
Association of a Global Invasive Pest Spodoptera frugiperda (Lepidoptera: Noctuidae) with Local Parasitoids: Prospects for a New Approach in Selecting Biological Control Agents


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
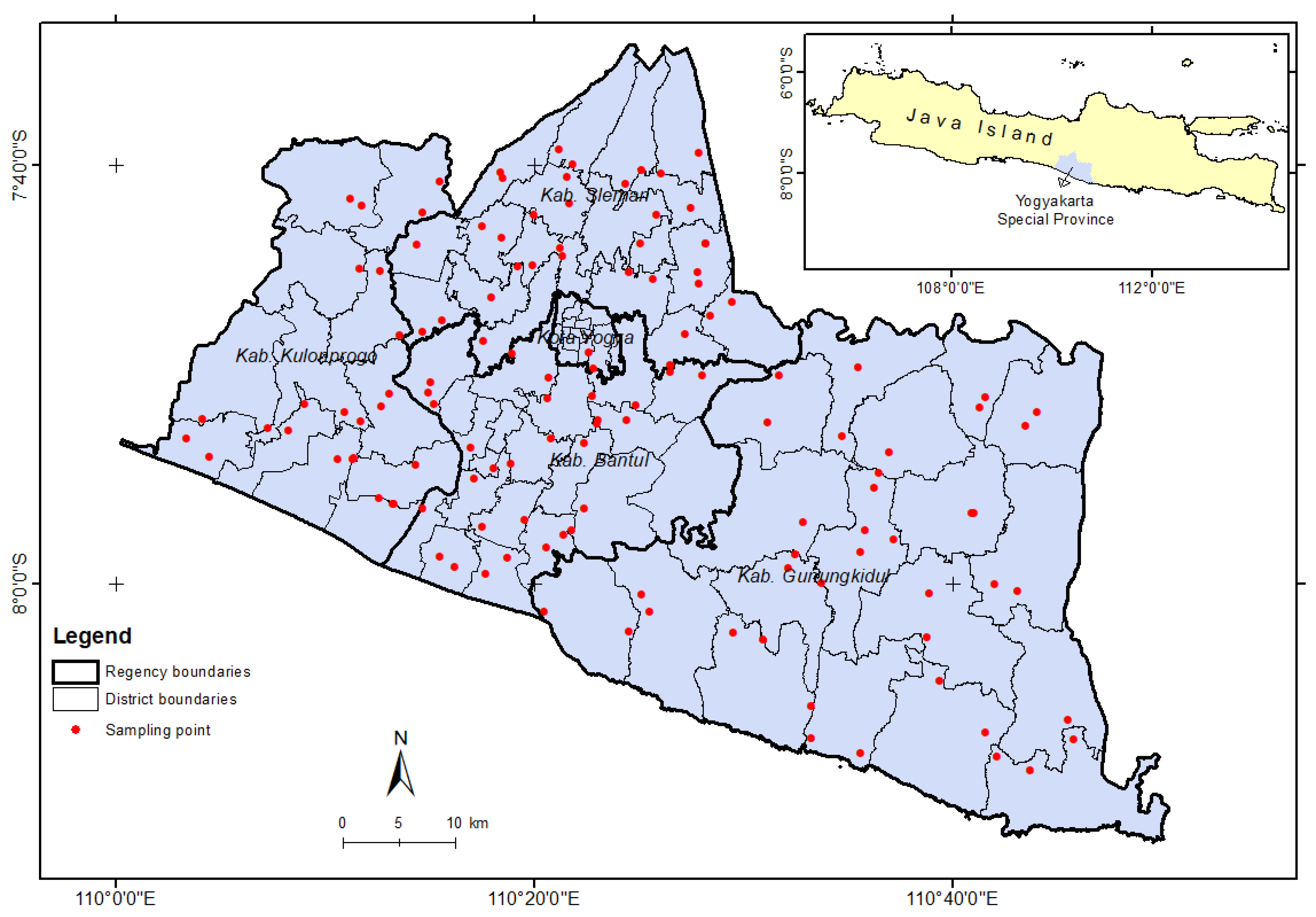
2.1. Sampling Location Determination
2.2. Sampling
2.3. Parasitoid Identification
2.4. Data Analysis
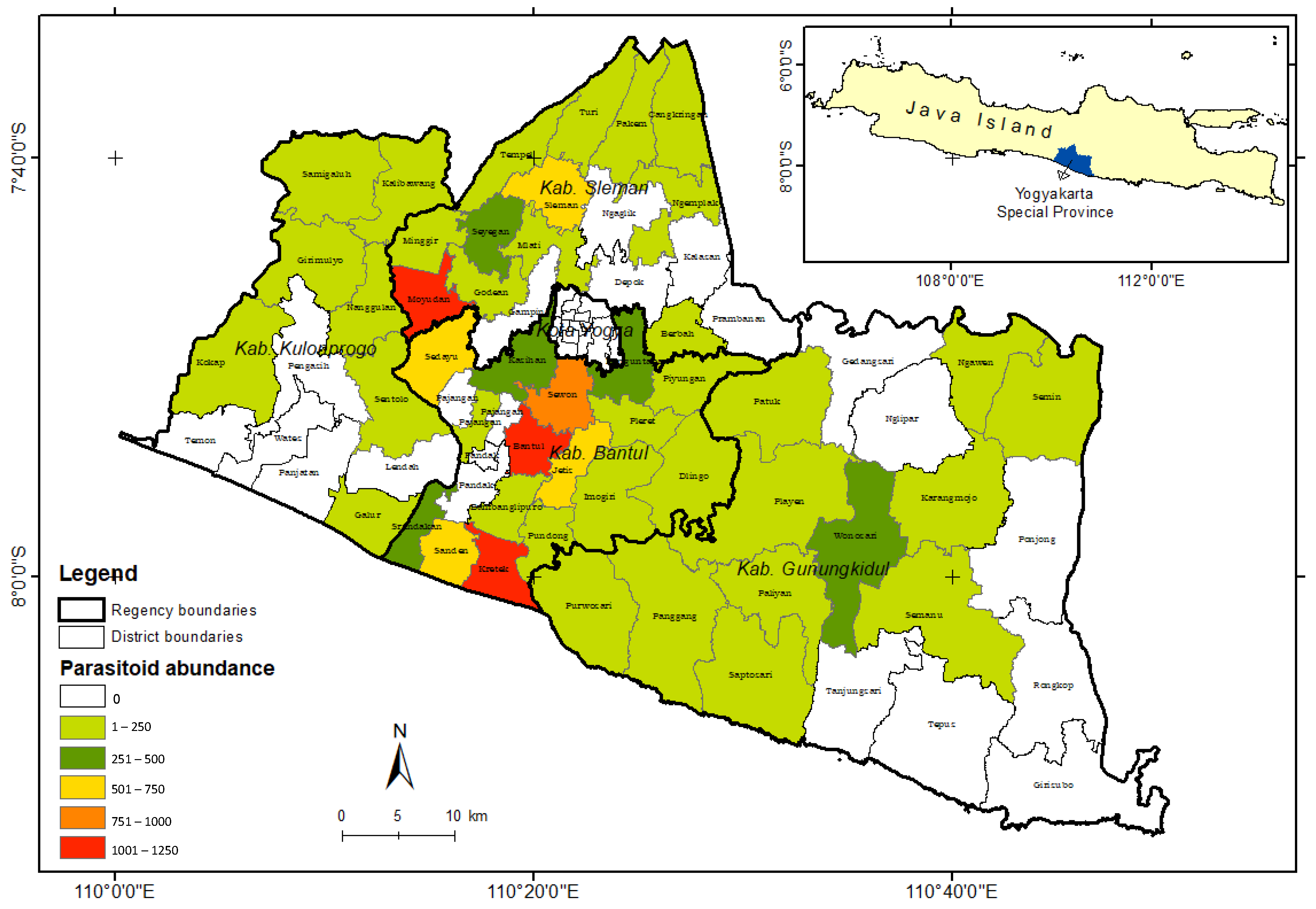
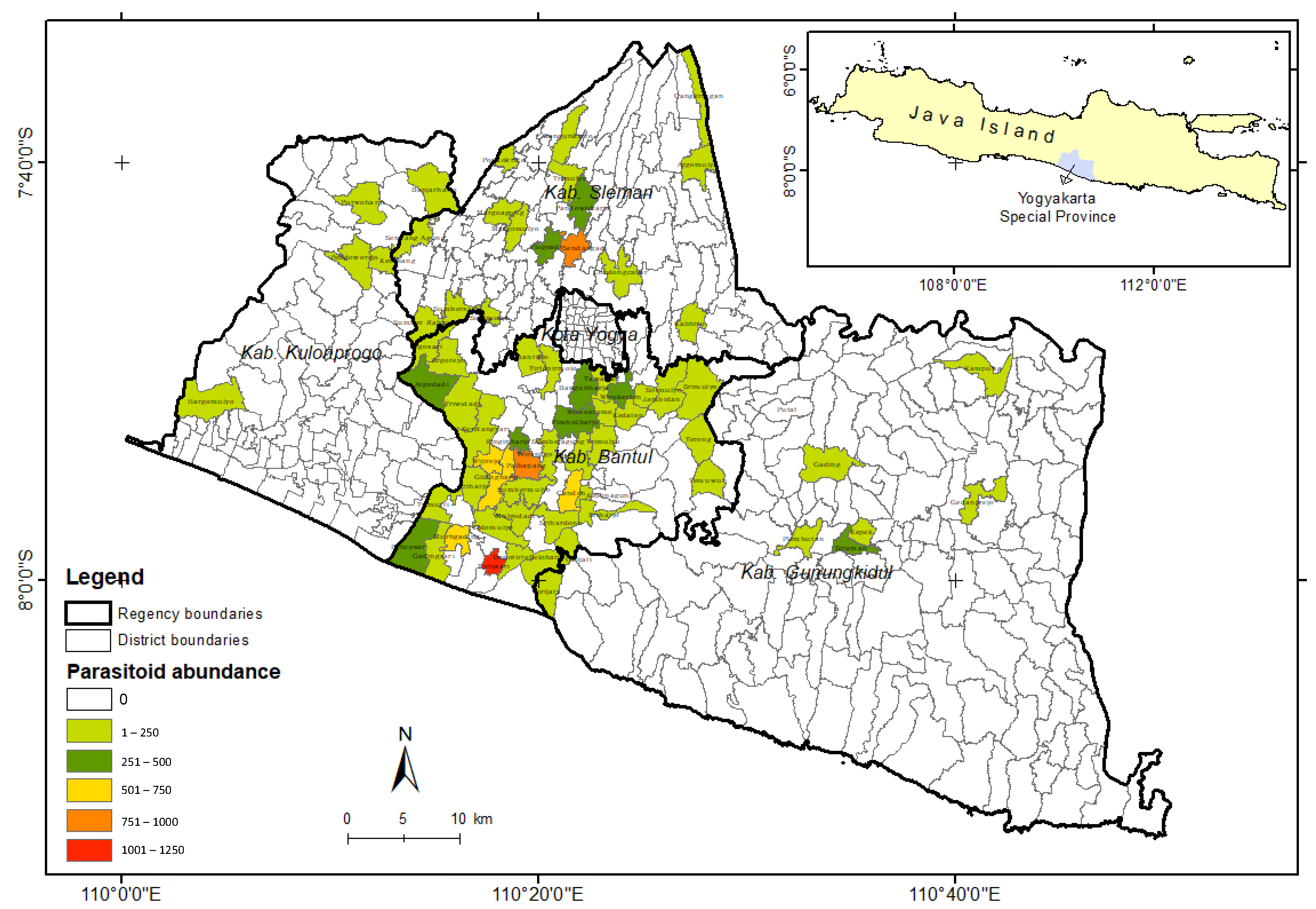
3. Results
4. Discussion

5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, M.P.; Clusella-Trullas, S.; Terblanche, J.S.; Richardson, D.M. Drivers, impacts, mechanisms and adaptation in insect invasions. Biol. Invasion 2016, 18, 883–891. [Google Scholar] [CrossRef]
- Rizali, A.; Oktaviyani, O.; Putri, S.; Doananda, M.; Linggani, A. Invasion of fall armyworm Spodoptera frugiperda, a new invasive pest, alters native herbivore attack intensity and natural enemy diversity. Biodivers. J. Biol. Divers. 2021, 22, 3482–3488. [Google Scholar] [CrossRef]
- Nonci, N.; Kalqutny, S.H.; Muis, A.; Azrai, M.; Aqil, M. Pengenalan Fall Armyworm (Spodoptera frugiperda JE Smith) Hama Baru pada Tanaman Jagung di Indonesia; Balai Penelitian Tanaman Serealia: Maros, Indonesia, 2019. (In Indonesian) [Google Scholar]
- Girsang, S.S.; Nurzannah, S.E.; Girsang, M.A.; Effendi, R. The distribution and impact of fall army worm (Spodoptera frugiperda) on maize production in North Sumatera. In Proceedings of the International Conference on Sustainable Cereals and Crops Production Systems in the Tropics, Makassar, Indonesia, 23–24 September 2020; p. 012099. [Google Scholar]
- Maharani, Y.; Dewi, V.K.; Puspasari, L.T.; Rizkie, L.; Hidayat, Y.; Dono, D. Cases of fall army worm Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) attack on maize in Bandung, Garut and Sumedang district, West Java. Cropsaver-J. Plant Prot. 2019, 2, 38–46. [Google Scholar] [CrossRef]
- Subiono, T. Preferences of Spodoptera frugiperda (Lepidoptera: Noctuidae) in several feed sources. J. Agroekoteknol. Trop. Lembab 2020, 2, 130–134. (In Indonesian) [Google Scholar] [CrossRef]
- Noer, H. Population and attack rate Spodoptera frugiperda on corn plants in Tulo Village, Sigi Regency. J. Agrotech 2020, 10, 66–68. (In Indonesian) [Google Scholar] [CrossRef]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2018, 65, 1–9. [Google Scholar] [CrossRef]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda JE Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Lubis, A.A.N.; Anwar, R.; Soekarno, B.P.; Istiaji, B.; Dewi, S.; Herawati, D. Coray wood corn (Spodoptera frugiperda) caterpillars in corn crops in Petir Village, Daramaga Sub-District, Bogor District, and its control potential using Metarizhium rileyi. J. Pus. Inov. Masy. 2020, 2, 931–939. (In Indonesian) [Google Scholar]
- Song, Y.; Yang, X.; Li, H.; Wu, K. The invasive Spodoptera frugiperda (JE Smith) has displaced Ostrinia furnacalis (Guenée) as the dominant maize pest in the border area of southwestern China. Pest Manag. Sci. 2023, 79, 3354–3363. [Google Scholar] [CrossRef]
- Kenis, M. Prospects for classical biological control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in invaded areas using parasitoids from the Americas. J. Econ. Entomol. 2023, 116, 331–341. [Google Scholar] [CrossRef]
