StyleKock, Lea-Sophie, Elisabeth Körs, Martin Husemann, Lkhagvasuren Davaa, and Lara-Sophie Dey. 2024. "Barcoding Fails to Delimit Species in Mongolian Oedipodinae (Orthoptera, Acrididae)" Insects 15, no. 2: 128. https://doi.org/10.3390/insects15020128
APA StyleKock, L. -S., Körs, E., Husemann, M., Davaa, L., & Dey, L. -S. (2024). Barcoding Fails to Delimit Species in Mongolian Oedipodinae (Orthoptera, Acrididae). Insects, 15(2), 128. https://doi.org/10.3390/insects15020128