
Insecticide Susceptibilities and Enzyme Activities of Four Stink Bug Populations in Mississippi, USA



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collections
2.2. Insecticides
2.3. Laboratory Spray Bioassays Containing Bean Pods (Phaseolus vulgaris L.)
2.4. Detoxification Enzyme Activity Assays
2.4.1. Chemicals
2.4.2. Enzyme Preparation
2.4.3. Esterase Activity Assays
2.4.4. Glutathione S-Transferase (GST) Activity Assays
2.4.5. Acetylcholinesterase (AChE) Activity Assay
2.4.6. Cytochrome P450 Monooxygenase (P450) Assays
2.5. Data Analysis
3. Results
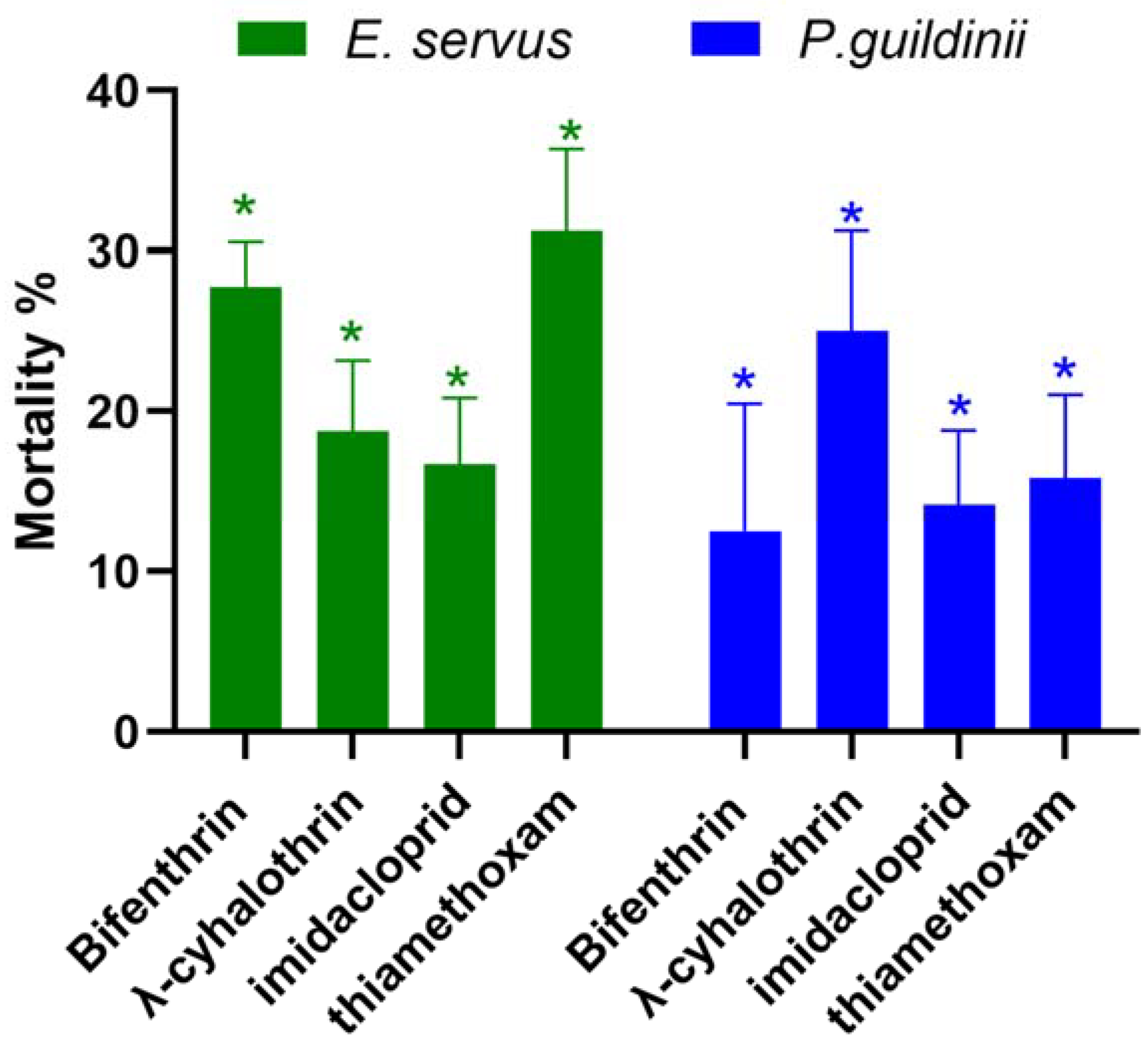
3.1. Spray Bioassay in SIMRU-2022
3.2. Spray Bioassay in SIMRU-2023
3.3. Spray Bioassay in Clarksdale-2023
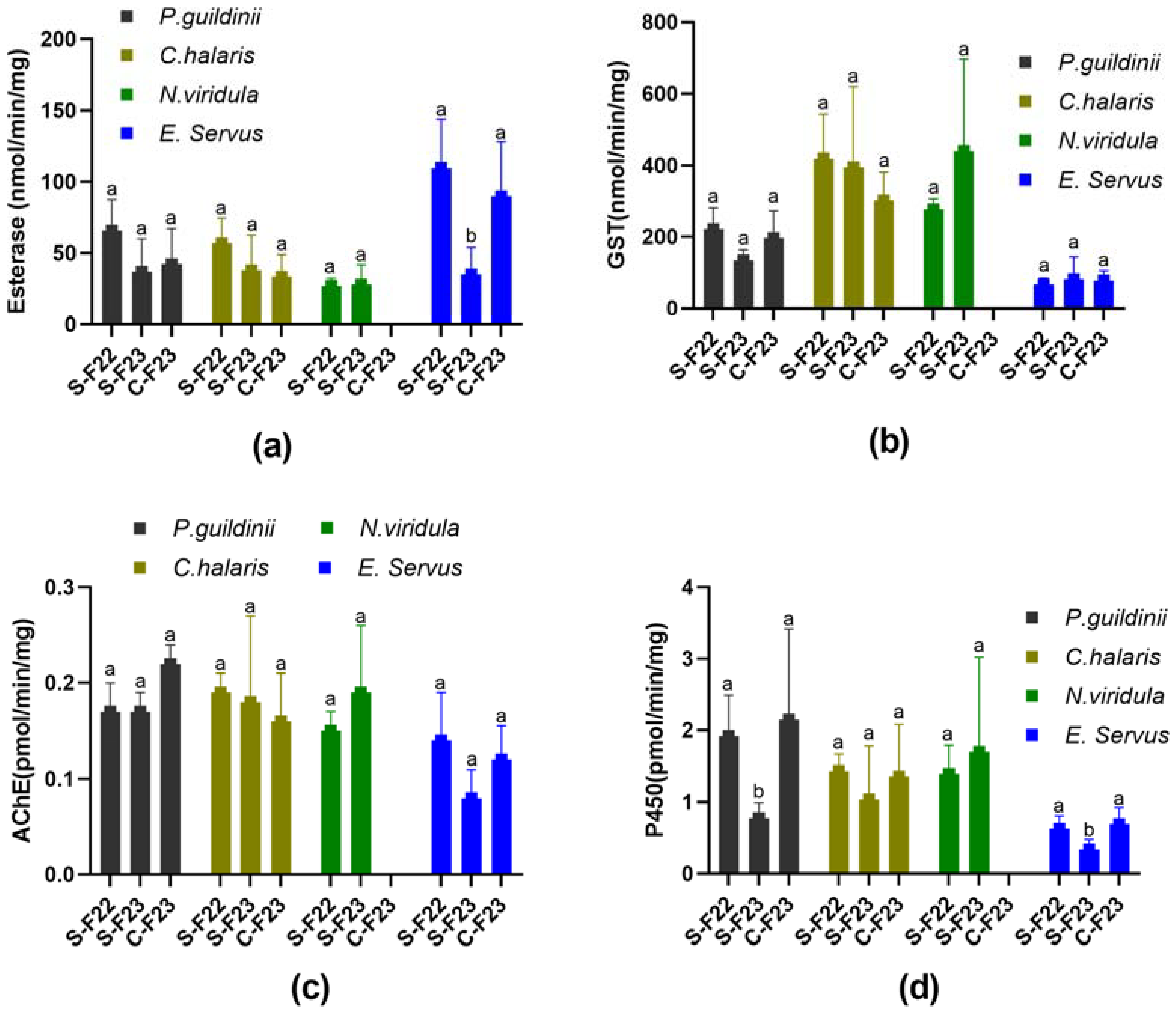
3.4. Detoxification Enzymes and AChE Activity Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McPherson, J.E.; McPherson, R. Stink Bugs of Economic Importance in America North of Mexico; CRC Press: Boca Raton, FL, USA, 2000; 272p. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Slansky, F. Review of phytophagous Pentatomids (Hemiptera: Pentatomidae) associated with soybean in the Americas. Fla. Entomol. 1985, 68, 184–214. [Google Scholar] [CrossRef]
- Panizzi, A.R.; McPherson, J.; James, D.G.; Javahery, M.; McPherson, R.M. Stink bugs (Pentatomidae). In Heteroptera of Economic Importance; CRC Press: Boca Raton, FL, USA, 2000; 828p. [Google Scholar]
- McPherson, J.E. Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Musser, F.; Catchot, A., Jr.; Conley, S.; Davis, J.; DiFonzo, C.; Greene, J.; Lorenz, G.; Owens, D.; Reed, T.; Reisig, D. Soybean insect losses in the United States. Midsouth. Entomol. 2019, 13, 1–23. [Google Scholar]
- McPherson, R.; Douce, G.; Hudson, R. Annual variation in stink bug (Heteroptera: Pentatomidae) seasonal abundance and species composition in Georgia soybean and its impact on yield and quality. J. Entomol. Sci. 1993, 28, 61–72. [Google Scholar] [CrossRef]
- Depieri, R.; Panizzi, A. Duration of feeding and superficial and in-depth damage to soybean seed by selected species of stink bugs (Heteroptera: Pentatomidae). Neotrop. Entomol. 2011, 40, 197–203. [Google Scholar] [CrossRef]
