
Spatial and Temporal Variations in Richness, Diversity and Abundance of Floral Visitors of Curry Plants (Bergera koenigii L.): Insights on Plant-Pollinator Interactions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Species
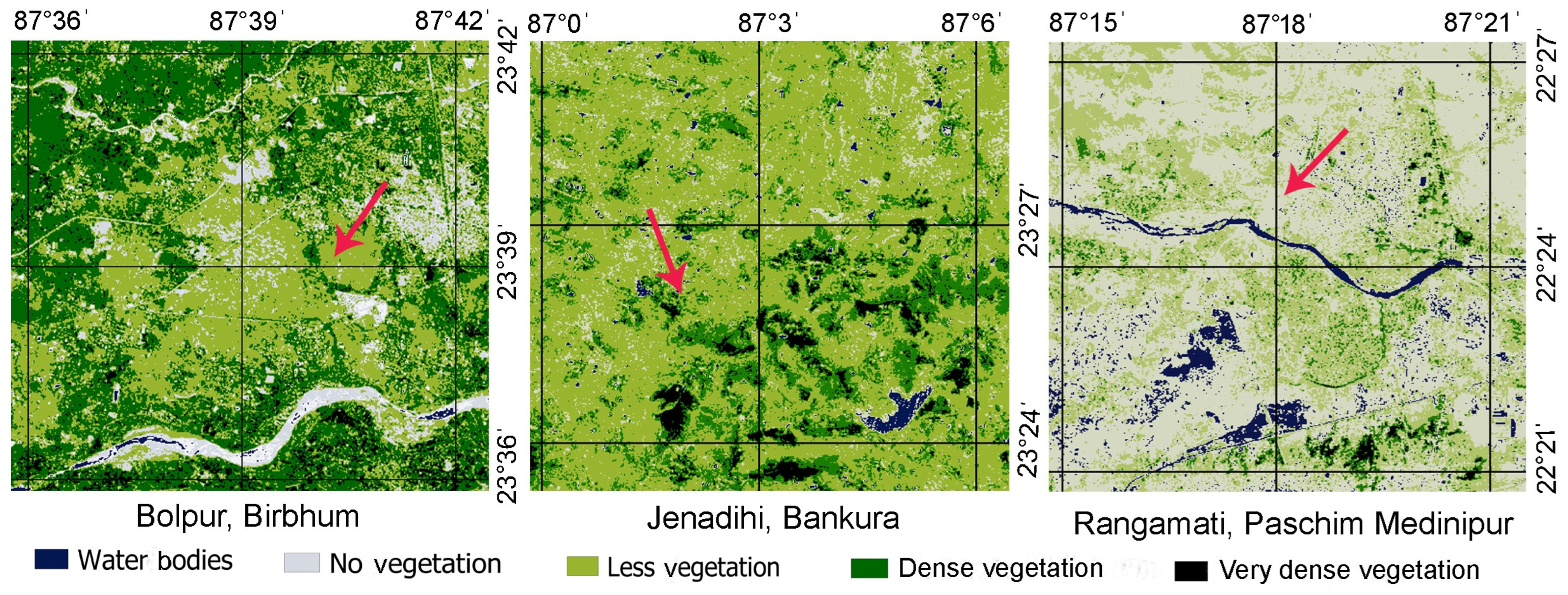
2.2. Study Sites and Vegetation Patterns
2.3. Data Collection about Floral Visitors
2.4. Data Analysis
3. Results
3.1. Vegetation Patterns in the Three Selected Zones
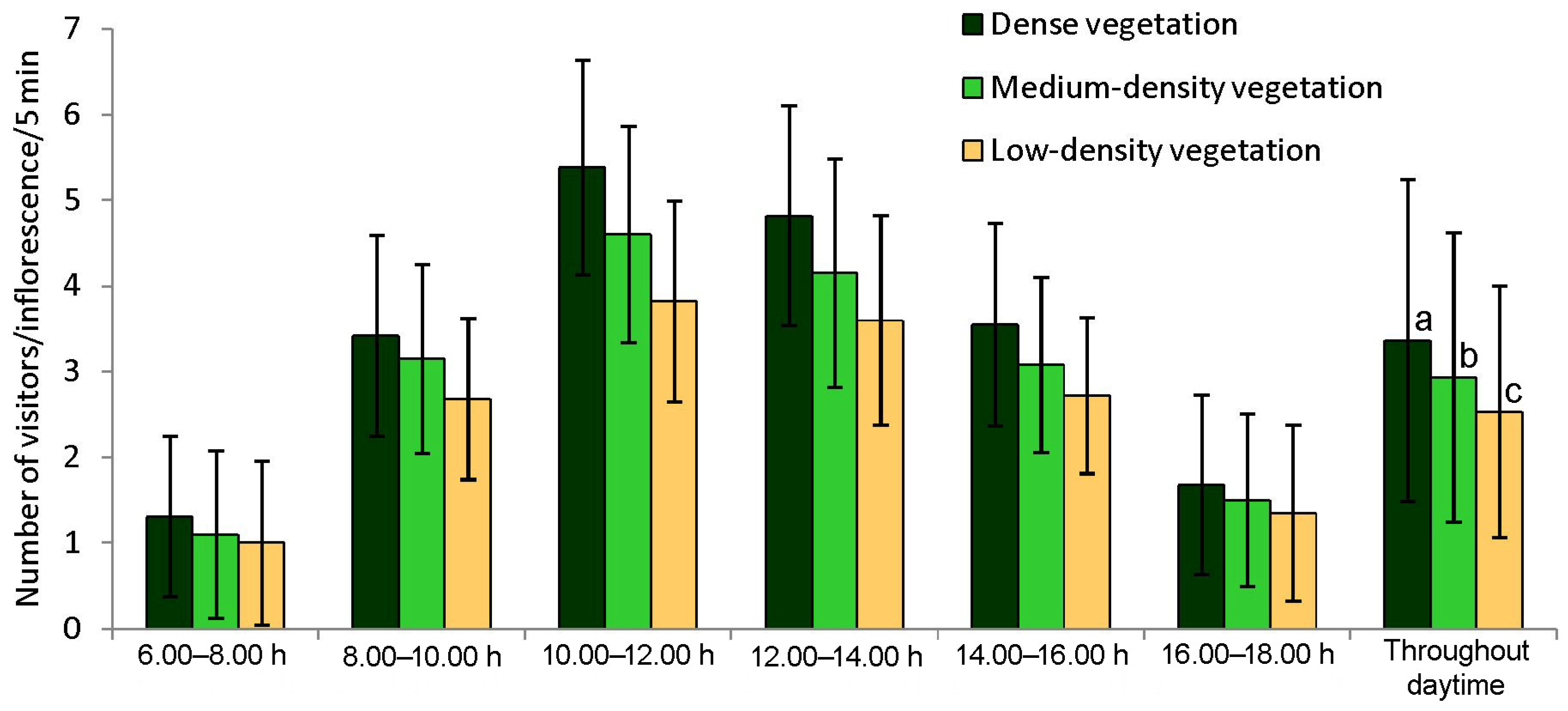
3.2. Floral Visitors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Robertson, C. The philosophy of flower seasons, and the phaenological relations of the entomophilous flora and the anthophilous insect fauna. Am. Nat. 1895, 29, 97–117. [Google Scholar] [CrossRef]
- Sargent, R.D.; Ackerly, D.D. Plant–pollinator interactions and the assembly of plant communities. Trends Ecol. Evol. 2008, 23, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.D.; Peakall, R.; van der Niet, T.; Johnson, S.D. Niche perspectives on plant–pollinator interactions. Trends Plant Sci. 2020, 25, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Moeller, D.A. Pollinator community structure and sources of spatial variation in plant–pollinator interactions in Clarkia xantiana ssp. xantiana. Oecologia 2005, 142, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, S.A.; Bronstein, J.L.; Rudgers, J.A. How context dependent are species interactions? Ecol. Lett. 2014, 17, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Chapin III, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef]
- Rogers, S.R.; Tarpy, D.R.; Burrack, H.J. Bee species diversity enhances productivity and stability in a perennial crop. PLoS ONE 2014, 9, e97307. [Google Scholar] [CrossRef]
- Brown, M.J.; Paxton, R.J. The conservation of bees: A global perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Fortuna, M.A.; Bascompte, J. Habitat loss and the structure of plant–animal mutualistic networks. Ecol. Lett. 2006, 9, 281–286. [Google Scholar] [CrossRef]
- Weiner, C.N.; Werner, M.; Linsenmair, K.E.; Blüthgen, N. Land-use impacts on plant–pollinator networks: interaction strength and specialization predict pollinator declines. Ecology 2014, 95, 466–474. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Laliberté, E.; Nielsen, A.; Bascompte, J. Conservation of species interaction networks. Biol. Conserv. 2010, 143, 2270–2279. [Google Scholar] [CrossRef]
