Sexual Communication in the Drosophila Genus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
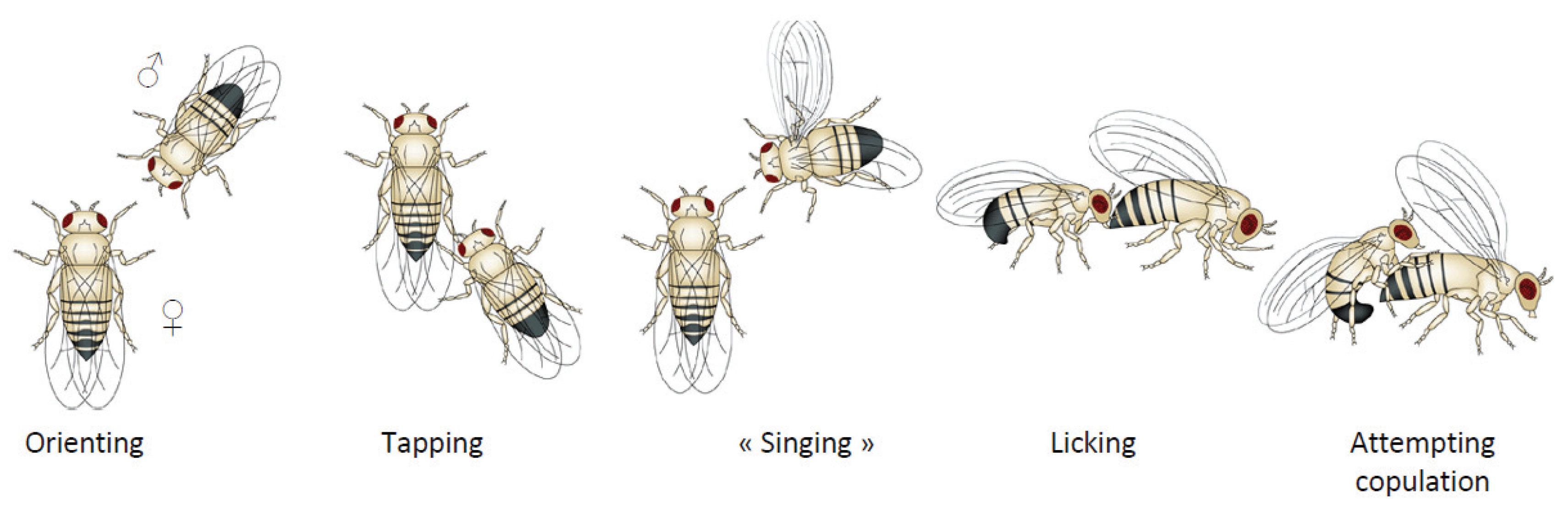
:1. Drosophila Courtship Behavior

2. Non-Chemical Stimuli
2.1. Acoustic Stimuli

2.2. Visual Stimuli
3. Sex Pheromone Signals
3.1. Cuticular Hydrocarbons-Generalities

3.2. Pheromonal Role of CHCs

3.3. Pheromonal Role of CHCs from Immature Flies
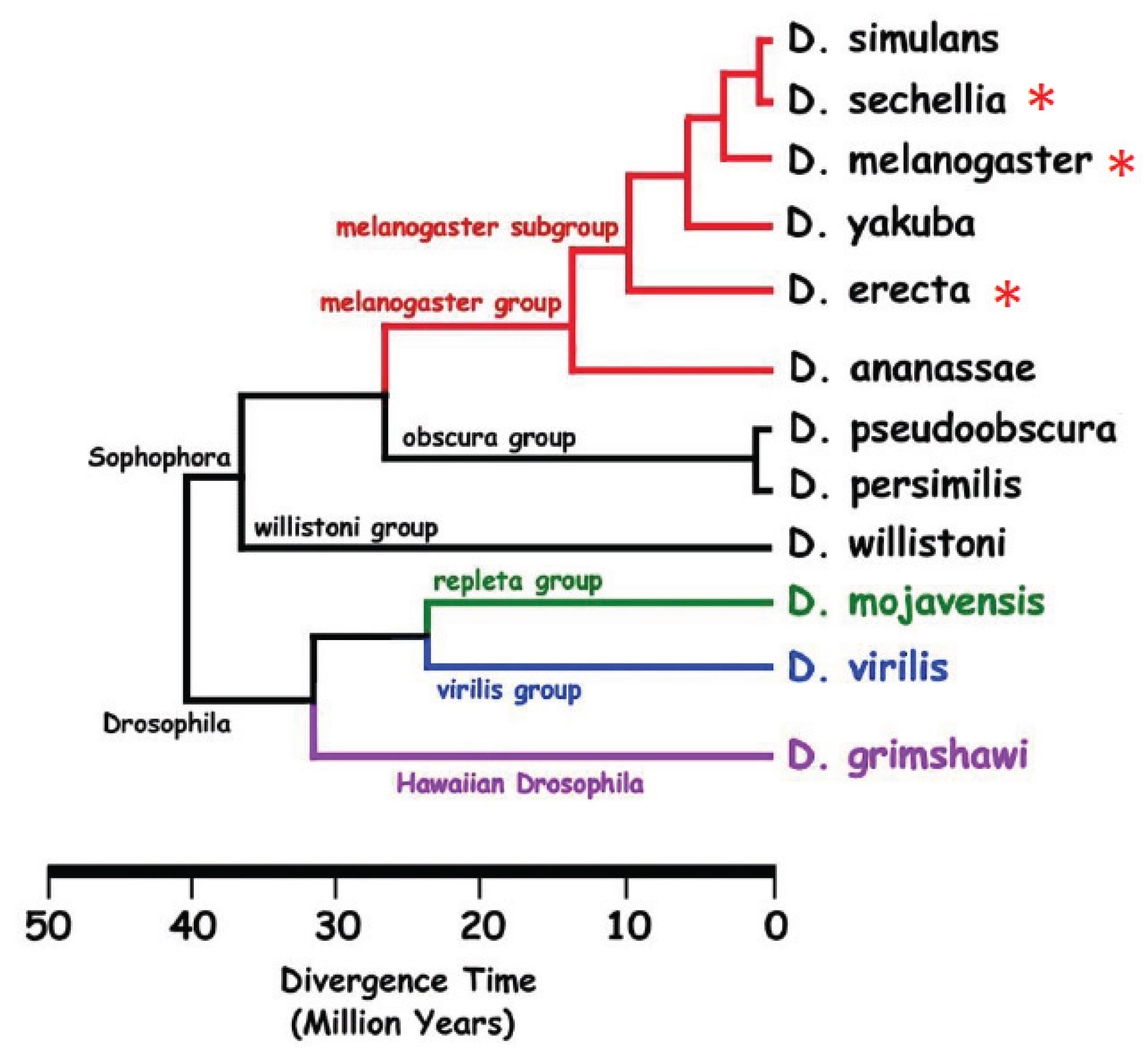
4. Role of CHCs in Reproductive Isolation
4.1. Role of Female Pheromones in D. melanogaster Reproductive Isolation
4.2. Role of Male Pheromones in D. melanogaster Reproductive Isolation
5. Plasticity and Evolution of CHCs
5.1. Plasticity of Hydrocarbons
5.2. Evolution of CHC Profiles
6. Non-Hydrocarbon Pheromones
6.1. cis-Vaccenyl Acetate
6.1.1. Production of cis-Vaccenyl Acetate

6.1.2. Role of cVA in Mating Behavior
6.1.3. Role of cVA as an Aggregation Pheromone
6.1.4. Other Behavioral Roles of cVA
6.2. Polar Compounds and Triacylglycerides
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Markow, T.A.; O’Grady, P.M. Evolutionary genetics of reproductive behavior in Drosophila: Connecting the dots. Annu. Rev. Genet. 2005, 39, 263–291. [Google Scholar] [CrossRef]
- Sturtevant, A.H. Experiments on sex recognition and the problem of sexual selection in Drosophila. J. Anim. Behav. 1915, 5, 351–366. [Google Scholar] [CrossRef]
- Spieth, T.H. Mating behavior within the genus Drosophila (diptera). Am. Museum Nat. Hist. 1952, 99, 395–474. [Google Scholar]
- Hall, J.C. The mating of a fly. Science 1994, 264, 1702–1714. [Google Scholar]
- Greenspan, R.J.; Ferveur, J.F. Courtship in Drosophila. Annu. Rev. Genet. 2000, 34, 205–232. [Google Scholar] [CrossRef]
- Bennet-Clark, H.C.; Ewing, A.W. The love song of the fruit fly. Sci. Am. 1970, 223, 85–90. [Google Scholar]
- Von Schilcher, F. The role of auditory stimuli in Drosophila melanogaster. Anim. Behav. 1976, 24, 18–26. [Google Scholar] [CrossRef]
- Fabre, C.C.; Hedwig, B.; Conduit, G.; Lawrence, P.A.; Goodwin, S.F.; Casal, J. Substrate-borne vibratory communication during courtship in Drosophila melanogaster. Curr. Biol. 2012, 22, 2180–2185. [Google Scholar] [CrossRef]
- Sokolowski, M.B. Drosophila: Genetics meets behaviour. Nat. Rev. Genet. 2001, 2, 879–890. [Google Scholar] [CrossRef]
- Connolly, K.; Cook, R.M. Rejection responses by female D. melanogaster: Their ontogeny, causality, and effects upon the behavior of the courting male. Behaviour 1973, 44, 142–167. [Google Scholar] [CrossRef]
- Lasbleiz, C.; Ferveur, J.F.; Everaerts, C. Courtship behavior of Drosophila melanogaster revisited. Anim. Behav. 2006, 72, 1001–1012. [Google Scholar] [CrossRef]
- Wolfner, M.F. Tokens of love: Functions and regulation of Drosophila male accessory gland products. Insect Biochem. Mol. Biol. 1997, 27, 179–192. [Google Scholar] [CrossRef]
- Wolfner, M.F. The gifts that keep on giving: Physiological functions and evolutionary dynamics of male seminal proteins in Drosophila. Heredity 2002, 88, 85–93. [Google Scholar] [CrossRef]
- Rezaval, C.; Pavlou, H.J.; Dornan, A.J.; Chan, Y.B.; Kravitz, E.A.; Goodwin, S.F. Neural circuitry underlying Drosophila female postmating behavioral responses. Curr. Biol. 2012, 22, 1155–1165. [Google Scholar] [CrossRef]
- Mehren, J.E.; Ejima, A.; Griffith, L.C. Unconventional sex: Fresh approaches to courtship learning. Curr. Opin. Neurobiol. 2004, 14, 745–750. [Google Scholar] [CrossRef]
- Everaerts, C.; Farine, J.P.; Cobb, M.; Ferveur, J.F. Drosophila cuticular hydrocarbons revisited: mating status alters cuticular profiles. PLoS One 2010, 5, e9607. [Google Scholar]