- Bundy, C.S.; Esquivel, J.F.; Panizzi, A.R.; Eger, J.E.; Davis, J.A.; Jones, W.A. Piezodorus guildinii (Westwood). In Invasive Stink Bugs and Related Species (Pentatomoidea); CRC Press: Boca Raton, FL, USA, 2018; pp. 425–452. [Google Scholar]
- Souza, E.; Silva, J.; Baldin, E.; Pierozzi, C.; Cunha, L.; Canassa, V.; Pannuti, L.; Lourenção, A. Response of soybean genotypes challenged by a stink bug complex (Hemiptera: Pentatomidae). J. Econ. Entomol. 2016, 109, 898–906. [Google Scholar] [CrossRef]
- Baur, M.E.; Sosa-Gomez, D.R.; Ottea, J.; Leonard, B.R.; Corso, I.C.; Da Silva, J.J.; Temple, J.; Boethel, D.J. Susceptibility to insecticides used for control of Piezodorus guildinii (Heteroptera: Pentatomidae) in the United States and Brazil. J. Econ. Entomol. 2010, 103, 869–876. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Adamczyk, J.J., Jr.; Gore, J. Toxicity of insecticides in a glass-vial bioassay to adult brown, green, and southern green stink bugs (Heteroptera: Pentatomidae). J. Econ. Entomol. 2005, 98, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Gómez, D.R.; Corrêa-Ferreira, B.S.; Kraemer, B.; Pasini, A.; Husch, P.E.; Delfino Vieira, C.E.; Reis Martinez, C.B.; Negrão Lopes, I.O. Prevalence, damage, management and insecticide resistance of stink bug populations (Hemiptera: Pentatomidae) in commodity crops. Agr. Forest Entomol. 2020, 22, 99–118. [Google Scholar] [CrossRef]
- Emfinger, K.; Leonard, B.R.; Gore, J.; Cook, D. Insecticide toxicity to southern green, Nezara viridula (L.), and brown, Euschistus servus (Say), stink bugs. In Proceedings of the Beltwide Cotton Conferences, Memphis, TN, USA, 9–12 January 2001; pp. 1159–1161. [Google Scholar]
- Temple, J.; Davis, J.; Hardke, J.; Moore, J.; Leonard, B. Susceptibility of southern green stink bug and redbanded stink bug to insecticides in soybean field experiments and laboratory bioassays. Southwest. Entomol. 2013, 38, 393–406. [Google Scholar] [CrossRef]