- Kaiser-Bunbury, C.N.; Blüthgen, N. Integrating network ecology with applied conservation: a synthesis and guide to implementation. AoB Plants 2015, 7, plv076. [Google Scholar] [CrossRef]
- Layek, U.; Das, A.; Karmakar, P. Supplemental stingless bee pollination in fennel (Foeniculum vulgare Mill.): An assessment of impacts on native pollinators and crop yield. Front. Sustain. Food Syst. 2022, 6, 820264. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Rathore, L.S.; Vishnoi, G.; Khan, A.U.; Kumar, B. Record of insect pollinators on the inflorescence of Murraya koenigii in Rajasthan. J. Agric. Ecol. 2018, 5, 38–41. [Google Scholar] [CrossRef]
- Dhore, M.M. Diversity and foraging behavior of insect pollinators on Murraya koenigii (L.) (Rutaceae). Int. J. Sci. Res. Sci. Technol. 2020, 7, 15–19. [Google Scholar]
- Layek, U.; Das, A.; Das, U. Floral biology, floral volatile organic compounds and floral visitors of Chromolaena odorata, an invasive alien species in West Bengal, India. Biodiversitas 2022, 23, 2118–2129. [Google Scholar] [CrossRef]
- Subedi, B.; Stewart, A.B.; Neupane, B.; Ghimire, S.; Adhikari, H. Butterfly species diversity and their floral preferences in the Rupa Wetland of Nepal. Ecol. Evol. 2021, 11, 2086–2099. [Google Scholar] [CrossRef]
- Corbet, S.A. Butterfly nectaring flowers: Butterfly morphology and flower form. Entomol. Exp. Et Appl. 2000, 96, 289–298. [Google Scholar] [CrossRef]
- Layek, U.; Bera, K.; Bera, B.; Bisui, S.; Pattanayek, S.K.; Karmakar, P. Assessment of yield enhancement in cashew (Anacardium occidentale L.) by the pollinator sharing effect of magnetic bee-friendly plants in India. Acta Ecol. Sin. 2021, 41, 243–252. [Google Scholar] [CrossRef]
- Gilpin, A.M.; Kobel, C.; Brettell, L.E.; O’Brien, C.; Cook, J.M.; Power, S.A. Co-flowering species richness increases pollinator visitation to apple flowers. Agriculture 2022, 12, 1246. [Google Scholar] [CrossRef]
- Brittain, C.; Williams, N.; Kremen, C.; Klein, A.M. Synergistic effects of non-Apis bees and honey bees for pollination services. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122767. [Google Scholar] [CrossRef]