- Cowling, D.E.; Burnet, B. Courtship song and genetic control of their acoustic characteristics in sibling species of D. melanogaster subgroup. Anim. Behav. 1981, 29, 924–935. [Google Scholar] [CrossRef]
- Ewing, A.W.; Bennet-Clark, H.C. The courtship songs of Drosophila. Behaviour 1968, 31, 288–301. [Google Scholar] [CrossRef]
- Kyriacou, C.P.; Hall, J.C. Circadian rhythm mutations in Drosophila melanogaster affect short-term fluctuations in the male’s courtship song. Proc. Natl. Acad. Sci. USA 1980, 77, 6729–6733. [Google Scholar] [CrossRef]
- Kyriacou, C.P.; Hall, J.C. Interspecific genetic control of courtship song production and reception in Drosophila. Science 1986, 232, 494–497. [Google Scholar]
- Ritchie, M.G.; Halsey, E.J.; Gleason, J.M. Drosophila song as a species-specific mating signal and the behavioural importance of Kyriacou & Hall cycles in D. melanogaster song. Anim. Behav. 1999, 58, 649–657. [Google Scholar] [CrossRef]
- Kyriacou, C.P.; Hall, J.C. Song rhythms in Drosophila. Trends Ecol. Evol. 1990, 5, 125–126. [Google Scholar]
- Tootoonian, S.; Coen, P.; Kawai, R.; Murthy, M. Neural representations of courtship song in the Drosophila brain. J. Neurosci. 2012, 32, 787–798. [Google Scholar] [CrossRef]
- Bennet-Clark, H.C.; Ewing, A.W. Stimuli provided by courtship of male Drosophila melanogaster. Nature 1967, 215, 669–671. [Google Scholar] [CrossRef]
- Crossley, S.A.; Bennet-Clark, H.C.; Evert, H.T. Courtship song components affect male and female Drosophila differently. Anim. Behav. 1995, 50, 827–839. [Google Scholar] [CrossRef]
- Cook, R.M. Courtship proceeding in D. melanogaster. Anim. Behav. 1973, 21, 349–358. [Google Scholar] [CrossRef]
- Bennet-Clark, H.C.; Ewing, A.W. Pulse interval as a critical parameter in the courtship song of Drosophila melanogaster. Anim. Behav. 1969, 17, 755–759. [Google Scholar] [CrossRef]
- Kyriacou, C.P. The function of courtship song rhythms in Drosophila. Anim. Behav. 1982, 30, 794–801. [Google Scholar] [CrossRef]
- Tomaru, M.; Matsubayashi, H.; Oguma, Y. Heterospecific inter-pulse intervals of courtship song elicit female rejection in Drosophila biauraria. Anim. Behav. 1995, 50, 905–914. [Google Scholar] [CrossRef]
- Saarikettu, M.; Liimatainen, J.O.; Hoikkala, A. The role of male courtship song in species recognition in Drosophila montana. Behav. Genet. 2005, 35, 257–263. [Google Scholar] [CrossRef]
- Bixler, A.; Jenkins, J.B.; Tompkins, L.; McRobert, P. Identification of acoustic stimuli that mediate sexual behavior in Drosophila busckii (diptera; drosophilidae). J. Insect Behav. 1992, 5, 469–478. [Google Scholar] [CrossRef]
- Liimatainen, J.O.; Hoikkala, A.; Aspi, J.; Welbergen, P. Courtship in Drosophila montana: The effects of male auditory signals on the behaviour of flies. Anim. Behav. 1992, 43, 35–48. [Google Scholar]
- Doi, M.; Matsuda, M.; Tomaru, M.; Matsubayashi, H.; Oguma, Y. A locus for female discrimination behavior causing sexual isolation in Drosophila. Proc. Natl. Acad. Sci. USA 2001, 98, 6714–6719. [Google Scholar] [CrossRef]
- Tomaru, M.; Doi, M.; Higuchi, H.; Oguma, Y. Courtship song recognition in the Drosophila melanogaster complex: Heterospecific songs make females receptive in D. melanogaster, but not in D. sechellia. Evolution 2000, 54, 1286–1294. [Google Scholar] [CrossRef]
- Tomaru, M.; Yamada, H.; Oguma, Y. Female mate recognition and sexual isolation depending on courtship song in Drosophila sechellia and its siblings. Genes Genet. Syst. 2004, 79, 145–150. [Google Scholar] [CrossRef]
- Tomaru, M.; Matsubayashi, H.; Oguma, Y. Effects of courtship song in interspecific crosses among the species of the Drosophila auraria complex (diptera: Drosophilidae). J. Insect Behav. 1998, 11, 383–398. [Google Scholar] [CrossRef]
