
A Novel Interaction of Nesidiocoris tenuis (Hemiptera: Miridae) as a Biological Control Agent of Bactericera cockerelli (Hemiptera: Triozidae) in Potato

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Potato Psyllid Colony
2.2. Tomato Bug Colony
2.3. Tomato Bug Adults for Trials
2.4. Assessing the Model or Functional Response of N. tenuis Preying on B. cockerelli on Potato under Laboratory and Greenhouse Conditions
2.5. Efficacy of N. tenuis on B. cockerelli in Potato in Field Cage Studies
2.6. Predation Efficacy of N. tenuis on B. cockerelli on Potato under Field Conditions, Coupled with a Reduced Insecticide Program, and the Impact on Potato Yield and Incidence and Severity of Zebra Chip Disease
2.6.1. Controlled of N. tenuis Releases
2.6.2. Scheduled-Program Controlled Releases of N. tenuis
2.6.3. Scheduled Program for the RIP
2.6.4. Insecticide Applications and Organic Management
2.6.5. Population Dynamics of B. cockerelli and Predator
2.6.6. Impact on Potato Yield and Incidence and Severity of Zebra Chip Disease
2.7. Data Analysis
3. Results
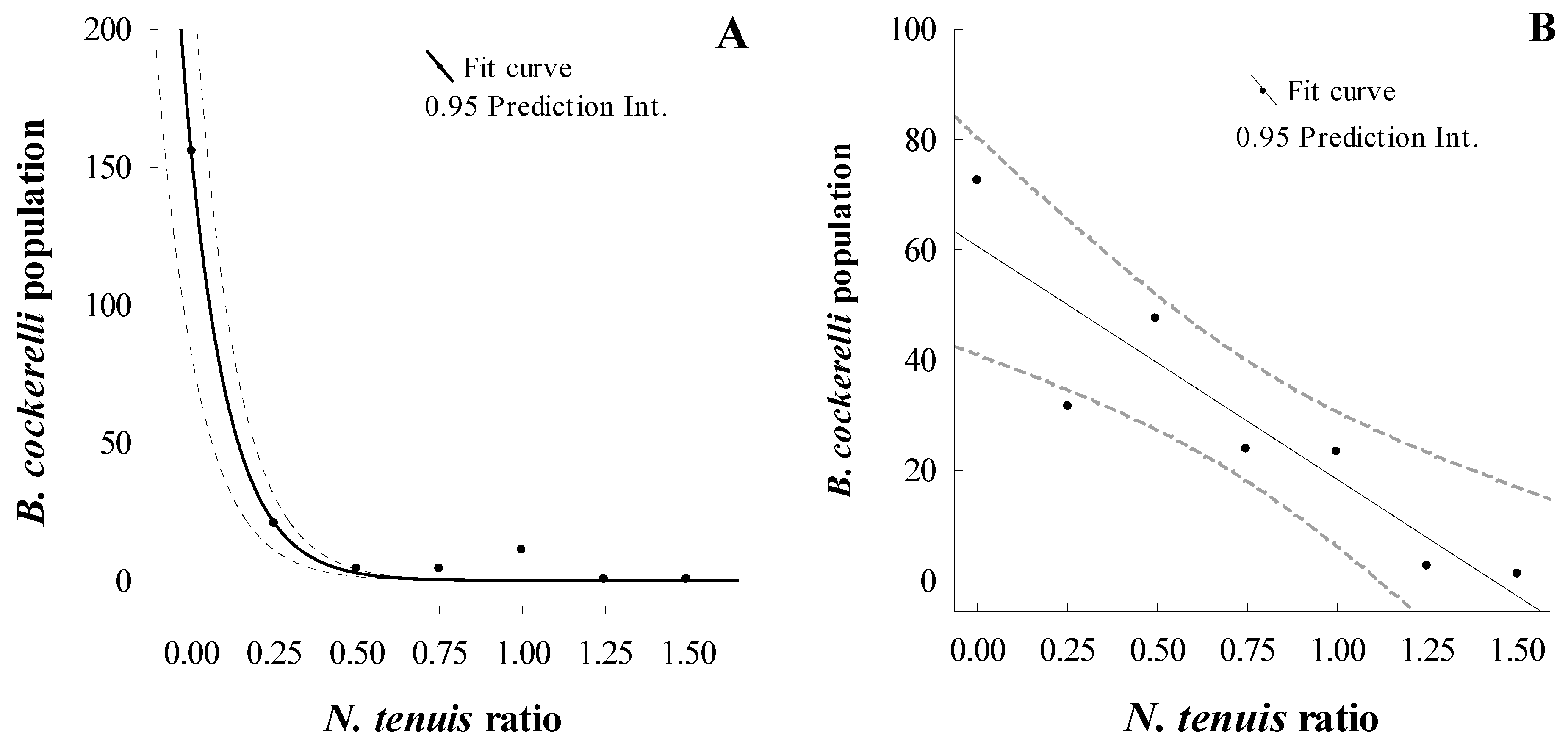
3.1. Assessing the Mathematical Model and Functional Response of N. tenuis Preying on B. cockerelli on Potato under Laboratory and Greenhouse Conditions
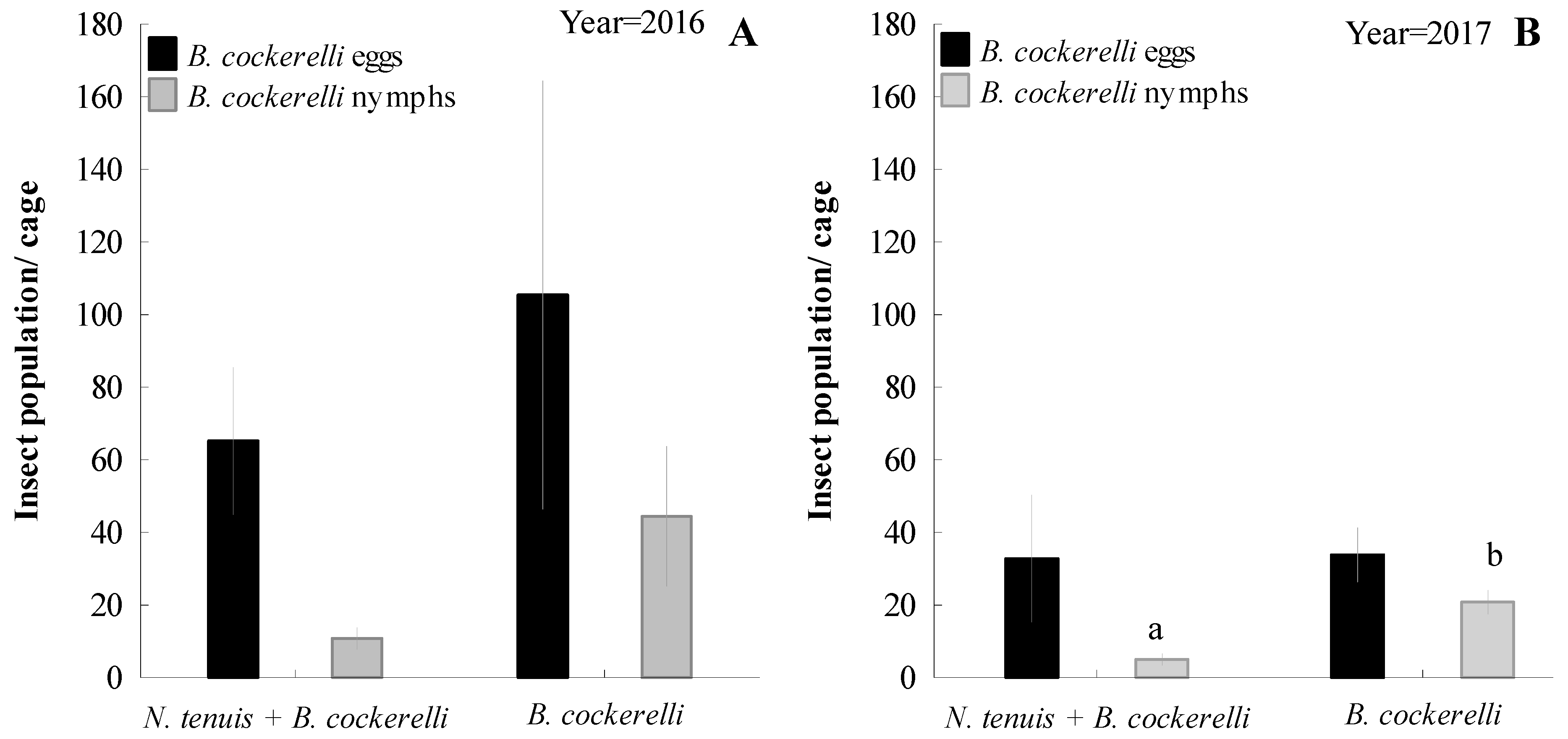
3.2. Efficacy of N. tenuis on B. cockerelli in Potato in Field Cage Studies
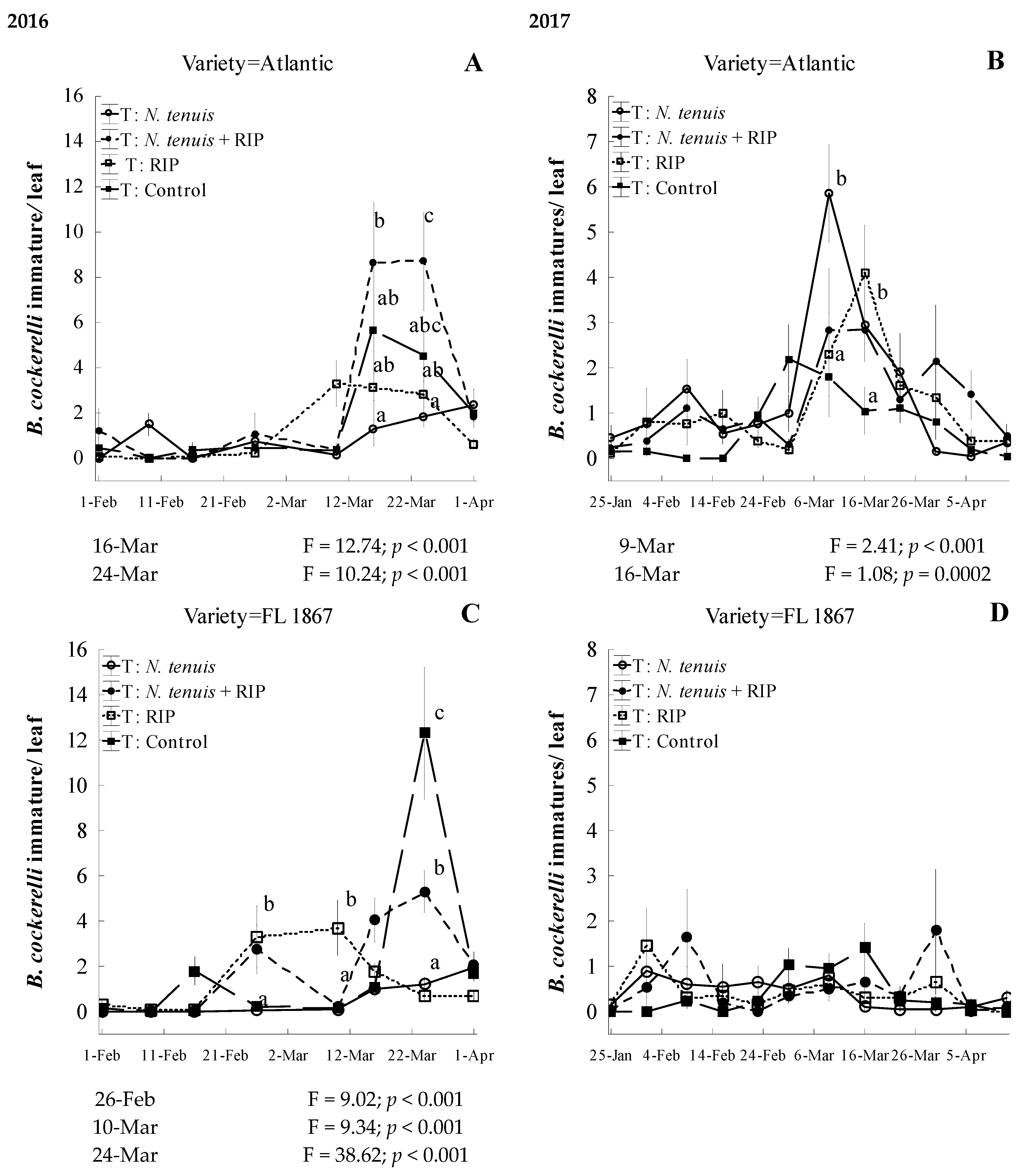
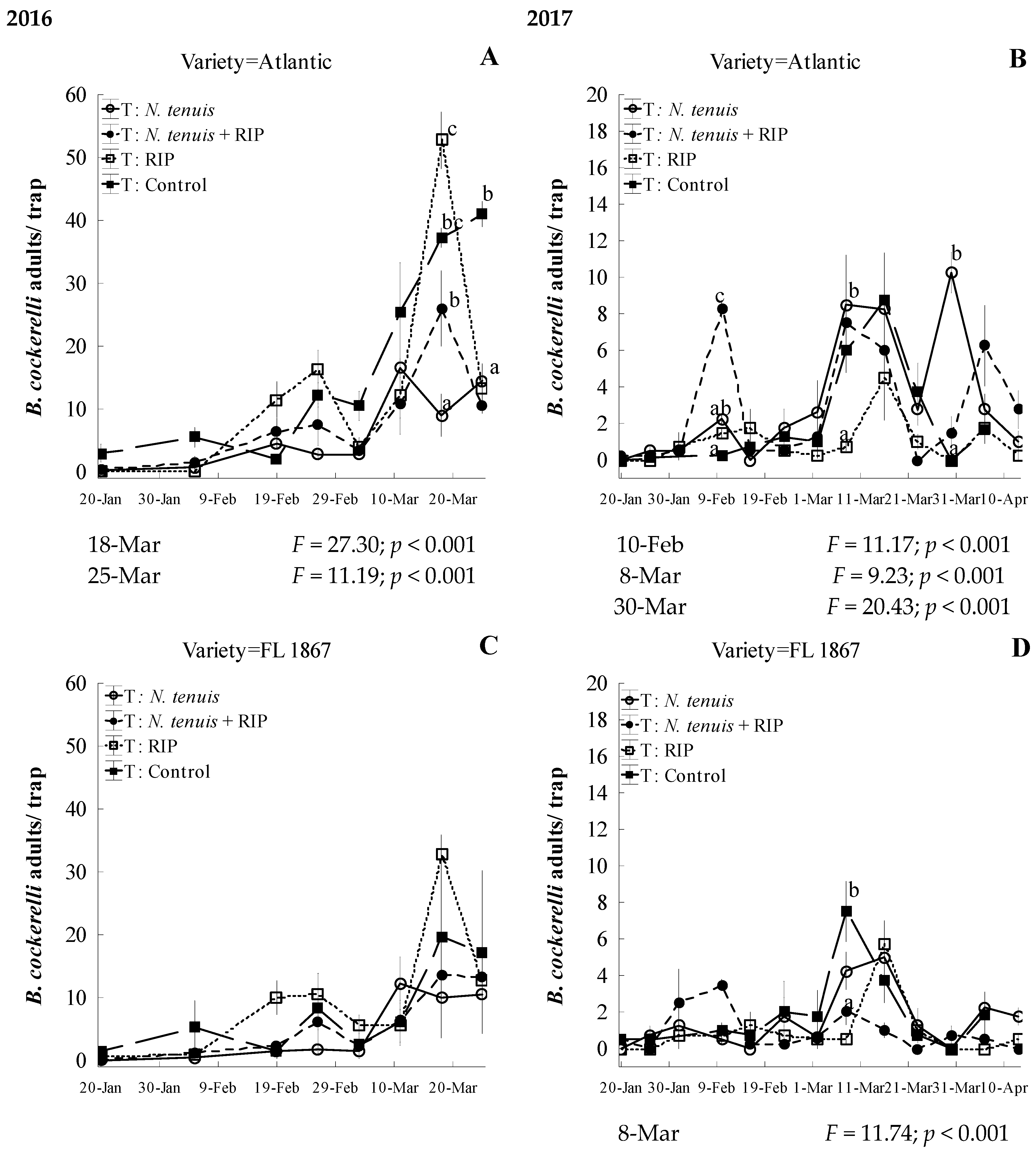
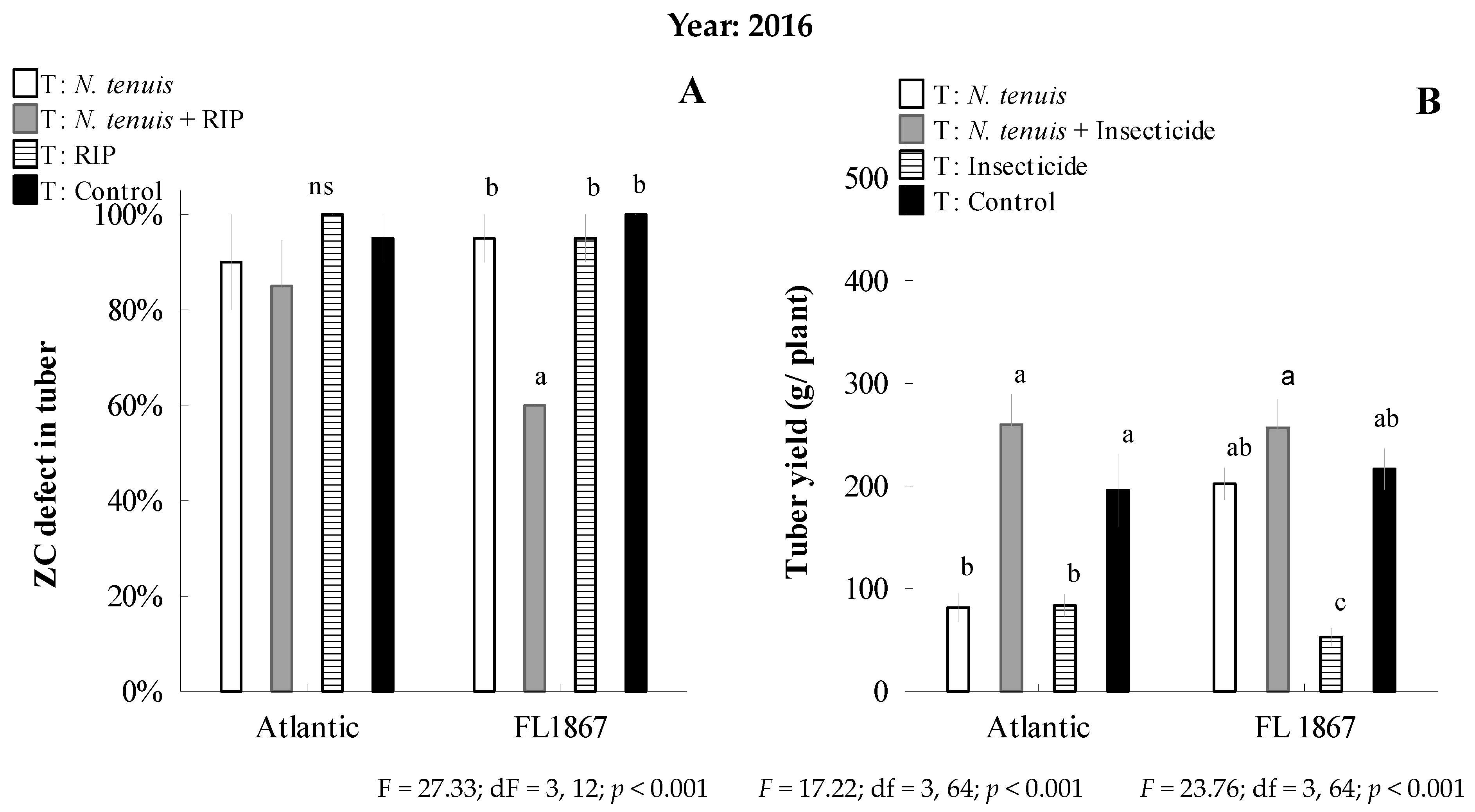
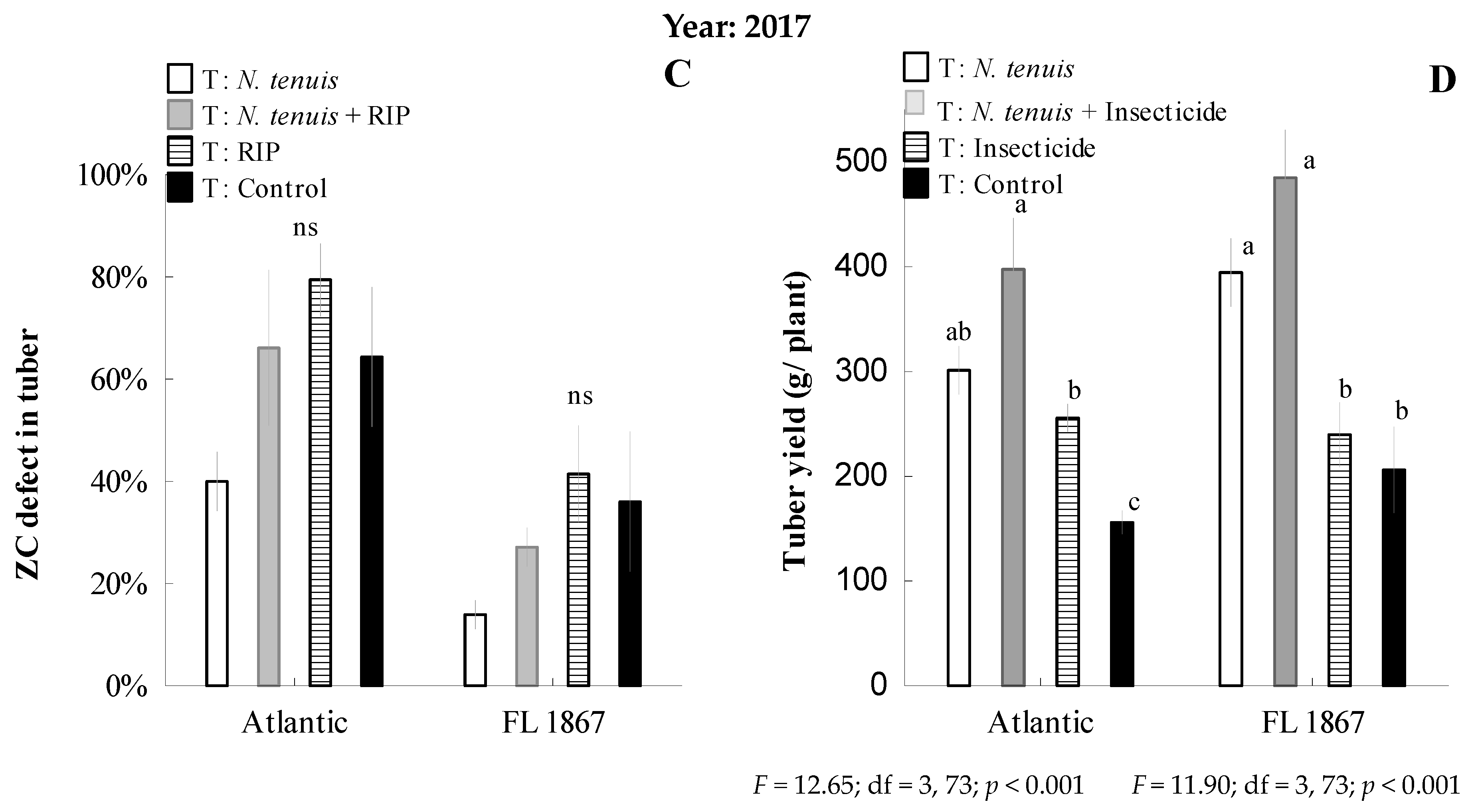
3.3. Predation Efficacy of N. tenuis on B. cockerelli on Potato under Field Conditions, Coupled with a Reduced Insecticide Program, and the Impact on Potato Yield and Incidence and Severity of Zebra Chip Disease
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- China, W.E. LXIX.—Synonymy of Engytatus tenuis Reut.(tobacco Capsid). J. Nat. Hist. 1938, 1, 604–607. [Google Scholar] [CrossRef]
- United Kingdom CAB International Distribution Maps of Plant Pests Map 290. Available online: https://www.cabi.org/ISC/abstract/20056600290 (accessed on 1 January 2016).