- Balfour, N.J.; Gandy, S.; Ratnieks, F.L. Exploitative competition alters bee foraging and flower choice. Behav. Ecol. Sociobiol. 2015, 69, 1731–1738. [Google Scholar] [CrossRef]
- Raju, A.J.S.; Ramana, K.V.; Lakshmi, P.V. Wendlandia tinctoria (Roxb.) DC. (Rubiaceae), a key nectar source for butterflies during the summer season in the southern Eastern Ghats, Andhra Pradesh, India. J. Threat. Taxa 2011, 3, 1594–1600. [Google Scholar] [CrossRef]
- Mayer, C.; Michez, D.; Chyzy, A.; Brédat, E.; Jacquemart, A.L. The abundance and pollen foraging behaviour of bumble bees in relation to population size of whortleberry (Vaccinium uliginosum). PLoS ONE 2012, 7, e50353. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Jeon, H.; Jung, C. Foraging behaviour and preference of pollen sources by honey bee (Apis mellifera) relative to protein contents. J. Ecol. Environ. 2020, 44, 4. [Google Scholar] [CrossRef]
- Layek, U.; Manna, S.S.; Karmakar, P. Pollen foraging behaviour of honey bee (Apis mellifera L.) in southern West Bengal, India. Palynology 2020, 44, 114–126. [Google Scholar] [CrossRef]
- Cruden, R.W.; Hermann-Parker, S.M. Butterfly pollination of Caesalpinia pulcherrima, with observations on a psychophilous syndrome. J. Ecol. 1979, 67, 155–168. [Google Scholar] [CrossRef]
- Goldblatt, P.; Manning, J.C. Evidence for moth and butterfly pollination in Gladiolus (Iridaceae-Crocoideae). Ann. Mo. Bot. Gard. 2002, 89, 110–124. [Google Scholar] [CrossRef]
- Balducci, M.G.; Van der Niet, T.; Johnson, S.D. Butterfly pollination of Bonatea cassidea (Orchidaceae): Solving a puzzle from the Darwin era. S. Afr. J. Bot. 2019, 123, 308–316. [Google Scholar] [CrossRef]
- Southwick, E.E.; Southwick, L., Jr. Estimating the economic value of honey bees (Hymenoptera: Apidae) as agricultural pollinators in the United States. J. Econ. Entomol. 1992, 85, 621–633. [Google Scholar] [CrossRef]
- Stanley, D.A.; Msweli, S.M.; Johnson, S.D. Native honeybees as flower visitors and pollinators in wild plant communities in a biodiversity hotspot. Ecosphere 2020, 11, e02957. [Google Scholar] [CrossRef]