- Yamada, H.; Sakai, T.; Tomaru, M.; Doi, M.; Matsuda, M.; Oguma, Y. Search for species-specific mating signal in courtship songs of sympatric sibling species, Drosophila ananassae and D. pallidosa. Genes Genet. Syst. 2002, 77, 97–106. [Google Scholar] [CrossRef]
- Hoikkala, A.; Crossley, S.A. Copulatory courtship in Drosophila: Behavior and songs of D. birchii and D. serrata. J. Insect Behav. 2000, 13, 71–86. [Google Scholar] [CrossRef]
- Ritchie, M.G.; Yate, V.H.; Kyriacou, C.P. Genetic variability of the interpulse interval of courtship song among some European populations of Drosophila melanogaster. Heredity 1994, 72, 459–464. [Google Scholar] [CrossRef]
- Ritchie, M.G.; Townhill, R.M.; Hoikkala, A. Female preference for fly song: Playback experiments confirm the targets of sexual selection. Anim. Behav. 1998, 56, 713–717. [Google Scholar] [CrossRef]
- Talyn, B.; Dowse, H. The role of courtship song in sexual selection and species recognition by female Drosophila melanogaster. Anim. Behav. 2004, 68, 1165–1180. [Google Scholar] [CrossRef]
- Ejima, A.; Griffith, L.C. Courtship initiation is stimulated by acoustic signals in Drosophila melanogaster. PLoS One 2008, 3, e3246. [Google Scholar] [CrossRef]
- Donegan, J.; Ewing, A.W. Duetting in Drosophila and Zaprionus species. Anim. Behav. 1980, 28, 1289. [Google Scholar] [CrossRef]
- Hoikkala, A. Genetic variation in the male courtship sound of Drosophila littoralis. Behav. Genet. 1985, 15, 135–142. [Google Scholar] [CrossRef]
- Yamada, H.; Tomaru, M.; Matsuda, M.; Oguma, Y. Behavioral sequence leading to sexual isolation between D. ananassae and D. pallidosa. J. Insect Behav. 2008, 21, 222–239. [Google Scholar] [CrossRef]
- Tompkins, L.; McRobert, S.P.; Kaneshiro, K.Y. Chemical communication in hawaiian Drosophila. Evolution 1993, 47, 1407–1419. [Google Scholar] [CrossRef]
- Gailey, D.A.; Lacaillade, R.C.; Hall, J.C. Chemosensory elements of courtship in normal and mutant, olfaction-deficient Drosophila melanogaster. Behav. Genet. 1986, 16, 375–405. [Google Scholar]
- Cook, R.M. The courtship tracking of D. melanogaster. Biol. Cybern. 1979, 34, 91–106. [Google Scholar]
- Edwards, K.A.; Doescher, L.T.; Kaneshiro, K.Y.; Yamamoto, D.A. Database of wing diversity in the Hawaiian Drosophila. PLoS One 2007, 2, e487. [Google Scholar] [CrossRef]
- Fuyama, Y. A visual stimulus in the courtship of Drosophila suzukii. Experientia 1979, 35, 1327–1328. [Google Scholar] [CrossRef]
- Grossfield, J. The influence of light on the mating behavior of Drosophila. Studies in Genetics III 1966, 6615, 147–176. [Google Scholar]
- Grossfield, J. Geographic distribution and light-dependent behavior in Drosophila. Proc. Natl. Acad. Sci. USA 1971, 68, 2669–2673. [Google Scholar] [CrossRef]
- Ranz, J.M.; Castillo-Davis, C.I.; Meiklejohn, C.D.; Hartl, D.L. Sex-dependent gene expression and evolution of the Drosophila transcriptome. Science 2003, 300, 1742–1745. [Google Scholar] [CrossRef]
- Stocker, R.F. The organization of the chemosensory system in Drosophila melanogaster: A review. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef]
- Boll, W.; Noll, M. The Drosophila Pox neuro gene: Control of male courtship behavior and fertility as revealed by a complete dissection of all enhancers. Development 2002, 129, 5667–5681. [Google Scholar]
- Bray, S.; Amrein, H. A putative Drosophila pheromone receptor expressed in male-specific taste neurons is required for efficient courtship. Neuron 2003, 39, 1019–1029. [Google Scholar] [CrossRef]