- Hegeto, L.A.; Ronqui, L.; Lapenta, A.S.; Albuquerque, F.A. Identification and functional characterization of esterases in Euschistus heros (Hemiptera, Pentatomidae) and their relationship with thiamethoxam and lambda-cyhalothrin. Genet. Mol. Res. 2015, 14, 11079–11088. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, C.E.; Miller, J.L.; Miller, M.J.; Gee, S.J.; Shan, G.; Hammock, B.D. Development of toxicity identification evaluation procedures for pyrethroid detection using esterase activity. Environ. Toxicol. Chem. Int. J. 2004, 23, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Wilce, M.C.; Parker, M.W. Structure and function of glutathione S-transferases. Biochem. Biophys. Acta 1994, 1205, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Field, L.M. Gene amplification and insecticide resistance. Pest Manag. Sci. 2011, 67, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Common and uncommon cytochrome P450 reactions related to metabolism and chemical toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef] [PubMed]
- Mittapelly, P.; Bansal, R.; Michel, A. Differential expression of cytochrome P450 CYP6 genes in the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2019, 112, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, G. Status of resistance of resistance in tarnished plant bug. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 3–6 January 2006. [Google Scholar]
- Du, Y.; Zhu, Y.C.; Portilla, M.; Zhang, M.; Reddy, G.V.P. The mechanisms of metabolic resistance to pyrethroids and neonicotinoids fade away without selection pressure in the tarnished plant bug Lygus lineolaris. Pest Manag. Sci. 2023, 79, 3893–3902. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Feeding toxicity and impact of imidacloprid formulation and mixtures with six representative pesticides at residue concentrations on honey bee physiology (Apis mellifera). PLoS ONE 2017, 12, e0178421. [Google Scholar] [CrossRef] [PubMed]
- Dorman, S.J.; Gross, A.D.; Musser, F.R.; Catchot, B.D.; Smith, R.H.; Reisig, D.D.; Reay-Jones, F.P.; Greene, J.K.; Roberts, P.M.; Taylor, S.V. Resistance monitoring to four insecticides and mechanisms of resistance in Lygus lineolaris Palisot de Beauvois (Hemiptera: Miridae) populations of southeastern USA cotton. Pest Manag. Sci. 2020, 76, 3935–3944. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Van Pottelberge, S.; Tirry, L. Biochemical analysis of a chlorfenapyr-selected resistant strain of Tetranychus urticae Koch. Pest Manag. Sci. 2006, 62, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Endo, N. Insecticide susceptibility of Nezara viridula (Heteroptera: Pentatomidae) and three other stink bug species composing a soybean pest complex in Japan. J. Econ. Entomol. 2012, 105, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Willrich, M.M.; Leonard, B.R.; Cook, D.R. Laboratory and field evaluations of insecticide toxicity to stink bugs (Heteroptera: Pentatomidae). J. Cotton Sci. 2003, 7, 156–163. [Google Scholar]
- Layton, M.B. Biology and damage of the tarnished plant bug, Lygus lineolaris, in cotton. Southwest. Entomol. 2000, 23, 7–20. [Google Scholar]
- Steinhaus, E.A.; Warpechowski, L.F.; Braga, L.E.; Morin, M.; Tenório, C.; Boff, J.S.; Bernardi, O.; Farias, J.R. Intra- and interspecific variation in the susceptibility to insecticides of stink bugs (Hemiptera: Pentatomidae) That attack soybean and maize in southern Brazil. J. Econ. Entomol. 2022, 115, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, T.P.; Doughty, H.; Kamminga, K.; Wallingford, A.; Philips, C.; Aigner, J. Evaluation of insecticides for the control of brown marmorated stink bug in bell peppers in Virginia experiment 2, 2011. Arthropod Manag. Tests 2012, 37, E40. [Google Scholar]
- Lee, D.-H.; Wright, S.E.; Leskey, T.C. Impact of insecticide residue exposure on the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae): Analysis of adult mobility. J. Econ. Entomol. 2013, 106, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Somavilla, J.C.; Reis, A.C.; Gubiani, P.d.S.; Godoy, D.N.; Stürmer, G.R.; Bernardi, O. Susceptibility of Euschistus heros and Dichelops furcatus (Hemiptera: Pentatomidae) to selected insecticides in Brazil. J. Econ. Entomol. 2020, 113, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Tibola, C.M.; Silva, L.; Sgubin, F.; Omoto, C. Monitoring resistance of Euschistus heros (Fabricius) (Hemiptera: Pentatomidae) to insecticides by using encapsulated artificial diet bioassay. Insects 2021, 12, 599. [Google Scholar] [CrossRef] [PubMed]