- Tambo, J.A.; Kansiime, M.K.; Mugambi, I.; Rwomushana, I.; Kenis, M.; Day, R.K.; Lamontagne-Godwin, J. Understanding smallholders’ responses to fall armyworm (Spodoptera frugiperda) invasion: Evidence from five African countries. Sci. Total Environ. 2020, 740, 140015. [Google Scholar] [CrossRef]
- Hajek, A.E. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Cock, M. A Review of Biological Control of Pests in the Commonwealth Caribbean and Bermuda up to 1982; Commonwealth Institute of Biological Control Technical Communication, Farnham Royal: London, UK, 1985. [Google Scholar]
- Rosen, D.; Bennett, F.D.; Capinera, J.L. Pest Management in the Subtropics; Intercept: Andover, UK, 1994. [Google Scholar]
- Hokkanen, H.M.; Pimentel, D. New associations in biological control: Theory and practice. Can. Entomol. 1989, 121, 829–840. [Google Scholar] [CrossRef]
- Chandler, A.C.; Read, C. Introduction to Parasitology; John Willey and Sons: New York, NY, USA, 1961. [Google Scholar]
- Pimentel, D. Introducing parasites and predators to control native pests. Can. Entomol. 1963, 95, 785–792. [Google Scholar] [CrossRef]
- Colmenarez, Y.C.; Babendreier, D.; Ferrer Wurst, F.R.; Vásquez-Freytez, C.L.; de Freitas Bueno, A. The use of Telenomus remus (Nixon, 1937) (Hymenoptera: Scelionidae) in the management of Spodoptera spp.: Potential, challenges and major benefits. CABI Agric. Biosci. 2022, 3, 1–13. [Google Scholar] [CrossRef]
- Wengrat, A.P.; Coelho Junior, A.; Parra, J.R.; Takahashi, T.A.; Foerster, L.A.; Corrêa, A.S.; Polaszek, A.; Johnson, N.F.; Costa, V.A.; Zucchi, R.A. Integrative taxonomy and phylogeography of Telenomus remus (Scelionidae), with the first record of natural parasitism of Spodoptera spp. in Brazil. Sci. Rep. 2021, 11, 14110. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.-A.; Day, R.; Desneux, N.; Harrison, R.D.; Kriticos, D.; Rwomushana, I.; van den Berg, J. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2022, 43, 187–241. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall armyworm, Spodoptera frugiperda infestations in East Africa: Assessment of damage and parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef]
- Huo, L.; Zhou, J.; Ning, S.; Zhao, Q.; Zhang, L.; Zhang, Z.; Zhang, L.; Dong, H. Biological characteristics of Telenomus remus against Spodoptera frugiperda and Spodoptera litura eggs. Plant Prot. 2019, 45, 60–64. [Google Scholar]
- Tang, J.; Lyu, B.; Lu, H.; Ji, X.; Yang, P.; Su, H.; Cai, B. Investigation and preliminary study of biological characteristic of parasitic wasps of Spodoptera frugiperda in Hainan. Chin. J. Trop. Crops 2020, 41, 1189. [Google Scholar]
- Navik, O.; Shylesha, A.; Patil, J.; Venkatesan, T.; Lalitha, Y.; Ashika, T. Damage, distribution and natural enemies of invasive fall armyworm Spodoptera frugiperda (JE smith) under rainfed maize in Karnataka, India. Crop Prot. 2021, 143, 105536. [Google Scholar] [CrossRef]
- Trisyono, Y.A.; Suputa, S.; Aryuwandari, V.E.F.; Hartaman, M.; Jumari, J. Occurrence of heavy infestation by the fall armyworm Spodoptera frugiperda, a new alien invasive pest, in corn Lampung Indonesia. J. Perlindungan Tanam. Indones. 2019, 23, 156–160. [Google Scholar] [CrossRef]