- Howard, K. Cuticular Hydrocarbons and Chemical Communication; Stanley-Samuelson, D.W., Nelson, D.D., Eds.; University of Nebraska Press: Lincoln, NE, USA, 1993; pp. 179–226. [Google Scholar]
- Tompkins, L.; Gross, A.C.; Hall, J.C.; Gailey, D.A.; Siegel, R.W. The role of female movement in the sexual behavior of D. melanogaster. Behav. Genet. 1982, 12, 295–307. [Google Scholar] [CrossRef]
- Jallon, J.M. A few chemical words exchanged by Drosophila during courtship and mating. Behav. Genet. 1984, 14, 441–478. [Google Scholar] [CrossRef]
- Antony, C.; Davis, T.L.; Carlson, D.A.; Péchiné, J.M.; Jallon, J.M. Compared behavioral responses of male Drosophila melanogaster (Canton S) to natural and synthetic aphrodisiacs. J. Chem. Ecol. 1985, 11, 1617–1629. [Google Scholar] [CrossRef]
- Jallon, J.M.; David, J.R. Variations in cuticular hydrocarbons among the eight species of the Drosophila melanogaster subgroup. Evolution 1987, 4, 294–302. [Google Scholar] [CrossRef]
- Sharma, M.D.; Mitchell, C.; Hunt, J.; Tregenza, T.; Hosken, D.J. The genetics of cuticular hydrocarbon profiles in the fruit fly Drosophila simulans. J. Hered. 2012, 103, 230–239. [Google Scholar] [CrossRef]
- Jallon, J.M.; Wicker-Thomas, C. Insect Pheromone Biochemistry and Molecular Biology. Part 1. Biochemistry and Molecular Biology of Pheromone Production; Blomquist, G.J., Vogt, R.G., Eds.; Elsevier Academic Press: London, UK, 2003; pp. 253–281. [Google Scholar]
- Scott, D.; Richmond, R.C. A genetic analysis of male-predominant pheromones in Drosophila melanogaster. Genetics 1988, 119, 639–646. [Google Scholar]
- Sureau, G.; Ferveur, J.F. Co-adaptation of pheromone production and behavioural responses in Drosophila melanogaster males. Genet. Res. 1999, 74, 129–137. [Google Scholar] [CrossRef]
- Grillet, M.; Dartevelle, L.; Ferveur, J.F. A Drosophila male pheromone affects female sexual receptivity. Proc. Biol. Sci. 2006, 273, 315–323. [Google Scholar] [CrossRef]
- Cobb, M.; Jallon, J.M. Pheromones, mate recognition and courtship stimulation in the Drosophila melanogaster species sub-group. Anim. Behav. 1990, 39, 1058–1069. [Google Scholar] [CrossRef]
- Coyne, J.A.; Crittenden, A.P.; Mah, K. Genetics of a pheromonal difference contributing to reproductive isolation in Drosophila. Science 1994, 265, 1461–1464. [Google Scholar]
- Coyne, J.A. Genetics of differences in pheromonal hydrocarbons between Drosophila melanogaster and D. simulans. Genetics 1996, 143, 353–364. [Google Scholar]
- Coyne, J.A.; Charlesworth, B. Genetics of a pheromonal difference affecting sexual isolation between Drosophila mauritiana and D. sechellia. Genetics 1997, 145, 1015–1030. [Google Scholar]
- Blows, M.W.; Allan, R.A. Levels of mate recognition within and between two Drosophila species and their hybrids. Am. Nat. 1998, 152, 826–837. [Google Scholar]
- Higgie, M.; Chenoweth, S.; Blows, M.W. Natural selection and the reinforcement of mate recognition. Science 2000, 290, 519–521. [Google Scholar] [CrossRef]
- Coyne, J.A.; Oyama, R. Localization of pheromonal sexual dimorphism in Drosophila melanogaster and its effect on sexual isolation. Proc. Natl. Acad. Sci. USA 1995, 92, 9505–9509. [Google Scholar] [CrossRef]
- Legendre, A.; Miao, X.X.; da Lage, J.L.; Wicker-Thomas, C. Evolution of a desaturase involved in female pheromonal cuticular hydrocarbon biosynthesis and courtship behavior in Drosophila. Insect Biochem. Mol. Biol. 2008, 38, 244–255. [Google Scholar] [CrossRef]
- Billeter, J.C.; Atallah, J.; Krupp, J.J.; Millar, J.G.; Levine, J.D. Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 2009, 461, 987–991. [Google Scholar] [CrossRef]