- Sosa-GóMez, D.R.; Da Silva, J.J.; De Oliveira Negrao Lopes, I.; Corso, I.C.; Almeida, A.M.; Piubelli De Moraes, G.C.; Baur, M.E. Insecticide susceptibility of Euschistus heros (Heteroptera: Pentatomidae) in Brazil. J. Econ. Entomol. 2009, 102, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Campos, S.; Smagghe, G.; Fragoso, D.; Barrigossi, J.; Guedes, R. Area-wide survey of thiamethoxam resistance and control failure likelihood in the rice stink bugs Oebalus poecilus and O. ypsilongriseus. J. Pest Sci. 2022, 95, 1151–1161. [Google Scholar] [CrossRef]
- Tuelher, E.S.; da Silva, É.H.; Rodrigues, H.S.; Hirose, E.; Guedes, R.N.C.; Oliveira, E.E. Area-wide spatial survey of the likelihood of insecticide control failure in the neotropical brown stink bug Euschistus heros. J. Pest Sci. 2018, 91, 849–859. [Google Scholar] [CrossRef]
- Castellanos, N.L.; Haddi, K.; Carvalho, G.A.; de Paulo, P.D.; Hirose, E.; Guedes, R.N.C.; Smagghe, G.; Oliveira, E.E. Imidacloprid resistance in the Neotropical brown stink bug Euschistus heros: Selection and fitness costs. J. Pest Sci. 2019, 92, 847–860. [Google Scholar] [CrossRef]
- Santos, T.L.B.; Baldin, E.L.L.; Lima, A.P.S.; Santana, A.S.; Santos, M.C.; Silveira, B.R.R.; Bueno, N.M.; Cabral, I.R.; Soares, M.C.E.; Pinheiro, A.M.; et al. Intraspecific and interspecific interaction and fitness cost of stink bugs Euschistus heros, Diceraeus melacanthus, and Piezodorus guildinii in soybean. Pest Manag. Sci. 2024, 80, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Lira, E.C.; Nascimento, A.R.; Bass, C.; Omoto, C.; Cônsoli, F.L. Transcriptomic investigation of the molecular mechanisms underlying resistance to the neonicotinoid thiamethoxam and the pyrethroid lambda-cyhalothrin in Euschistus heros (Hemiptera: Pentatomidae). Pest Manag. Sci. 2023, 79, 5349–5361. [Google Scholar] [CrossRef] [PubMed]
- Ademokoya, B.; Athey, K.; Ruberson, J. Natural enemies and biological control of stink bugs (Hemiptera: Heteroptera) in north America. Insects 2022, 13, 932. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compounds 1 | Population | Slope | LC50 (μg/mL) 2 | 95% Confidence Intervals (μg/mL) 2 | χ2 | p |