- Megasari, D.; Putra, I.L.I.; Martina, N.D.; Wulanda, A.; Khotimah, K. Biology of Spodoptera frugiperda JE Smith on some types of feed in the laboratory. Agrovigor. J. Agroekoteknol. 2022, 15, 63–67. (In Indonesian) [Google Scholar] [CrossRef]
- Putra, I.L.I.; Martina, N.D. Life cycle of Spodoptera frugiperda with feeding kale and leeks in laboratory. J. Ilmu Pertan. Indones. 2021, 26, 386–391. (In Indonesian) [Google Scholar]
- Nurkomar, I.; Trisnawati, D.W.; Fahmi, F.; Buchori, D. Survival, development, and fecundity of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on various host plant species and their implication for the pest management. Insects 2023, 14, 629. [Google Scholar] [CrossRef] [PubMed]
- Sari, A.; Buchori, D.; Nurkomar, I. The potential of Telenomus remus Nixon (Hymenoptera: Scelinoidae) as biocontrol agent for the new fall armyworm S. frugiperda (Lepidoptera: Noctuidae) in Indonesia. Planta Trop. 2020, 8, 69–74. [Google Scholar] [CrossRef]
- Putra, I.L.I.; Wati, C.D.N.S. Parasicity level of Telenomus sp. parasitoid against Spodoptera frugiperda JE Smith eggs in the laboratory. J. Nat. Sci. Math. Res. 2020, 6, 73–77. [Google Scholar] [CrossRef]
- Minarni, E.W. Eksplorasi musuh alami hama Spodoptera frugiperda pada pertanaman jagung di lahan kelompok tani raden Kelurahan Pabuwaran Kecamatan Purwokerto Utara Kabupaten Banyumas (in Indonesia). In Proceedings of the Prosiding Seminar Nasional LPPM Unsoed, Purwokerto, Indonesia, 17–18 November 2023; pp. 139–147. [Google Scholar]
- Polem, E.S.N. Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae): Serangan dan Parasitoidnya di Nagan Raya, Aceh. Undergraduate Thesis, IPB University, Bogor, Indonesia, 2021. (In Indonesian). [Google Scholar]
- Rongkok, H.T.; Pasaru, F. Identification of paracitoids of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its paracity levels in farmer-owned corn plantation in District Sigi and in District Donggala. J. Agrotekbis 2021, 9, 972–978. (In Indonesian) [Google Scholar]
- Sari, W.; Nelly, N. Natural enemies of Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) on corn plants in West Sumatera. In Proceedings of the 2nd Agrifood System International Conference, Padang, Indonesia, 8–9 November 2023; p. 012045. [Google Scholar]
- Supeno, B.; Tarmizi, T.; Meidiwarman, M.; Haryanto, H. Keragaman parasitoid yang berasosiasi dengan telur hama baru Spodoptera frugiperda di Pulau Lombok. Pros. Saintek 2021, 3, 418–423. (In Indonesian) [Google Scholar]
- Suroto, A.; Soesanto, L.; Bahrudin, M. Attack rate and natural enemies of Spodoptera frugiperda JE Smith on corn plants in five districts in Banyumas regency. Proc. Ser. Phys. Form. Sci. 2021, 2, 44–49. (In Indonesian) [Google Scholar] [CrossRef]
- Tarigan, S.; Maryana, N.; Mubin, N. Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae): Attacks and their natural enemies on corn plantations in Munte Village, Munte Sub-district, Karo District, North Sumatera. In Proceedings of the International Conference on Modern and Sustainable Agriculture 2022, Padang, Indonesia, 24 November 2022; p. 012030. [Google Scholar]