- Artz, D.R.; Allan, M.J.; Wardell, G.I.; Pitts-Singer, T.L. Nesting site density and distribution affect Osmia lignaria (Hymenoptera: Megachilidae) reproductive success and almond yield in a commercial orchard. Insect Conserv. Divers. 2013, 6, 715–724. [Google Scholar] [CrossRef]
- Santos, A.O.R.; Bartelli, B.F.; Nogueira-Ferreira, F.H. Potential pollinators of tomato, Lycopersicon esculentum (Solanaceae), in open crops and the effect of a solitary bee in fruit set and quality. J. Econ. Entomol. 2014, 107, 987–994. [Google Scholar] [CrossRef]
- Heard, T.A. Behaviour and pollinator efficiency of stingless bees and honey bees on macadamia flowers. J. Apic. Res. 1994, 33, 191–198. [Google Scholar] [CrossRef]
- Bisui, S.; Layek, U.; Karmakar, P. Utilization of Indian dammar bee (Tetragonula iridipennis Smith) as a pollinator of bitter gourd. Acta Agrobot. 2020, 73, 7316. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Insect Order | Family | Insect Species |
|---|---|---|
| Diptera | Rhiniidae | Stomorhina discolor *# |
| Stratiomyidae | Oplodontha viridula | |
| Hymenoptera | Apidae | Amegilla zonata, Apis cerana, Apis dorsata, Apis florea ∆, Ceratina binghami, Ceratina compacta, Tetragonula iridipennis, Thyreus nitidulus #∆ |
| Halictidae | Halictus acrocephalus, Lasioglossum funebre, Nomia iridescens ∆, Sphecodes gibbus #∆ | |
| Scoliidae | Scolia soror ∆ | |
| Lepidoptera | Erebidae | Syntomoides imaon |
| Hesperiidae | Ancistroides folus #∆, Baoris farri, Suastus gremius, and Telicota colon | |
| Lycaenidae | Anthene lycaenina #∆, Catochrysops strato ∆, Chilades lajus #∆, Chilades pandava ∆, Jamides bochus #∆, Rapala manea, Rapala varuna, and Tarucus indica #∆ | |
| Nymphalidae | Danaus chrysippus, Danaus genutia #∆, Euploea core, Junonia almana #∆, Junonia atlites, Junonia iphita, Mycalesis perseus #∆, Tirumala limniace | |
| Papilionidae | Pachliopta hector, Papilio demoleus #∆, Papilio polytes, | |
| Pieridae | Appias libythea, Catopsilia pomona, Eurema blanda, Eurema hecabe, Leptosia nina, and Pareronia hippie |
| Daytime | Values of D | ||
|---|---|---|---|
| Dense Vegetation | Medium-Density Vegetation | Low-Density Vegetation | |
| 6.00–8.00 h | 0.35 c ± 0.64 | 0.33 c ± 0.60 | 0.21 b ± 0.48 |
| 8.00–10.00 h | 1.06 b ± 0.63 | 0.86 ab ± 0.56 | 0.63 a ± 0.63 |
| 10.00–12.00 h | 1.47 a ± 0.52 | 1.12 a ± 0.47 | 0.86 a ± 0.64 |