|---|---|---|---|---|---|---|
| Oxamyl | C. halaris | 2.391 ± 0.395 | 88.84 a | 65.60–116.27 | 0.29 | 0.99 |
| N. viridula | 3.028 ± 0.464 | 140.10 a | 113.79–168.32 | 1.37 | 0.85 | |
| P. guildinii | 3.221 ± 0.331 | 97.55 a | 85.69–117.46 | 6.52 | 0.16 | |
| Acephate | C. halaris | 2.842 ± 0.485 | 157.88 a | 125.03–200.95 | 2.07 | 0.56 |
| N. viridula | 4.783 ± 0.817 | 213.36 a | 183.40–244.66 | 1.59 | 0.45 | |
| P. guildinii | 2.811 ± 0.327 | 48.36 b | 40.61–55.80 | 2.54 | 0.28 | |
| Bifenthrin | C. halaris | 2.011 ± 0.379 | 5.68 a | 3.84–7.92 | 2.35 | 0.50 |
| N. viridula | 2.163 ± 0.383 | 5.57 a | 3.86–7.45 | 3.09 | 0.38 | |
| P. guildinii | 1.944 ± 0.301 | 8.40 a | 6.66–10.53 | 2.80 | 0.25 | |
| λ-Cyhalothrin | C. halaris | 1.339 ± 0.343 | 6.13 a | 2.64–9.99 | 0.70 | 0.95 |
| N. viridula | 2.168 ± 0.598 | 3.14 a | 1.37–4.56 | 0.82 | 0.66 | |
| P. guildinii | 1.378 ± 0.200 | 5.97 a | 4.13–7.78 | 0.43 | 0.93 | |
| Imidacloprid | C. halaris | 2.065 ± 0.405 | 22.17 b | 15.05–31.31 | 1.53 | 0.68 |
| N. viridula | 2.074 ± 0.364 | 74.58 a | 57.04–103.23 | 3.84 | 0.28 | |
| P. guildinii | 1.770 ± 0.222 | 17.39 b | 13.74–21.24 | 3.78 | 0.29 | |
| Thiamethoxam | C. halaris | 2.077 ± 0.534 | 13.67 a | 8.62–19.18 | 1.40 | 0.50 |
| N. viridula | 1.768 ± 0.366 | 15.22 a | 10.12–20.81 | 0.28 | 0.96 | |
| P. guildinii | 1.688 ± 0.233 | 11.45 a | 8.57–14.23 | 0.40 | 0.94 | |
| Sulfoxaflor | C. halaris | 2.711 ± 0.545 | 174.04 a | 129.55–227.03 | 2.44 | 0.49 |
| N. viridula | 2.663 ± 0.422 | 223.62 a | 180.86–287.57 | 3.43 | 0.49 | |
| P. guildinii | 1.272 ± 0.263 | 90.52 b | 66.84–128.10 | 0.88 | 0.83 |
| Compounds 1 | Population | Slope | LC50 (μg/mL) 2 | 95% Confidence Intervals (μg/mL) 2 | χ2 | p |
|---|---|---|---|---|---|---|
| Oxamyl | C. halaris | 2.085 ± 0.491 | 98.89 b | 73.57–131.60 | 1.12 | 0.29 |
| E. servus | 1.801 ± 0.612 | 356.56 a | 237.91–714.94 | 2.41 | 0.30 | |
| P. guildinii | 3.109 ± 0.447 | 87.97 b | 73.20–102.35 | 0.12 | 0.73 | |
| Acephate | C. halaris | 2.400 ± 0.973 | 163.21 a | 73.70–256.43 | 2.07 | 0.56 |
| E. servus | 4.958 ± 1.405 | 269.96 a | 209.31–312.61 | 1.31 | 0.25 | |