- Tawakkal, M.; Buchori, D.; Maryana, N. New association between Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) and native natural enemies: Bioprospection of native natural enemies as biological control agents. In Proceedings of the 2nd International Symposium on Transdisciplinarity Approach for Knowledge Co-Creation in Sustainability—Understanding Complexity and Transdisciplinarity for Environmental Sustainability, Bogor, Indonesia, 6 November 2021; p. 012030. [Google Scholar]
- Nurkomar, I.; Putra, I.; Trisnawati, D.; Saman, M.; Pangestu, R.; Triyono, A. The existence and population dynamic of new fall armyworm species Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) in Yogyakarta, Indonesia. In Proceedings of the the 3rd International COnference on Sustainable Agriculture, Yogyakarta, Indonesia, 22–23 November 2021; p. 012023. [Google Scholar]
- Agboyi, L.K.; Mensah, S.A.; Clottey, V.A.; Beseh, P.; Glikpo, R.; Rwomushana, I.; Day, R.; Kenis, M. Evidence of leaf consumption rate decrease in fall armyworm, Spodoptera frugiperda, larvae parasitized by Coccygidium luteum. Insects 2019, 10, 410. [Google Scholar] [CrossRef]
- Shylesha, A.; Jalali, S.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K. Studies on new invasive pest Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) and its natural enemies. J. Biol. Control 2018, 32, 1–7. [Google Scholar] [CrossRef]
- Wei, Y.-W.; Zhou, Y.-B.; Zou, Q.-C.; Sheng, M.-L. A new species of Campoletis Förster (Hymenoptera, Ichneumonidae) with a key to species known from China, Japan and South Korea. ZooKeys 2020, 1004, 99–108. [Google Scholar] [CrossRef] [PubMed]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- DinasPerindustrianPerdagangandanKoperasiKabupatenBantul. Prospektus Investasi Sektor Pangan Kabupaten Bantul (in Indonesian). Available online: https://dpmptsp.bantulkab.go.id/web/potensi_investasi/detail/19-sektor-pangan (accessed on 12 September 2023).
- Sunari, A.A.A.A.S.; Sumiartha, I.K.; Supartha, I.W.; Mahaputra, I.F.; Yudha, I.K.W.; Utama, I.W.E.K.; Wiradana, P.A. Potential yield reduction of sweet and glutinous corn varieties damaged by the invasive pest Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae): Field trial scale. In Proceedings of the 2nd International Conference on Education and Technology (ICETECH 2021), Madiun, Indonesia, 26–27 November 2022; pp. 14–19. [Google Scholar]
- Kishinevsky, M.; Keasar, T.; Bar-Massada, A. Parasitoid abundance on plants: Effects of host abundance, plant species, and plant flowering state. Arthropod-Plant Interact. 2017, 11, 155–161. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Elton, C.S. 1958: The ecology of invasions by animals and plants. London: Methuen. Prog. Phys. Geogr. 2007, 31, 659–666. [Google Scholar] [CrossRef]
- Jindal, J.; Sharma, K.P.; Shera, P.S.; Cheema, H.K. Native Parasitoids of Fall Army Worm Spodoptera frugiperda (J E Smith) in Maize. Indian J. Entomol. 2022, 84, 865–867. [Google Scholar] [CrossRef]
- Pu’u, Y.M.; Mutiara, C. Attacks of invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera:Noctuidae) on corn plantation in Ende District Flores, Indonesia. J. Entomol. Indones. 2021, 18, 153. (In Indonesian) [Google Scholar] [CrossRef]