| 12.00–14.00 h | 1.35 a ± 0.53 | 1.06 a ± 0.52 | 0.91 a ± 0.58 |
| 14.00–16.00 h | 0.93 b ± 0.58 | 0.77 b ± 0.62 | 0.67 a ± 0.61 |
| 16.00–18.00 h | 0.45 c ± 0.64 | 0.47 c ± 0.68 | 0.33 b ± 0.60 |
| Overall | 0.94 a ± 0.72 | 0.77 b ± 0.64 | 0.60 c ± 0.64 |
| Daytime | Values of H’ | ||
|---|---|---|---|
| Dense Vegetation | Medium-Density Vegetation | Low-Density Vegetation | |
| 6.00–8.00 h | 0.22 c ± 0.36 | 0.18 d ± 0.32 | 0.16 c ± 0.31 |
| 8.00–10.00 h | 0.83 b ± 0.36 | 0.72 c ± 0.43 | 0.58 b ± 0.40 |
| 10.00–12.00 h | 1.16 a ± 0.24 | 1.11 a ± 0.21 | 0.87 a ± 0.30 |
| 12.00–14.00 h | 1.09 a ± 0.31 | 0.96 b ± 0.28 | 0.80 a ± 0.35 |
| 14.00–16.00 h | 0.83 b ± 0.33 | 0.78 c ± 0.33 | 0.54 b ± 0.41 |
| 16.00–18.00 h | 0.33 c ± 0.39 | 0.28 d ± 0.39 | 0.21 c ± 0.33 |
| Overall | 0.74 a ± 0.48 | 0.67 a ± 0.47 | 0.53 b ± 0.44 |
| Floral Visitors | Flower Visitation Rate | Flower Handling Time (s) | ||||
|---|---|---|---|---|---|---|
| Dense Vegetation | Medium-Density Vegetation | Low-Density Vegetation | Dense Vegetation | Medium-Density Vegetation | Low-Density Vegetation | |
| ||||||
| Oplodontha viridula | - | - | - | - | - | - |
| Stomorhina discolor | - | - | - | - | - | - |
| ||||||
| Amegilla zonata | 14.35 ± 3.47 | 14.20 ± 3.25 | 13.95 ± 3.03 | 1.42 ± 0.84 | 1.48 ± 0.85 | 1.54 ± 0.93 |
| Apis cerana | 11.05 a ± 2.49 | 9.90 b ± 2.40 | 9.12 c ± 2.43 | 3.66 c ± 1.15 | 4.03 b ± 1.24 | 4.32 a ± 1.41 |
| Apis dorsata | 19.53 a ± 3.68 | 16.10 b ± 4.43 | 14.98 c ± 3.81 | 1.39 c ± 0.59 | 1.59 b ± 0.51 | 1.81 a ± 0.54 |
| Apis florea | 12.07 ± 2.24 | 10.87 ± 2.33 | - | 3.43 ± 0.74 | 3.89 ± 0.96 | - |
| Ceratina binghami | 8.08 a ± 2.50 | 6.87 b ± 2.27 | 5.83 c ± 2.02 | 7.45 ± 3.87 | 8.07 ± 4.13 | 8.30 ± 4.32 |
| Ceratina compacta | 8.60 a ± 1.67 | 6.97 b ± 2.14 | 6.23 c ± 2.11 | 7.04 c ± 3.62 | 7.76 b ± 3.75 | 8.23 a ± 3.91 |
| Halictus acrocephalus | 8.92 a ± 2.49 | 6.82 b ± 2.27 | 5.35 c ± 2.15 | 6.22 c ± 2.50 | 7.85 b ± 3.33 | 9.55 a ± 3.67 |
| Lasioglossum funebre | 7.93 a ± 1.78 | 6.87 b ± 1.99 | 6.27 b ± 1.93 | 5.47 c ± 2.48 | 6.32 b ± 2.84 | 7.26 a ± 3.33 |
| Nomia iridescens | 11.92 ± 2.78 | 10.28 ± 2.90 | - | 5.44 ± 3.09 | 6.25 ± 3.69 | - |
| Scolia soror | 7.23 ± 1.94 | 7.07 ± 1.91 | - | 3.28 ± 0.83 | 3.72 ± 0.98 | - |