- Savarit, F.; Sureau, G.; Cobb, M.; Ferveur, J.F. Genetic elimination of known pheromones reveals the fundamental chemical bases of mating and isolation in Drosophila. Proc. Natl. Acad. Sci. USA 1999, 96, 9015–9020. [Google Scholar] [CrossRef]
- Coyne, J.A.; Kim, S.Y.; Chang, A.S.; Lachaise, D.; Elwyn, S. Sexual isolation between two sibling species with overlapping ranges: Drosophila santomea and Drosophila yakuba. Evolution 2002, 56, 2424–2434. [Google Scholar] [CrossRef]
- Mas, F.; Jallon, J.M. Sexual isolation and cuticular hydrocarbon differences between Drosophila santomea and Drosophila yakuba. J. Chem. Ecol. 2005, 31, 2747–2752. [Google Scholar] [CrossRef]
- Jallon, J.M.; Hotta, Y. Genetic and behavioral studies of female sex appeal in Drosophila. Behav. Genet. 1979, 9, 257–275. [Google Scholar] [CrossRef]
- McRobert, S.P.; Tompkins, L. Courtship of young males is ubiquitous in Drosophila melanogaster. Behav. Genet. 1979, 13, 517–523. [Google Scholar] [CrossRef]
- Péchiné, J.M.; Antony, C.; Jallon, J.M. Precise characterization of cuticular compounds in young Drosophila by mass spectrometry. J. Chem. Ecol. 1988, 14, 1071–1085. [Google Scholar] [CrossRef]
- Vias, L.J.; Napolitano, L.M.; Tompkins, L. Identification of stimuli that mediate experience-dependent modification of homosexual courtship in Drosophila melanogaster. Behav. Genet. 1993, 23, 91–97. [Google Scholar] [CrossRef]
- Dukas, R. Causes and consequences of male-male courtship in fruit flies. Anim. Behav. 2010, 80, 913–919. [Google Scholar] [CrossRef]
- Ishii, K.; Hirai, Y.; Katagiri, C.; Kimura, M.T. Sexual isolation and cuticular hydrocarbons in Drosophila elegans. Heredity 2002, 87, 392–399. [Google Scholar]
- Veltsos, P.; Wicker-Thomas, C.; Butlin, R.K.; Hoikkala, A.; Ritchie, M.G. Sexual selection on song and cuticular hydrocarbons in two distinct populations of Drosophila montana. Ecol. Evol. 2012, 2, 80–94. [Google Scholar] [CrossRef] [Green Version]
- Jallon, J.M.; Péchiné, J.M. Une autre race de chimique Drosophila melanogaster en Afrique. C. R. Acad. Sci. Paris 1989, 309, 1551–1556. [Google Scholar]
- Wu, C.I.; Hollocher, H.; Begun, D.J.; Aquadro, C.F.; Xu, Y.; Wu, M.L. Sexual isolation in Drosophila melanogaster: A possible case of incipient speciation. Proc. Natl. Acad. Sci. USA 1995, 92, 2519–2523. [Google Scholar] [CrossRef]
- Ferveur, J.F.; Cobb, M.; Boukella, H.; Jallon, J.M. World-wide variation in Drosophila melanogaster sex pheromone: Behavioural effects, genetic bases and potential evolutionary consequences. Genetica 1996, 97, 73–80. [Google Scholar] [CrossRef]
- Hollocher, H.; Ting, C.T.; Wu, M.L.; Wu, C.I. Incipient speciation by sexual isolation in Drosophila melanogaster: Extensive genetic divergence without reinforcement. Genetics 1997, 147, 1191–1201. [Google Scholar]
- Dallerac, R.; Labeur, C.; Jallon, J.M.; Knipple, D.C.; Roelofs, W.L.; Wicker-Thomas, C. A delta 9 desaturase gene with a different substrate specificity is responsible for the cuticular diene hydrocarbon polymorphism in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2000, 97, 9449–9454. [Google Scholar] [CrossRef]
- Yukilevich, R.; True, J.R. African morphology, behavior and phermones underlie incipient sexual isolation between us and Caribbean Drosophila melanogaster. Evolution 2008, 62, 2807–2828. [Google Scholar] [CrossRef]
- Takahashi, A.; Tsaur, S.C.; Coyne, J.A.; Wu, C.I. The nucleotide changes governing cuticular hydrocarbon variation and their evolution in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 3920–3925. [Google Scholar] [CrossRef]
- Coyne, J.A.; Elwyn, S. Does the desaturase-2 locus in Drosophila melanogaster cause adaptation and sexual isolation? Evol. Int. J. Org. Evol. 2006, 60, 279–291. [Google Scholar] [CrossRef]