- Salazar-Mendoza, P.; Peralta-Aragón, I.; Romero-Rivas, L.; Salamanca, J.; Rodriguez-Saona, C. The abundance and diversity of fruit flies and their parasitoids change with elevation in guava orchards in a tropical Andean forest of Peru, independent of seasonality. PLoS ONE 2021, 16, e0250731. [Google Scholar] [CrossRef] [PubMed]
- Syahidah, T.; Prasetyo, L.; Buchori, D. Landscape structure and parasitoid diversity as measures of sustainable landscape. IOP Conf. Ser. Earth Environ. Sci. 2019, 325, 012012. [Google Scholar] [CrossRef]
- Schoeninger, K.; Souza, J.L.; Krug, C.; Oliveira, M.L. Diversity of parasitoid wasps in conventional and organic guarana (Paullinia cupana var. sorbilis) cultivation areas in the Brazilian Amazon. Acta Amaz. 2019, 49, 283–293. [Google Scholar] [CrossRef]
- Abang, A.; Nanga, S.; Fotso Kuate, A.; Kouebou, C.; Suh, C.; Masso, C.; Saethre, M.; Fiaboe, K. Natural Enemies of Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Agro-Ecologies. Insect 2021, 12, 509. [Google Scholar] [CrossRef] [PubMed]
- Sharanabasappa, S.; Kalleshwaraswamy, C.; Poorani, J.; Maruthi, M.; Pavithra, H.; Diraviam, J. Natural enemies of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), a recent invasive pest on maize in South India. Fla. Entomol. 2019, 102, 619–623. [Google Scholar] [CrossRef]
- Kenis, M.; Du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef]
- Liao, Y.-L.; Yang, B.; Xu, M.-F.; Lin, W.; Wang, D.-S.; Chen, K.-W.; Chen, H.-Y. First report of Telenomus remus parasitizing Spodoptera frugiperda and its field parasitism in southern China. J. Hymenopt. Res. 2019, 73, 95–102. [Google Scholar] [CrossRef]
- Sari, A.; Nurkomar, I.; Buchori, D. Comparison of Spodoptera frugiperda parasitoid performance under laboratory conditions. In Proceedings of the 3rd International Symposium on Transdisciplinary Approach for Knowledge Co-Creation in Sustainability, Bogor, Indonesia, 24 November 2023; p. 012031. [Google Scholar]
- Junaedi, E.; Yunus, M.; Hasriyanty, H. Parasitoids and it is parasitisme on white rice stem borer (Scirpophaga innotata Walker) in two different altitudes of rice fields (Oryza sativa l.) in District of Sigi. e-J. Agrotekbis 2016, 4, 280–287. (In Indonesian) [Google Scholar]
- Legault, S.; Hébert, C.; Blais, J.; Berthiaume, R.; Bauce, E.; Brodeur, J. Seasonal ecology and thermal constraints of Telenomus spp.(Hymenoptera: Scelionidae), egg parasitoids of the hemlock looper (Lepidoptera: Geometridae). Environ. Entomol. 2012, 41, 1290–1301. [Google Scholar] [CrossRef]
- Goulart, M.M.P.; Bueno, A.d.F.; Bueno, R.C.O.d.F.; Diniz, A.F. Host preference of the egg parasitoids Telenomus remus and Trichogramma pretiosum in laboratory. Rev. Bras. Entomol. 2011, 55, 129–133. [Google Scholar] [CrossRef]
- Hernandez, D.; Ferrer, F.; Linares, B. Introducción de Telenomus remus Nixon (Hym.: Scelionidae) para controlar Spodoptera frugiperda (Lep.: Noctuidae) en Yaritagua-Venezuela. Agron. Trop. 1989, 39, 45–61. [Google Scholar]
- Cave, R.D. Biology, ecology and use in pest management of Telenomus remus. Biocontrol News Inf. 2000, 21, 21N–26N. [Google Scholar]
- Copeland, R. Fall Armyworm Natural Enemy of the Day: Telenomus remus (Hymenoptera: Scelionidae). Available online: http://www.icipe.org/content/fall-armyworm-natural-enemy-day-telenomus-remus (accessed on 12 September 2023).