| Sphecodes gibbus | 11.22 ± 1.85 | - | - | 3.48 ± 0.86 | - | - |
| Tetragonula iridipennis | 2.95 a ± 1.03 | 2.30 b ± 0.98 | 1.88 c ± 0.80 | 11.76 c ± 3.71 | 14.03 b ± 4.04 | 16.34 a ± 4.40 |
| Thyreus nitidulus | 13.80 ± 3.27 | - | - | 3.17 ± 0.74 | - | - |
| ||||||
| Ancistroides folus | 3.02 ± 1.17 | - | - | 9.84 ± 6.12 | - | - |
| Anthene lycaenina | 2.15 ± 0.97 | - | - | 12.15 ± 8.37 | - | - |
| Appias libythea | 5.62 a ± 1.75 | 4.55 b ± 1.59 | 4.07 b ± 1.45 | 4.83 c ± 2.10 | 6.02 b ± 2.18 | 7.07 a ± 2.23 |
| Baoris farri | 3.25 a ± 1.10 | 3.00 b ± 1.02 | 2.83 c ± 0.95 | 7.83 c ± 5.36 | 9.75 b ± 5.92 | 11.47 a ± 5.98 |
| Catochrysops strato | 2.33 ± 1.06 | 2.03 ± 0.76 | - | 11.78 ± 8.07 | 13.37 ± 8.43 | - |
| Catopsilia pomona | 5.10 a ± 1.47 | 4.37 b ± 1.47 | 4.17 c ± 1.42 | 6.44 c ± 4.34 | 7.12 b ± 5.04 | 7.68 a ± 5.93 |
| Chilades lajus | 2.13 ± 0.97 | - | - | 11.85 ± 8.07 | - | - |
| Chilades pandava | 2.35 ± 0.99 | 2.03 ± 0.93 | - | 11.43 ± 7.84 | 13.57 ± 8.21 | - |
| Danaus chrysippus | 4.37 ± 1.61 | 4.10 ± 1.32 | 4.03 ± 1.27 | 5.39 ± 4.47 | 6.43 ± 5.04 | 7.88 ± 5.62 |
| Danaus genutia | 4.23 ± 1.68 | - | - | 5.12 ± 4.38 | - | - |
| Euploea core | 4.63 a ± 1.61 | 4.25 b ± 1.63 | 4.10 c ± 1.42 | 6.37 c ± 4.96 | 7.36 b ± 5.72 | 8.28 a ± 6.45 |
| Eurema blanda | 3.87 a ± 1.66 | 3.63 b ± 1.61 | 3.37 c ± 1.47 | 5.81 c ± 3.82 | 6.82 b ± 4.65 | 7.79 a ± 5.43 |
| Eurema hecabe | 4.12 a ± 1.63 | 3.83 b ± 1.51 | 3.53 c ± 1.28 | 5.46 c ± 3.65 | 6.54 b ± 4.39 | 7.42 a ± 5.22 |
| Jamides bochus | 2.23 ± 0.90 | - | - | 11.96 ± 8.22 | - | - |
| Junonia almana | 3.68 ± 1.42 | - | - | 5.38 ± 3.87 | - | - |
| Junonia atlites | 4.37 a ± 1.63 | 3.52 b ± 1.20 | 3.25 c ± 1.19 | 6.73 c ± 4.86 | 7.94 b ± 5.79 | 9.12 a ± 6.66 |
| Junonia iphita | 2.40 ± 1.07 | 2.13 ± 0.94 | 2.03 ± 0.96 | 11.04 ± 7.49 | 13.53 ± 7.72 | 15.16 ± 7.93 |
| Leptosia nina | - | - | - | - | - | - |
| Mycalesis perseus | 2.03 ± 0.96 | - | - | 10.83 ± 7.84 | - | - |
| Pachliopta hector | 6.53 a ± 2.30 | 5.90 b ± 1.58 | 5.47 c ± 1.59 | 4.71 c ± 2.28 | 5.34 b ± 2.57 | 5.82 a ± 2.81 |
| Papilio demoleus | 6.70 ± 1.95 | - | - | 4.78 ± 2.19 | - | - |
| Papilio polytes | 6.13 ± 1.87 | 5.93 ± 1.68 | 5.78 ± 1.65 | 4.95 ± 2.24 | 5.62 ± 2.43 | 6.18 ± 2.67 |