- Grillet, M.; Everaerts, C.; Houot, B.; Ritchie, M.G.; Cobb, M.; Ferveur, J.F. Incipient speciation in Drosophila melanogaster involves chemical signals. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Bontonou, G.; Denis, B.; Wicker-Thomas, C. Interaction between temperature and male pheromone in sexual isolation in Drosophila melanogaster. J. Evol. Biol. 2013, 26, 2008–2020. [Google Scholar] [CrossRef]
- Rouault, J.; Capy, P.; Jallon, J.M. Variations of male cuticular hydrocarbons with geoclimatic variables: An adaptative mechanism in Drosophila melanogaster? Genetica 2001, 110, 117–130. [Google Scholar] [CrossRef]
- Rouault, J.D.; Marican, C.; Wicker-Thomas, C.; Jallon, J.M. Relations between cuticular hydrocarbon (HC) polymorphism, resistance against desiccation and breeding temperature; a model for HC evolution in D. melanogaster and D. simulans. Genetica 2004, 120, 195–212. [Google Scholar] [CrossRef]
- Bontonou, G.; Denis, B.; Wicker-Thomas, C. Male pheromone polymorphism and reproductive isolation in populations of Drosophila simulans. Ecol. Evol. 2012, 2, 2527–2536. [Google Scholar] [CrossRef]
- Ting, C.T.; Takahashi, A.; Wu, C.I. Incipient speciation by sexual isolation in Drosophila: Concurrent evolution at multiple loci. Proc. Natl. Acad. Sci. USA 2001, 98, 6709–6713. [Google Scholar]
- Stennett, M.D.; Etges, W.J. Premating isolation is determined by larval rearing substrates in cactophilic Drosophila mojavensis. III. Epicuticular hydrocarbon variation is determined by use of different host plants in Drosophila mojavensis and Drosophila arizonae. J. Chem. Ecol. 1997, 23, 2803–2824. [Google Scholar] [CrossRef]
- Havens, J.A.; Etges, W.J. Premating isolation is determined by larval rearing substrates in cactophilic Drosophila mojavensis. IX. Host plant and population specific epicuticular hydrocarbon expression influences mate choice and sexual selection. J. Evol. Biol. 2013, 26, 562–576. [Google Scholar] [CrossRef]
- Kent, C.; Azanchi, R.; Smith, B.; Formosa, A.; Levine, J.D. Social context influences chemical communication in D. melanogaster males. Curr. Biol. 2008, 18, 1384–1389. [Google Scholar] [CrossRef]
- Liu, W.; Liang, X.; Gong, J.; Yang, Z.; Zhang, Y.H.; Zhang, J.X.; Rao, Y. Social regulation of aggression by pheromonal activation of Or65a olfactory neurons in Drosophila. Nat. Neurosci. 2011, 14, 896–902. [Google Scholar] [CrossRef]
- Frentiu, F.D.; Chenoweth, S.F. Clines in cuticular hydrocarbons in two Drosophila species with independent population histories. Evolution 2010, 64, 1784–1794. [Google Scholar] [CrossRef]
- Kwan, L.; Rundle, H.D. Adaptation to desiccation fails to generate pre- and postmating isolation in replicate Drosophila melanogaster laboratory populations. Evolution 2010, 64, 710–723. [Google Scholar] [CrossRef]
- Foley, B.R.; Telonis-Scott, M. Quantitative genetic analysis suggests causal association between cuticular hydrocarbon composition and desiccation survival in Drosophila melanogaster. Heredity 2011, 106, 68–77. [Google Scholar] [CrossRef]
- Gibbs, A.G. Waterproofing properties of cuticular lipids. Am. Zool. 1998, 38, 471–482. [Google Scholar]
- Chenoweth, S.F.; Blows, M.W. Contrasting mutual sexual selection on homologous signal traits in Drosophila serrata. Am. Nat. 2005, 165, 291–289. [Google Scholar]
- Hine, E.; Lachish, S.; Higgie, M.; Blows, M.W. Positive genetic correlation between female preference and offspring fitness. Proc. Biol. Sci. 2002, 269, 2215–2219. [Google Scholar] [CrossRef]