- Polaszek, A.; Kimani, S.W. Telenomus species (Hymenopetra: Scelionidae) attacking eggs of pyralid pests (Lepidoptera) in Africa: A review and guide to identification. Bull. Entomol. Res. 1990, 80, 57–71. [Google Scholar] [CrossRef]
- Kalyanasundaram, M.; Kamala, I.M. Parasitoids. In Ecofriendly Pest Management for Food Security; Elsevier: Amsterdam, The Netherlands, 2016; pp. 109–138. [Google Scholar]
- Buhl, P.N. Species of Platygastrinae and Sceliotrachelinae from rainforest canopies in Tanzania, with keys to the Afrotropical species of Amblyaspis, Inostemma, Leptacis, Platygaster and Synopeas (Hymenoptera, Platygastridae). Tijdschr. Voor Entomol. 2011, 154, 75–126. [Google Scholar] [CrossRef]
- Buhl, P.N. Key to Platygaster (Hymenoptera: Platygastridae) from Denmark, with descriptions of new species. Steenstrupia 2006, 29, 127–167. [Google Scholar]
- McCutcheon, G.S.; Salley, W.; Turnipseed, S. Biology of Apanteles ruficrus, an imported parasitoid of Pseudoplusia includens, Trichoplusia ni, and Spodoptera frugiperda (Lepidoptera: Noctuidae). Environ. Entomol. 1983, 12, 1055–1058. [Google Scholar] [CrossRef]
- Rajapakse, R.H.; Ashley, T.R.; Waddill, V.H. Biology and host acceptance of Microplitis manilae (Hymenoptera: Braconidae) raised on fall armyworm larvae Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 1985, 68, 653–657. [Google Scholar] [CrossRef]
- Alam, M. Attempts at the biological control of major insect pests of maize in Barbados, WI. Proceeding Caribb. Food Crops Soc. 1978, 15, 127–135. [Google Scholar]
- Whitfield, J.B.; Austin, A.D.; Fernandez-Triana, J.L. Systematics, biology, and evolution of microgastrine parasitoid wasps. Annu. Rev. Entomol. 2018, 63, 389–406. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Meagher, R.L. The Spodoptera frugiperda host strains: What they are and why they matter for understanding and controlling this global agricultural pest. J. Econ. Entomol. 2022, 115, 1729–1743. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.R. Little-known fly-promising biocontrol weapon. Agric. Res. 1985, 33, 12. [Google Scholar]
- Deshmukh, S.S.; Kiran, S.; Naskar, A.; Pradeep, P.; Kalleshwaraswamy, C.; Sharath, K. First record of a parasitoid, Megaselia (M) scalaris (Diptera: Phoridae) of fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) from India. Egypt. J. Biol. Pest Control 2021, 31, 94. [Google Scholar] [CrossRef]
- Tang, Y.; Li, Q.; Xiang, L.; Gu, R.; Wu, Y.; Zhang, Y.; Bai, X.; Niu, X.; Li, T.; Wei, J. First report on Megaselia scalaris Loew (Diptera: Phoridae) infestation of the invasive pest Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in China. Insects 2021, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Ruíz-Nájera, R.E.; Molina-Ochoa, J.; Carpenter, J.E.; Espinosa-Moreno, J.A.; Ruíz-Nájera, J.A.; Lezama-Gutierrez, R.; Foster, J.E. Survey for hymenopteran and dipteran parasitoids of the fall armyworm (Lepidoptera: Noctuidae) in Chiapas, Mexico. J. Agric. Urban Entomol. 2007, 24, 35–42. [Google Scholar] [CrossRef]
- Disney, R.H.L. Natural history of the scuttle fly, Megaselia scalaris. Annu. Rev. Entomol. 2008, 53, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Daif, M.K.; Mosallam, A.; Ebrahim, A. The scuttle fly, Megaselia scalaris (Loew, 1866)(Diptera: Phoridae): A new threat on laboratory aass production of fruit flies. J. Plant Prot. Pathol. 2023, 14, 109–113. [Google Scholar] [CrossRef]