- Waterhouse, D.F. The Major Arthropod Pests and Weeds of Agriculture in Southeast Asia: Distribution, Importance and Origin. ACIAR Monogr. 1993, 21, 141. [Google Scholar]
- Duarte, L.; Nacional, C.; Agropecuaria, D.S.; Nacional, C.; Agropecuaria, D.S.; Rivas, A. Míridos depredadores (Hemiptera: Heteroptera: Miridae) en cultivos de tomate y tabaco en Cuba. Rev. Protección Veg. 2014, 29, 204–207. [Google Scholar]
- Kerzhner, I.M.; Josifov, M. Cimicomorpha II. Miridae. In Catalogue of the HeteroPtera of the Palaearctic Region; Aukema, B., Rieger, C., Eds.; Netherlands Entomological Society: Wageningen, The Netherlands, 1999; Volume 3, pp. 1–577. [Google Scholar]
- Biondi, A.; Zappalà, L.; Di Mauro, A.; Tropea Garzia, G.; Russo, A.; Desneux, N.; Siscaro, G. Can alternative host plant and prey affect phytophagy and biological control by the zoophytophagous mirid Nesidicoris tenuis? BioControl 2016, 61, 79–90. [Google Scholar] [CrossRef]
- Sohrabi, F.; Hosseini, R. Nesidicoris tenuis (Reuter) (Heteroptera: Miridae), a predatory species of the tomato leafminer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). J. Plant Prot. Res. 2015, 55, 3–4. [Google Scholar] [CrossRef]
- El Arnaouty, S.A.; Kortam, M.N. First record of the mired predatory species, Nesidicoris tenuis Reuter (Heteroptera: Miridae) on the tomato leafminer, Tuta absoluta. Egypt. J. Biol. Pest Control 2012, 22, 223–224. [Google Scholar]
- Sridhar, V.; Jayashankar, M.; Vinesh, L.S. Population dynamics of zoophytophagous mirid bug, Nesidicoris tenuis (Reuter) (Heteroptera: Miridae) and its prey, Bemisia tabaci Genn. (Homoptera: Aleyrodidae) on tomato (Solanum lycopersicum Mill). Pest Manag. Hortic. Ecosyst. 2012, 18, 35–38. [Google Scholar]
- Malausa, J.C.; Ehanno, B. First observations in France of Cyrtopeltis (Nesidiocoris)tenuis Reuter, 1895 (Het. Miridae). Nouv. Rev. D’entomologie 1988, 5, 180. [Google Scholar]
- Esparza-Díaz, G.; Villanueva, R.T. Actividad fitófaga de Nesidicoris tenuis (Hemiptera: Miridae) en cultivos de jitomate en el sur de Texas. In XXXVII Congreso Nacional de Control Biológico. Mérida; Lozano-Contreras, M.G., Burgos Tan, M.J., Arroyo López, J.I., Eds.; Mérida, Mexico, 2014; p. 768. Available online: http://sistemanodalsinaloa.gob.mx/archivoscomprobatorios/_15_memoriaextenso/12560.pdf (accessed on 1 January 2016).
- Esparza-Díaz, G.; Marconi, T.; Avila, C.A.; Villanueva, R.T. Persistence of the Exotic Mirid Nesidicoris tenuis in South Texas. Insects 2021, 12, 715. [Google Scholar] [CrossRef] [PubMed]
- Pineda, S.; Henry, T.J.; Corrales-Madrid, J.L.; Martínez, A.M.; Figueroa, J.I. First records of the dicyphine plant bug Nesidicoris tenuis (Hemiptera: Heteroptera: Miridae: Bryocorinae) in Mexico. Proc. Entomol. Soc. Wash. 2017, 119, 290–295. [Google Scholar] [CrossRef]
- Hoberlandt, L. Results of the Zoological Scientific Expedition of the National Museum in Prague to Turkey. 18. Hemiptera IV. Terrestrial Hemiptera-Heteroptera of Turkey. Acta Entomol. Musei Natl. Pragae 1956, 3, 1–264. [Google Scholar]
- Schuh, R. On-line Systematic Catalog of Plant Bugs (Insecta: Heteroptera: Miridae). Available online: http://research.amnh.org/pbi/catalog (accessed on 1 January 2016).
- Calvo, J.; Bolckmans, K.; Stansly, P.A.; Urbaneja, A. Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato. BioControl 2009, 54, 237–246. [Google Scholar] [CrossRef]
- Urbaneja, A.; Montón, H.; Mollá, O. Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J. Appl. Entomol. 2009, 133, 292–296. [Google Scholar] [CrossRef]
- Sanchez, J.A.; La-Spina, M.; Lacasa, A. Numerical response of Nesidiocoris tenuis (Hemiptera: Miridae) preying on Tuta absoluta (Lepidoptera: Gelechiidae) in tomato crops. Eur. J. Entomol. 2014, 111, 387–395. [Google Scholar] [CrossRef]
- Calvo, F.J.; Lorente, M.J.; Stansly, P.A.; Belda, J.E. Preplant release of Nesidicoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisia tabaci in greenhouse tomato. Entomol. Exp. Appl. 2012, 143, 111–119. [Google Scholar] [CrossRef]
- Romney, V.E. Breeding Areas of the Tomato Psyllid, Paratrioza cockerelli (Sulc). J. Econ. Entomol. 1939, 32, 150–151. [Google Scholar]
- Pletsch, D.J. The potato psyllid, Paratrioza cockerelli (Sulc), its biology and control. Bull. Mont. Agric. Exp. Stn. 1947, 446, 95. [Google Scholar]
- Wallis, R.L. Ecological studies on the potato psyllid as a pest of potatoes. USDA Tech. Bull. 1995, 1107, 25. [Google Scholar]
- Papp, M.R. Index to the Bulletin of the California Department of Agriculture; Sierra Graphics Typography—Sierra Pacific Press: Sacramento, CA, USA, 1965; Volume 2. [Google Scholar]
- Munyaneza, J.E.; Henne, D.C. Leafhopper and psyllid pests of potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Academic Press: San Diego, CA, USA, 2012; pp. 65–102. [Google Scholar]
- List, M.G. Potato and Tomato Psyllid and Its Control on Tomatoes. Color. Exp. Station. Color. State Coll. Bull. 1939, 454, 33. [Google Scholar]
- Pletsch, D.J. The Effect of Some Insecticides on the Immature Stages of the Potato and Tomato Psyllid, Paratrioza cockerelli (Sulc). J. Econ. Entomol. 1942, 35, 58–60. [Google Scholar] [CrossRef]
- Tuthill, L.D. Contributions to the Knowledge of the Psyllidae of Mexico (continued). Kans. Entomol. Soc. 1945, 18, 1–29. [Google Scholar]
- Cranshaw, W.S. An Annotated Bibliography of Potato/Tomato Psyllid, Paratrioza cockerelli (Sulc) (Homoptera; Psyllidae). Color. State Univ. Agric. Exp. Stn. Bull. 1993, TB93-5, 51. [Google Scholar]
- Jackson, B.; Goolsby, J.; Wyzykowski, A.; Vitovksy, N.; Bextine, B. Analysis of genetic relationships between potato psyllid (Bactericera cockerelli) populations in the United States, Mexico and Guatemala using ITS2 and Inter Simple Sequence. Subtrop. Plant Sci. 2009, 61, 1–5. [Google Scholar]
- Espinoza, H.R. Facing the Bactericera cockerelli/Candidatus liberibacter complex in Honduras. In Proceedings of the 10th Annual Zebra Chip Reporting Session, Dallas, TX, USA, 7–10 November 2010; pp. 42–46. [Google Scholar]
- Castillo Carrillo, C.; Fu, Z.; Burckhardt, D. First record of the tomato potato Psyllid Bactericera cockerelli from South America. Bull. Insectol. 2019, 72, 85–91. [Google Scholar]
- Liefting, L.W.; Perez-Egusquiza, Z.C.; Clover, G.R.G.; Anderson, J.A.D. A new ‘Candidatus liberibacter’ species in Solanum tuberosum in New Zealand. Plant Dis. 2008, 92, 1474. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Kawchuk, L.; Meers, S. Can Psyll net. Newsletter of the Canadian Potato Psyllid and Zebra Chip Monitoring Network. Available online: http://scholar.ulethbridge.ca/sites/default/files/danjohnson/fil%0Aes/can-psyll-net-dec2017.pdf (accessed on 3 June 2022).