| P. guildinii | 3.668 ± 0.531 | 43.51 b | 36.54–50.10 | 0.41 | 0.52 | |
| Bifenthrin | C. halaris | 2.121 ± 0.411 | 9.07 b | 6.85–11.60 | 1.85 | 0.17 |
| E. servus | 2.980 ± 0.512 | 31.07 a | 23.91–39.74 | 0.47 | 0.79 | |
| P. guildinii | 0.897 ± 0.312 | 10.11 b | 5.61–18.79 | 0.03 | 0.85 | |
| λ-Cyhalothrin | C. halaris | 1.343 ± 0.289 | 7.22 b | 4.46–10.68 | 0.01 | 0.99 |
| E. servus | 1.972 ± 0.425 | 45.28 a | 32.54–67.52 | 0.14 | 0.93 | |
| P. guildinii | 2.360 ± 0.465 | 7.58 b | 5.37–9.49 | 0.07 | 0.80 | |
| Imidacloprid | C. halaris | 3.783 ± 0.905 | 23.77 b | 17.31–31.61 | 0.11 | 0.74 |
| E. servus | 1.763 ± 0.444 | 139.53 a | 93.57–329.16 | 1.47 | 0.48 | |
| P. guildinii | 1.281 ± 0.237 | 8.87 c | 5.50–11.40 | 0.43 | 0.51 | |
| Thiamethoxam | C. halaris | 1.567 ± 0.278 | 12.22 b | 8.01–17.01 | 1.82 | 0.34 |
| E. servus | 2.084 ± 0.500 | 94.36 a | 68.16–139.57 | 0.43 | 0.81 | |
| P. guildinii | 1.191 ± 0.211 | 5.18 c | 2.90–7.55 | 0.67 | 0.72 | |
| Sulfoxaflor | C. halaris | 6.666 ± 1.762 | 276.09 b | 231.43–320.99 | 0.12 | 0.73 |
| E. servus | 5.638 ± 2.191 | 425.08 a | 363.36–607.45 | 1.51 | 0.22 | |
| P. guildinii | 0.998 ± 0.234 | 61.95 c | 37.78–86.06 | 1.80 | 0.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Scheibener, S.; Zhu, Y.-C.; Allen, K.C.; Reddy, G.V.P. Insecticide Susceptibilities and Enzyme Activities of Four Stink Bug Populations in Mississippi, USA. Insects 2024, 15, 265. https://doi.org/10.3390/insects15040265
Du Y, Scheibener S, Zhu Y-C, Allen KC, Reddy GVP. Insecticide Susceptibilities and Enzyme Activities of Four Stink Bug Populations in Mississippi, USA. Insects. 2024; 15(4):265. https://doi.org/10.3390/insects15040265
Chicago/Turabian StyleDu, Yuzhe, Shane Scheibener, Yu-Cheng Zhu, K. Clint Allen, and Gadi V. P. Reddy. 2024. "Insecticide Susceptibilities and Enzyme Activities of Four Stink Bug Populations in Mississippi, USA" Insects 15, no. 4: 265. https://doi.org/10.3390/insects15040265
APA StyleDu, Y., Scheibener, S., Zhu, Y. -C., Allen, K. C., & Reddy, G. V. P. (2024). Insecticide Susceptibilities and Enzyme Activities of Four Stink Bug Populations in Mississippi, USA. Insects, 15(4), 265. https://doi.org/10.3390/insects15040265