| Pareronia hippie | 4.17 a ± 1.66 | 3.20 b ± 1.13 | 2.70 c ± 1.12 | 6.02 c ± 4.62 | 7.28 b ± 5.53 | 8.40 a ± 6.44 |
| Rapala manea | 2.28 a ± 0.94 | 1.97 b ± 0.89 | 1.83 c ± 0.83 | 12.27 c ± 8.19 | 13.71 b ± 8.73 | 15.25 a ± 9.28 |
| Rapala varuna | 2.47 a ± 1.01 | 2.17 b ± 0.91 | 1.90 c ± 0.84 | 11.73 c ± 7.94 | 13.26 b ± 8.54 | 14.83 a ± 8.92 |
| Suastus gremius | 3.15 a ± 1.09 | 2.77 b ± 1.14 | 2.25 c ± 0.98 | 9.96 c ± 5.72 | 12.46 b ± 8.06 | 14.42 a ± 10.12 |
| Syntomoides imaon | 11.60 ± 2.55 | 11.73 ± 2.72 | 11.93 ± 2.80 | 2.46 ± 0.61 | 2.68 ± 0.68 | 2.93 ± 0.73 |
| Tarucus indica | 1.95 ± 0.85 | - | - | 12.28 ± 8.23 | - | - |
| Telicota colon | 3.30 a ± 1.12 | 2.80 b ± 1.21 | 2.47 c ± 1.11 | 7.86 c ± 4.72 | 9.91 b ± 5.77 | 12.28 a ± 7.50 |
| Tirumala limniace | 5.68 a ± 1.86 | 5.27 b ± 1.60 | 4.90 c ± 1.37 | 5.87 c ± 3.30 | 6.54 b ± 3.99 | 7.28 a ± 5.22 |
| Floral Visitors | Pollen-Carrying Value | APV | ||||
|---|---|---|---|---|---|---|
| PCV1 | PCV2 | PCV | Dense Vegetation | Medium-Density Vegetation | Low-Density Vegetation | |
| ||||||
| Oplodontha viridula | - | - | - | - | - | - |
| Stomorhina discolor | - | - | - | - | - | - |
| ||||||
| Amegilla zonata | 1 | 0.5 | 1.5 | 26.69 | 36.42 | 34.53 |
| Apis cerana | 1 | 1 | 2 | 46.63 | 56.23 | 159.24 |
| Apis dorsata | 1 | 0.5 | 1.5 | 163.47 | 72.21 | 118.42 |
| Apis florea | 0.5 | 0.5 | 1 | 22.45 | 15.44 | - |
| Ceratina binghami | 0.5 | 0.5 | 1 | 12.04 | 5.84 | 4.78 |
| Ceratina compacta | 0.5 | 0.5 | 1 | 13.85 | 16.87 | 15.39 |
| Halictus acrocephalus | 0.5 | 1 | 1.5 | 121.22 | 181.89 | 136.18 |
| Lasioglossum funebre | 0.5 | 0.5 | 1 | 23.63 | 16.63 | 16.55 |
| Nomia iridescens | 1 | 0 | 1 | 208.48 | 90.67 | - |
| Scolia soror | - | - | - | - | - | - |
| Sphecodes gibbus | - | - | - | - | - | - |
| Tetragonula iridipennis | 0.5 | 1 | 1.5 | 26.37 | 52.03 | 57.61 |
| Thyreus nitidulus | 0.5 | 0 | 0.5 | 6.83 | - | - |
| ||||||
| Ancistroides folus | - | - | - | - | - | - |
| Anthene lycaenina | - | - | - | - | - | - |
| Appias libythea | 0.5 | 0 | 0.5 | 48.46 | 15.22 | 14.75 |
| Baoris farri | - | - | - | - | - | - |
| Catochrysops strato | - | - | - | - | - | - |
| Catopsilia pomona | 0.5 | 0 | 0.5 | 10.43 | 13.68 | 14.07 |
| Chilades lajus | - | - | - | - | - | - |
| Chilades pandava | - | - | - | - | - | - |