- Walker, P.W.; Allen, G.R.; Tegg, R.S.; White, L.R.; Wilson, C.R. The tomato potato psyllid, Bactericera cockerelli (Sulc, 1909) (Hemiptera: Triozidae): A review of the threat of the psyllid to Australian solanaceous crop industries and surveillance for incursions in potato crops. Austral Entomol. 2015, 54, 339–349. [Google Scholar] [CrossRef]
- Richards, B.L. Further studies with psyllid yellows of the potato. Phytopathology 1931, 21, 103. [Google Scholar]
- Blood, H.L.; Richards, B.L.; Wann, F.B. Studies of psyllid yellows of tomato. Phytopathology 1933, 23, 930. [Google Scholar]
- Eyer, J.R.; Crawford, R.F. Observations on the feeding habits of the potato psyllid (Paratrioza cockerelli Sulc.) and the pathological history of the “psyllid yellows” which it produces. J. Econ. Entomol. 1933, 26, 846–850. [Google Scholar] [CrossRef]
- Richards, B.L.; Blood, H.L. Psyllid yellows of the potato. J. Agric. Res. 1933, 46, 189–216. [Google Scholar]
- Eyer, J.R. Physiology of psyllid yellows of potatoes. J. Econ. Entomol. 1937, 30, 891–898. [Google Scholar] [CrossRef]
- Daniels, L.B. The Nature of the Toxicogenic Condition Resulting from the Feeding of the Tomato Psyllid Paratrioza cockerelli (Sulc). Ph.D. Thesis, University of Minnesota ProQuest Dissertations Publishing, St. Paul, MN, USA, 1954. [Google Scholar]
- Arslan, A.; Bessey, P.M.; Matsuda, K.; Oebker, N.F. Physiological effects of psyllid (Paratrioza cockerelli) on potato. Am. Potato J. 1985, 62, 9–22. [Google Scholar] [CrossRef]
- Goolsby, J.A.; Bextine, B.; Munyaneza, J.E.; Setamou, M.; Adamczyk, J.; Bester, G. Seasonal abundance of sharpshooters, leafhoppers, and psyllids associated with potatoes affected by zebra chip disorder. Subtrop. Plant Sci. 2007, 59, 15–23. [Google Scholar]
- Munyaneza, J.E.; Crosslin, J.M.; Upton, J.E. Association of Bactericera cockerelli (Homoptera: Psyllidae) with “zebra chip,” a new potato disease in southwestern United States and Mexico. J. Econ. Entomol. 2007, 100, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Trumble, J.T.; Stouthamer, R.; Paine, T.D. A new huanglongbing species, “Candidatus liberibacter psyllaurous”, found to infect tomato and potato, is vectored by the psyllid Bactericera cockerelli (Sulc). Appl. Environ. Microbiol. 2008, 74, 5862–5865. [Google Scholar] [CrossRef] [PubMed]
- Secor, G.A.; Rivera-Varas, V.V. Emerging diseases of cultivated potato and their impact on Latin America. Rev. Latinoam. Papa 2004, 1, 1–8. [Google Scholar]
- Liefting, L.W.; Sutherland, P.W.; Ward, L.I.; Paice, K.L.; Weir, B.S.; Clover, G.R.G. A new ‘Candidatus liberibacter’ species associated with diseases of solanaceous crops. Plant Dis. 2009, 93, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Munyaneza, J.E.; Sengoda, V.G.; Crosslin, J.M.; Garzon-Tiznado, J.A.; Cardenas-Valenzuela, O.G. First report of “Candidatus liberibacter solanacearum” in tomato plants in Mexico. Plant Dis. 2009, 93, 1076. [Google Scholar] [CrossRef] [PubMed]
- Munyaneza, J.E. Zebra Chip Disease of Potato: Biology, Epidemiology, and Management. Am. J. Potato Res. 2012, 89, 329–350. [Google Scholar] [CrossRef]
- Munyaneza, J.E. Zebra Chip Disease, Candidatus Liberibacter, and Potato Psyllid: A Global Threat to the Potato Industry. Am. J. Potato Res. 2015, 92, 230–235. [Google Scholar] [CrossRef]
- Bextine, B.; Arp, A.; Flores, E.; Aguilar, E.; Lastrea, L.; Gomez, F.S.; Powell, C.; Rueda, A. First report of zebra chip and ‘Candidatus liberibacter solanacearum’on potatoes in Nicaragua. Plant Dis. 2013, 97, 1109. [Google Scholar] [CrossRef]
- Munyaneza, J.E.; Sengoda, V.G.; Aguilar, E.; Bextine, B.R.; McCue, K.F. First report of ‘Candidatus liberibacter solanacearum’ infecting eggplant in Honduras. Plant Dis. 2013, 97, 1654. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Rashed, A. Biology, Ecology, and Management of the Potato Psyllid, Bactericera cockerelli (Hemiptera: Triozidae), and Zebra Chip Disease in Potato. Annu. Rev. Entomol. 2024, 69, 139–157. [Google Scholar] [CrossRef]
- Goolsby, J.A.; Adamczyk, J.; Bextine, B.; Lin, D.; Munyaneza, J.E.; Bester, G. Development of an IPM program for management of the potato psyllid to reduce incidence of zebra chip disorder in potatoes. Subtrop. Plant Sci. 2007, 59, 85–94. [Google Scholar]
- Munyaneza, J.E.; Goolsby, J.A.; Crosslin, J.M.; Upton, J.E. Further evidence that zebra chip potato disease in the lower Rio Grande Valley of Texas is associated with Bactericera cockerelli. Subtrop. Plant Sci. 2007, 59, 30–37. [Google Scholar]
- Szczepaniec, A.; Varela, K.A.; Kiani, M.; Paetzold, L.; Rush, C.M. Incidence of resistance to neonicotinoid insecticides in Bactericera cockerelli across Southwest US. Crop Prot. 2019, 116, 188–195. [Google Scholar] [CrossRef]
- Swisher, K.D.; Henne, D.C.; Crosslin, J.M. Identification of a fourth haplotype of Bactericera cockerelli (Hemiptera: Triozidae) in the United States. J. Insect Sci. 2014, 14, 161. [Google Scholar] [CrossRef] [PubMed]
- Henne, D.; Bradshaw, J.D.; Schuster, G. Overview of the 2011-2012 Potato Psyllid Areawide Monitoring Program. In Proceedings of the 14th Annual SCRI Zebra Chip Reporting Session, San Antonio, TX, USA, 30 October–12 November 2012. [Google Scholar]