- Rajan, R.; Shamsudeen, R. First record of Megaselia scalaris (Diptera: Phoridae) as a parasitoid of Thyas coronata (Lepidoptera: Erebidae). Indian J. Entomol. 2023, 85, 4. [Google Scholar] [CrossRef]
- Acevedo-Alcalá, A.; Lomeli-Flores, J.R.; Rodríguez-Leyva, E.; Rodríguez-Rodríguez, S.E.; Ortiz-Andrade, E. Does Megaselia scalaris (Diptera: Phoridae) have potential as a biological control agent of fall armyworm? Fla. Entomol. 2023, 106, 56–58. [Google Scholar] [CrossRef]
- Wibowo, L. The abundance and diversity of the parasitoid of banana leaf skipper pest (Erionota thrax L.) in South Lampung Regency. J. Hama Dan Penyakit Tumbuh. Trop. 2015, 15, 26–32. (In Indonesian) [Google Scholar] [CrossRef]
- Fawzi, F.S.; Iman, I.a.-S.; Adil, A.H.; Muhammad, A.a.-S. Survey and distribution density of genus Brachymeria species (Hymenoptera: Chalcididae) in Egypt. Egypt. J. Plant Prot. Res. Inst. 2020, 3, 415–432. [Google Scholar]
- Buchori, D.; Herawati, E.; Sari, A. The effectiveness of Telenomus remus (Nixon)(Hymenoptera: Scelionidae) for controlling welsh onion pest Spodoptera exigua Hübner (Lepidoptera: Noctuidae). J. Entomol. Indones. 2008, 5, 81–95. (In Indonesian) [Google Scholar] [CrossRef]
- Ratna, E.S. Efficiency of parasitization on larval host, Spodoptera litura (F.), by an endoparasitoid Snellenius (=Microplitis) manilae Ashmead in the laboratory. J. Hama Dan Penyakit Tumbuh. Trop. 2008, 8, 8–16. (In Indonesian) [Google Scholar] [CrossRef]
- Jin, T.; Lin, Y.; Ma, G.; Liu, J.; Hao, Z.; Han, S.; Peng, Z. Biocontrol potential of Trichogramma species against Spodoptera frugiperda and their field efficacy in maize. Crop Prot. 2021, 150, 105790. [Google Scholar] [CrossRef]
- Abd-Rabou, S. The efficacy of indigenous parasitoids in the biological control of Siphoninus phillyreae (Homoptera: Aleyrodidae) on pomegranate in Egypt. Pan-Pac. Entomol. 1998, 74, 169–173. [Google Scholar]
- Abd-Rabou, S.; Simmons, A.M. Augmentation and evaluation of a parasitoid, Encarsia inaron, and a predator, Clitostethus arcuatus, for biological control of the pomegranate whitefly, Siphoninus phillyreae. Arch. Phytopathol. Plant Prot. 2010, 43, 1318–1334. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| District | Number of S. frugiperda (Mean ± SE) | Parasitism Rate (%) (Mean ± SE) | n |
|---|---|---|---|
| Egg | |||
| Bantul | 1918.00 ± 465.18 b | 28.10 ± 6.81 b | 17 |
| Gunung Kidul | 997.14 ± 376.88 ab | 10.38 ± 3.92 a | 7 |
| Kulonprogo | 146.00 ± 65.29 a | 71.97 ± 32.18 ab | 5 |
| Sleman | 190.09 ± 57.31 a | 34.03 ± 10.26 ab | 11 |
| Larva | |||
| Bantul | 43.20 ± 21.60 a | 5.34 ± 2.67 a | 4 |
| Gunung Kidul | 40.50 ± 20.25 a | 18.39 ± 9.20 a | 4 |
| Kulonprogo | 14.00 ± 9.90 a | 11.45 ± 8.10 a | 2 |
| Sleman | 48.50 ± 19.80 a | 10.55 ± 4.31 a | 6 |
| Order | Family | Species | Parasitism Rate (Mean ± SE, %) | |||
|---|---|---|---|---|---|---|
| Bantul | Gunung Kidul | Kulon Progo | Sleman | |||
| Egg parasitoid | ||||||
| Hymenoptera | Platygasteridae | Platygasteridae.sp01 * | 1.46 ± 1.01 | |||
| Platygasteridae | Platygasteridae.sp02 * | 42.00 ± 0.00 | ||||
| Platygasteridae | Telenomus remus * | 37.29 ± 4.16 | 14.74 ± 5.58 | 71.97 ± 15.62 | 33.23 ± 8.33 | |