- Higgie, M.; Blows, M.W. The evolution of reproductive character displacement conflicts with how sexual selection operates within a species. Evolution 2008, 62, 1192–1203. [Google Scholar] [CrossRef]
- Higgie, M.; Blows, M.W. Are traits that experience reinforcement also under sexual selection? Am. Nat. 2007, 170, 409–420. [Google Scholar] [CrossRef]
- Rundle, H.D.; Chenoweth, S.F.; Doughty, P.; Blows, M.W. Divergent selection and the evolution of signal traits and mating preferences. PLoS Biol. 2005, 3, e368. [Google Scholar] [CrossRef] [Green Version]
- Hine, E.; McGuigan, K.; Blows, M.W. Natural selection stops the evolution of male attractiveness. Proc. Natl. Acad. Sci. USA 2011, 108, 3659–3664. [Google Scholar] [CrossRef]
- Greenberg, A.J.; Moran, J.R.; Coyne, J.A.; Wu, C.I. Ecological adaptation during incipient speciation revealed by precise gene replacement. Science 2003, 302, 1754–1757. [Google Scholar]
- Rundle, H.D. Divergent environments and population bottlenecks fail to generate premating isolation in Drosophila pseudoobscura. Evolution 2003, 57, 2557–2565. [Google Scholar] [CrossRef]
- Chung, H.; Loehlin, D.W.; Dufour, H.D.; Vaccarro, K.; Millar, J.G.; Carroll, S.B. A single gene affects both ecological divergence and mate choice in Drosophila. Science 2014, 343, 1148–1151. [Google Scholar] [CrossRef]
- Bartelt, R.J.; Schaner, A.M.; Jackson, L.L. Cis-vaccenyl acetate as an aggregation pheromone in Drosophila melanogaster. J. Chem. Ecol. 1985, 11, 1747–1756. [Google Scholar] [CrossRef]
- Brieger, G.; Butterworth, F.M. Drosophila melanogaster: Identity of male lipid in reproductive system. Science 1970, 167, 1262. [Google Scholar]
- Butterworth, F. Lipids of Drosophila: A newly detected lipid in the male. Science 1969, 163, 1356–1357. [Google Scholar]
- Jallon, J.M.; Antony, C.; Benamar, O. Un anti-aphrodisiaque produit par les mâles de Drosophila melanogaster et transféré aux femelles lors de la copulation. C. R. Acad. Sci. Paris 1981, 292, 1147–1149. [Google Scholar]
- Yew, J.Y.; Dreisewerd, K.; Luftmann, H.; Muthing, J.; Pohlentz, G.; Kravitz, E.A. A new male sex pheromone and novel cuticular cues for chemical communication in Drosophila. Curr. Biol. 2009, 19, 1245–1254. [Google Scholar] [CrossRef]
- Scott, D.; Shields, A.; Straker, M.; Dalrymple, H.; Dhillon, P.K.; Harbinder, S. Variation in the male pheromones and mating success of wild caught Drosophila melanogaster. PLoS One 2011, 6, e23645. [Google Scholar]
- Chertemps, T.; Duportets, L.; Labeur, C.; Wicker-Thomas, C. A new elongase selectively expressed in Drosophila male reproductive system. Biochem. Biophys. Res. Commun. 2005, 333, 1066–1072. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Obin, M.S.; Zawistowski, S.; Sheehan, K.B.; Richmond, R.C. A reevaluation of the role of cis-vaccenylacetate, cis-vaccenol and esterase 6 in the regulation of mated female sexual attractiveness in Drosophila melanogaster. J. Insect Physiol. 1986, 32, 681–686. [Google Scholar] [CrossRef]
- Bairati, A.; Perotti, M.E. Occurrence of a compact plug in the genital duct of Drosophila females after mating. Drosop. Inf. Serv. 1970, 45, 67–68. [Google Scholar]
- Mane, S.D.; Tompkins, L.; Richmond, R.C. Male esterase 6 catalyzes the synthesis of a sex pheromone in Drosophila melanogaster females. Science 1983, 222, 419–421. [Google Scholar]
- Zawistowski, Z.; Richmond, R.C. Inhibition of courtship and mating of Drosophila melanogaster by the male-produced lipid, cis-vaccenyl acetate. J. Insect Physiol. 1986, 1986, 189–192. [Google Scholar] [CrossRef]
- Ejima, A.; Smith, B.P.; Lucas, C.; van der Goes van Naters, W.; Miller, C.J.; Carlson, J.R.; Levine, J.D.; Griffith, L.C. Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate. Curr. Biol. 2007, 17, 599–605. [Google Scholar] [CrossRef]