- Rush, C.M.; Workneh, F.; Rashed, A. Significance and epidemiological aspects of late-season infections in the management of potato zebra chip. Phytopathology 2015, 105, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Nachappa, P.; Levy, J.; Pierson, E.; Tamborindeguy, C. Diversity of endosymbionts in the potato psyllid, Bactericera cockerelli (Hemiptera: Triozidae), vector of zebra chip disease of potato. Curr. Microbiol. 2011, 62, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Nakaishi, K.; Fukui, Y.; Arakawa, R. Reproduction of Nesidicoris tenuis (Reuter) on sesame. Jpn. J. Appl. Entomol. Zool. 2011, 55, 199–205. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Dick, J.T.A.; Dickey, J.W.E.; Coughlan, N.E.; Joyce, P.W.S. The Functional Response Ratio (FRR): Advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biol. Invasions 2019, 21, 2543–2547. [Google Scholar] [CrossRef]
- Bowyer, R.T.; Kie, J.G.; Person, D.K.; Monteith, K.L. Metrics of predation: Perils of predator-prey ratios: Predator-prey ratios. Acta Theriol. 2013, 58, 329–340. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Release rate for a pre-plant application of Nesidicoris tenuis for Bemisia tabaci control in tomato. BioControl 2012, 57, 809–817. [Google Scholar] [CrossRef]
- Liu, D.; Trumble, J.T. Tomato psyllid behavioral responses to tomato plant lines and interactions of plant lines with insecticides. J. Econ. Entomol. 2004, 97, 1078–1085. [Google Scholar] [CrossRef]
- Gharalari, A.H.; Nansen, C.; Lawson, D.S.; Gilley, J.; Munyaneza, J.E.; Vaughn, K. Knockdown mortality, repellency, and residual effects of insecticides for control of adult Bactericera cockerelli (Hemiptera: Psyllidae). J. Econ. Entomol. 2009, 102, 1032–1038. [Google Scholar] [CrossRef]
- Ziaei Madbouni, M.A.; Samih, M.A.; Qureshi, J.A.; Biondi, A.; Namvar, P. Compatibility of insecticides and fungicides with the zoophytophagous mirid predator Nesidicoris tenuis. PLoS ONE 2017, 12, e0187439. [Google Scholar] [CrossRef] [PubMed]
- Perdikis, D.; Kapaxidi, E.; Papadoulis, G. Biological control of insect and mite pests in greenhouse solanaceous crops. Biocontrol 2008, 2, 125–144. [Google Scholar]
- Fytrou, N.; Ilias, A.; Sklivakis, J.; Tsagkarakou, A. Lethal and sublethal effects of selected insecticides on commercially available natural enemies of whiteflies. IOBC-WPRS Bull. 2017, 125, 19–27. [Google Scholar]
- Sawadogo, M.W.; Mano, E.; Régis, B. Nesidicoris tenuis in Burkina Faso: Distribution, predatory capacity and insecticide sensibility. Physiol. Entomol. 2022, 47, 201–208. [Google Scholar] [CrossRef]
- Walker, G.P.; MacDonald, F.H.; Wright, P.J.; Puketapu, A.J.; Gardner-Gee, R.; Connolly, P.G.; Anderson, J.A.D. Development of Action Thresholds for Management of Bactericera cockerelli and Zebra Chip Disease in Potatoes at Pukekohe, New Zealand. Am. J. Potato Res. 2015, 92, 266–275. [Google Scholar] [CrossRef]
- Rondon, S.; Schreiber, A.; Jensen, A.; Hamm, P.; Munyaneza, J.; Nolte, P.; Olsen, N.; Wenninger, E.; Henne, D.; Wohleb, C.; et al. Potato psyllid vector of zebra chip disease in the pacific northwest. Pac. Northwest Ext. Publ. 2012, 633, 1–8. [Google Scholar]
- Yen, A.L.; Madge, D.G.; Berry, N.A.; Yen, J.D.L. Evaluating the effectiveness of five sampling methods for detection of the tomato potato psyllid, Bactericera cockerelli (Sulc)(Hemiptera: Psylloidea: Triozidae). Aust. J. Entomol. 2013, 52, 168–174. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Yamamura, K. Transformation using (x + 0.5) to stabilize the variance of populations. Popul. Ecol. 1999, 41, 229–234. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Chaves, L.F. An entomologist guide to demystify pseudoreplication: Data analysis of field studies with design constraints. J. Med. Entomol. 2010, 47, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Sharifian, I.; Sabahi, Q.; Khoshabi, J. Functional response of Macrolophus pygmaeus (Rambur) and Nesidicoris tenuis (Reuter) feeding on two different prey species. Arch. Phytopathol. Plant Prot. 2015, 48, 910–920. [Google Scholar] [CrossRef]
- Michailidis, G.; Sfenthourakis, S.; Pitsillou, M.; Seraphides, N. Functional response and multiple predator effects of two generalist predators preying on Tuta absoluta eggs. Pest Manag. Sci. 2018, 74, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, R.A.; Pallini, A.; Venzon, M.; De Souza, O.F.F.; Molina-Rugama, A.J.; De Oliveira, C.L. Functional response of the predator Eriopis connexa (Coleoptera: Coccinellidae) to different prey types. Braz. Arch. Biol. Technol. 2007, 50, 121–126. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Hemerik, L.; Lins, J.C.; Bueno, V.H.P. Functional Responses of three neotropical mirid predators to eggs of Tuta absoluta on tomato. Insects 2016, 7, 34. [Google Scholar] [CrossRef]
- Kalinkat, G.; Schneider, F.D.; Digel, C.; Guill, C.; Rall, B.C.; Brose, U. Body masses, functional responses and predator-prey stability. Ecol. Lett. 2013, 16, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Bressendorff, B.B.; Toft, S. Dome-shaped functional response induced by nutrient imbalance of the prey. Biol. Lett. 2011, 7, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Kreuzinger-Janik, B.; Brüchner- Hüttemann, H.; Traunspurge, W. Effect of prey size and structural complexity on the functional response in a nematode- nematode system. Sci. Rep. 2019, 9, 5696. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, E.R.; Saville, D.J.; van Koten, C.; Wratten, S.D.; Goldson, S.L. Potential of the mirid bug, Engytatus nicotianae, for the biological control of the tomato-potato psyllid in greenhouses. Crop Prot. 2022, 156, 105941. [Google Scholar] [CrossRef]