| Trichogrammatidae | Trichogramma sp. | 5.78 ± 4.36 | ||||
| Larval parasitoid | ||||||
| Hymenoptera | Braconidae | Cotesia sp. * | 11.89 ± 7.30 | 16.67 ± 0.00 | 5.02 ± 0.79 | |
| Ichneumonidae | Campoletis sp. * | 1.61 ± 0.00 | 2.78 ± 0.00 | |||
| Braconidae | Coccygidium sp. * | 5.56 ± 0.00 | 2.78 ± 0.00 | |||
| Eupelmidae | Eupelmidae.sp01 * | 6.25 ± 0.00 | ||||
| Braconidae | Microplitis sp. * | 39.70 ± 26.92 | 61.29 ± 0.00 | |||
| Braconidae | Stenobracon sp.* | 11.54 ± 0.00 | ||||
| Diptera | Tachinidae | Archytas marmoratus | 4.03 ± 0.22 | |||
| Phoridae | Megaselia scalaris | 3.70 ± 0.00 | ||||
| Larval–pupal parasitoid | ||||||
| Hymenoptera | Chalcididae | Brachymeria femorata * | 6.25 ± 0.00 | |||
| Chalcididae | Brachymeria lasus * | 6.82 ± 0.00 | ||||
| Ichneumonidae | Charops sp. * | 1.04 ± 0.00 | 4.02 ± 0.87 | |||
| Order | Family | Species | Abundance | |||
|---|---|---|---|---|---|---|
| Bantul | Gunung Kidul | Kulon Progo | Sleman | |||
| Egg parasitoid | ||||||
| Hymenoptera | Platygasteridae | Platygasteridae.sp01 | 49 | |||
| Platygasteridae | Platygasteridae.sp02 | 21 | ||||
| Platygasteridae | Telenomus remus | 8536 | 831 | 466 | 2324 | |
| Trichogrammatidae | Trichogramma sp. | 1988 | ||||
| Larval parasitoid | ||||||
| Hymenoptera | Braconidae | Cotesia sp. | 27 | 12 | 110 | |
| Ichneumonidae | Campoletis sp. | 1 | 1 | |||
| Braconidae | Coccygidium sp. | 1 | 1 | |||
| Eupelmidae | Eupelmidae.sp01 | 1 | ||||
| Braconidae | Microplitis sp. | 15 | 19 | |||
| Braconidae | Stenobracon sp. | 3 | ||||
| Diptera | Tachinidae | Archytas marmoratus | 2 | |||
| Phoridae | Megaselia scalaris | 5 | ||||
| Larval–pupal parasitoid | ||||||
| Hymenoptera | Chalcididae | Brachymeria femorata | 5 | |||
| Chalcididae | Brachymeria lasusi | 3 | ||||
| Ichneumonidae | Charops sp. | 1 | 2 | |||
| Species richness | 8 | 6 | 3 | 7 | ||
| Total abundance | 8753 | 924 | 479 | 2478 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nurkomar, I.; Putra, I.L.I.; Buchori, D.; Setiawan, F. Association of a Global Invasive Pest Spodoptera frugiperda (Lepidoptera: Noctuidae) with Local Parasitoids: Prospects for a New Approach in Selecting Biological Control Agents. Insects 2024, 15, 205. https://doi.org/10.3390/insects15030205
Nurkomar I, Putra ILI, Buchori D, Setiawan F. Association of a Global Invasive Pest Spodoptera frugiperda (Lepidoptera: Noctuidae) with Local Parasitoids: Prospects for a New Approach in Selecting Biological Control Agents. Insects. 2024; 15(3):205. https://doi.org/10.3390/insects15030205
Chicago/Turabian StyleNurkomar, Ihsan, Ichsan Luqmana Indra Putra, Damayanti Buchori, and Fajar Setiawan. 2024. "Association of a Global Invasive Pest Spodoptera frugiperda (Lepidoptera: Noctuidae) with Local Parasitoids: Prospects for a New Approach in Selecting Biological Control Agents" Insects 15, no. 3: 205. https://doi.org/10.3390/insects15030205
APA StyleNurkomar, I., Putra, I. L. I., Buchori, D., & Setiawan, F. (2024). Association of a Global Invasive Pest Spodoptera frugiperda (Lepidoptera: Noctuidae) with Local Parasitoids: Prospects for a New Approach in Selecting Biological Control Agents. Insects, 15(3), 205. https://doi.org/10.3390/insects15030205