| Danaus chrysippus | 0.5 | 0 | 0.5 | 4.35 | 5.82 | 4.65 |
| Danaus genutia | - | - | - | - | - | - |
| Euploea core | 0.5 | 0 | 0.5 | 3.73 | 4.23 | 4.39 |
| Eurema blanda | 0.5 | 0 | 0.5 | 2.40 | 3.10 | 2.22 |
| Eurema hecabe | 0.5 | 0 | 0.5 | 2.31 | 2.45 | 2.03 |
| Jamides bochus | - | - | - | - | - | - |
| Junonia almana | - | - | - | - | - | - |
| Junonia atlites | 0.5 | 0 | 0.5 | 4.06 | 4.51 | 4.55 |
| Junonia iphita | - | - | - | - | - | - |
| Leptosia nina | - | - | - | - | - | - |
| Mycalesis perseus | - | - | - | - | - | - |
| Pachliopta hector | - | - | - | - | - | - |
| Papilio demoleus | - | - | - | - | - | - |
| Papilio polytes | - | - | - | - | - | - |
| Pareronia hippie | 0.5 | 0 | 0.5 | 3.11 | 3.18 | 2.67 |
| Rapala manea | - | - | - | - | - | - |
| Rapala varuna | - | - | - | - | - | - |
| Suastus gremius | 0.5 | 0 | 0.5 | 3.32 | 3.74 | 2.97 |
| Syntomoides imaon | 0.5 | 0 | 0.5 | 3.60 | 10.03 | 13.78 |
| Tarucus indica | - | - | - | - | - | - |
| Telicota colon | - | - | - | - | - | - |
| Tirumala limniace | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Layek, U.; Das, A.D.; Das, U.; Karmakar, P. Spatial and Temporal Variations in Richness, Diversity and Abundance of Floral Visitors of Curry Plants (Bergera koenigii L.): Insights on Plant-Pollinator Interactions. Insects 2024, 15, 83. https://doi.org/10.3390/insects15020083
Layek U, Das AD, Das U, Karmakar P. Spatial and Temporal Variations in Richness, Diversity and Abundance of Floral Visitors of Curry Plants (Bergera koenigii L.): Insights on Plant-Pollinator Interactions. Insects. 2024; 15(2):83. https://doi.org/10.3390/insects15020083
Chicago/Turabian StyleLayek, Ujjwal, Anirban Deep Das, Uday Das, and Prakash Karmakar. 2024. "Spatial and Temporal Variations in Richness, Diversity and Abundance of Floral Visitors of Curry Plants (Bergera koenigii L.): Insights on Plant-Pollinator Interactions" Insects 15, no. 2: 83. https://doi.org/10.3390/insects15020083
APA StyleLayek, U., Das, A. D., Das, U., & Karmakar, P. (2024). Spatial and Temporal Variations in Richness, Diversity and Abundance of Floral Visitors of Curry Plants (Bergera koenigii L.): Insights on Plant-Pollinator Interactions. Insects, 15(2), 83. https://doi.org/10.3390/insects15020083