- Scott, D.; Richmond, R.C. Evidence against an antiaphrodisiac role for cis-vaccenyl acetate in Drosophila melanogaster. J. Insect Physiol. 1987, 33, 363–369. [Google Scholar] [CrossRef]
- Spieth, T.H. Courtship behavior in Drosophila. Ann. Rev. Entomol. 1974, 19, 385–405. [Google Scholar] [CrossRef]
- Schaner, A.M.; Bartelt, R.J.; Jackson, L.L. (Z)-11-octadecenyl acetate, an aggregation pheromone in Drosophila simulans. J. Chem. Ecol. 1987, 13, 1777–1786. [Google Scholar] [CrossRef]
- Hedlund, K.; Bartelt, R.J.; Dicke, M.; Vet, L.E.M. Aggregation pheromones of Drosophila immigrans, D. phaleratan and D. obscura. J. Chem. Ecol. 1996, 22, 1835–1844. [Google Scholar] [CrossRef]
- Jaenike, K.; Bartelt, R.J.; Huberty, A.F.; Thibault, S.; Libler, J.S. Aggregations in mycophagous Drosophila (Diptera: Drosophilidae): Candidate pheromones and field responses. Ann. Entomol. Soc. Am. 1992, 85, 696–704. [Google Scholar]
- Symonds, M.R.; Wertheim, B. The mode of evolution of aggregation pheromones in Drosophila species. J. Evol. Biol. 2005, 18, 1253–1263. [Google Scholar] [CrossRef]
- Wertheim, B.; van Baalen, E.J.; Dicke, M.; Vet, L.E. Pheromone-mediated aggregation in nonsocial arthropods: An evolutionary ecological perspective. Annu. Rev. Entomol. 2005, 50, 321–346. [Google Scholar] [CrossRef]
- Wertheim, B.; Allemand, R.; Vet, L.E.M.; Dicke, M. Effects of aggregation pheromone on individual behavior and food web interactions: A field study on Drosophila. Ecol. Entomol. 2006, 31, 216–226. [Google Scholar] [CrossRef]
- Wang, L.; Anderson, D.J. Identification of an aggression-promoting pheromone and its receptor neurons in Drosophila. Nature 2010, 463, 227–232. [Google Scholar] [CrossRef]
- Yew, J.Y.; Dreisenwerd, K.; de Oliveira, C.C.; Etges, W.J. Male-specific transfer and fine scale spatial differences of newly identified cuticular hydrocarbons and triacylglycerides in a Drosophila species pair. PLoS One 2011, 6, e16898. [Google Scholar]
- Chin, J.S.R.; Ellis, S.R.; Pham, H.T.; Blanksby, S.J.; Mori, K.; Koh, Q.L.; Etges, W.J.; Yew, J.Y. Sex-specific triacylglycerides are widely conserved in Drosophila and mediate mating behavior. eLife 2014, 3, e01751. [Google Scholar] [CrossRef]
- Rota-Stabelli, O.; Blaxter, M.; Anfora, G. Drosophila suzukii. Curr. Biol. 2013, 23, R8–R9. [Google Scholar] [CrossRef]
- Levine, J.D.; Funes, P.; Dowse, H.B.; Hall, J.C. Resetting the circadian clock by social experience in Drosophila melanogaster. Science 2002, 298, 2010–2012. [Google Scholar] [CrossRef]
- Fujii, S.; Krishnan, P.; Hardin, P.; Amrein, H. Nocturnal male sex drive in Drosophila. Curr. Biol. 2007, 17, 244–251. [Google Scholar]
- Krupp, J.J.; Kent, C.; Billeter, J.C.; Azanchi, R.; So, A.K.; Schonfeld, J.A.; Smith, B.P.; Lucas, C.; Levine, J.D. Social experience modifies pheromone expression and mating behavior in male Drosophila melanogaster. Curr. Biol. 2008, 18, 1373–1383. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bontonou, G.; Wicker-Thomas, C. Sexual Communication in the Drosophila Genus. Insects 2014, 5, 439-458. https://doi.org/10.3390/insects5020439
Bontonou G, Wicker-Thomas C. Sexual Communication in the Drosophila Genus. Insects. 2014; 5(2):439-458. https://doi.org/10.3390/insects5020439
Chicago/Turabian StyleBontonou, Gwénaëlle, and Claude Wicker-Thomas. 2014. "Sexual Communication in the Drosophila Genus" Insects 5, no. 2: 439-458. https://doi.org/10.3390/insects5020439
APA StyleBontonou, G., & Wicker-Thomas, C. (2014). Sexual Communication in the Drosophila Genus. Insects, 5(2), 439-458. https://doi.org/10.3390/insects5020439