- Hassanpour, M.; Bagheri, M.; Golizadeh, A.; Farrokhi, S. Functional response of Nesidicoris tenuis (Hemiptera: Miridae) to Trialeurodes vaporariorum (Hemiptera: Aleyrodidae): Effect of different host plants. Biocontrol Sci. Technol. 2016, 26, 1489–1503. [Google Scholar] [CrossRef]
- Madbouni, M.A.Z.; Samih, M.A.; Namvar, P.; Biondi, A. Temperature-dependent functional response of Nesidicoris tenuis (Hemiptera: Miridae) to different densities of pupae of cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Eur. J. Entomol. 2017, 114, 325–331. [Google Scholar] [CrossRef]
- Pazyuk, I.M.; Musolin, D.L.; Reznik, S.Y. Geographic variation in thermal and photoperiodic effects on development of zoophytophagous plant bug Nesidicoris tenuis. J. Appl. Entomol. 2014, 138, 36–44. [Google Scholar] [CrossRef]
- Walker, G.P.; MacDonald, F.H.; Puketapu, A.J.; Wright, P.J.; Connolly, P.G.; Anderson, J.A.D. A field trial to assess action thresholds for management of Bactericera cockerelli in main crop processing potatoes at Pukekohe. N. Z. Plant Prot. 2013, 66, 349–355. [Google Scholar] [CrossRef]
- Villanueva, R.T.; Esparza-Diaz, G.; Welbourn, C.W. Phytoseiids as potential natural enemies of potato psyllids in organic potato production in South Texas, USA. J. Acarol. Soc. Jpn. 2016, 25, S137–S145. [Google Scholar] [CrossRef]
- Velázquez-Rodríguez, Y.B.; Morales-Alonso, S.I.; Pineda-Guillermo, S.; Figueroa-De-la-Rosa, I.H.-A.A.; Martínez-Castillo, A.M. Capacidad de consumo de Engytatus varians (Distant)(Hemiptera: Miridae) sobre ninfas de Bactericera cockerelli (Sulc.)(Hemiptera: Triozidae). Entomol. Mex. 2015, 2, 235–240. [Google Scholar]
- Martínez, A.M.; Baena, M.; Figueroa, J.I.; Del Estal, P.; Medina, M.; Guzmán-Lara, E.; Pineda, S. Primer registro de Engytatus varians (Distant) (Hemiptera: Heteroptera: Miridae) en México y su depredación sobre Bactericera cockerelli (Šulc) (Hemiptera: Triozidae): Una revisión de su distribución y hábitos. Biocontrol 2014, 30, 617–624. [Google Scholar]
- Calvo, F.J.; Torres-Ruiz, A.; Velázquez-González, J.C.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R. Evaluation of Dicyphus hesperus for biological control of sweet potato whitefly and potato psyllid on greenhouse tomato. BioControl 2016, 61, 415–424. [Google Scholar] [CrossRef]
- Arnó, J.; Castañé, C.; Riudavets, J.; Gabarra, R. Risk of damage to tomato crops by the generalist zoophytophagous predator Nesidiocoris tenuis (Reuter) (Hemiptera: Miridae). Bull. Entomol. Res. 2010, 100, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, D.R.; McGregor, R.R. The functions of plant feeding in the omnivorous predator Dicyphus hesperus: Water places limits on predation. Ecol. Entomol. 2000, 25, 380–386. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Urbaneja-Bernat, P.; Jaques, J.A.; Flors, V.; Urbaneja, A. Defensive plant responses induced by Nesidiocoris tenuis (Hemiptera: Miridae) on tomato plants. J. Pest Sci. 2015, 88, 543–554. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insecticide * | Formulation ** | Foliar Application | Field Nesidiocoris tenuis Releases | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Quantity | Rate | Unit *** | Dates | No. Adults < 4-d (Sex Ratio 1:1) | Dates | ||||
| 20 | 4 February 2016 | 30 January 2017 | |||||||
| 50 | 11 February 2016 | 5 February 2017 | |||||||
| Spirotetramat | SC 2.00 LG | 5 | ZMA | 1 | 12 February 2016 | 2 February 2017 | |||
| 50 | 19 February 2016 | 13 February 2017 | |||||||
| Abamectin | EC 0.15 LG | 8 | ZMA | 2 | 2 March 2016 | 17 February 2017 | |||
| 50 | 9 March 2016 | 24 February 2017 | |||||||
| 20 | 13 March 2017 | ||||||||
| Pymetrozine | WG 50.00% | 5.5 | OMA | 3 | 21 March 2016 | 3 March 2017 | |||
| 50 | 28 March 2016 | 24 March 2017 | |||||||
| 50 | 7 April 2017 | ||||||||
| Spinetoram J and L | SC 1.00 LG | 8 | ZMA | 4 | 16 March 2017 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esparza-Diaz, G.; Villanueva, R.T.; Badillo-Vargas, I.E. A Novel Interaction of Nesidiocoris tenuis (Hemiptera: Miridae) as a Biological Control Agent of Bactericera cockerelli (Hemiptera: Triozidae) in Potato. Insects 2024, 15, 261. https://doi.org/10.3390/insects15040261
Esparza-Diaz G, Villanueva RT, Badillo-Vargas IE. A Novel Interaction of Nesidiocoris tenuis (Hemiptera: Miridae) as a Biological Control Agent of Bactericera cockerelli (Hemiptera: Triozidae) in Potato. Insects. 2024; 15(4):261. https://doi.org/10.3390/insects15040261
Chicago/Turabian StyleEsparza-Diaz, Gabriela, Raul T. Villanueva, and Ismael E. Badillo-Vargas. 2024. "A Novel Interaction of Nesidiocoris tenuis (Hemiptera: Miridae) as a Biological Control Agent of Bactericera cockerelli (Hemiptera: Triozidae) in Potato" Insects 15, no. 4: 261. https://doi.org/10.3390/insects15040261
APA StyleEsparza-Diaz, G., Villanueva, R. T., & Badillo-Vargas, I. E. (2024). A Novel Interaction of Nesidiocoris tenuis (Hemiptera: Miridae) as a Biological Control Agent of Bactericera cockerelli (Hemiptera: Triozidae) in Potato. Insects, 15(4), 261. https://doi.org/10.3390/insects15040261