
Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Sampling and Analyses
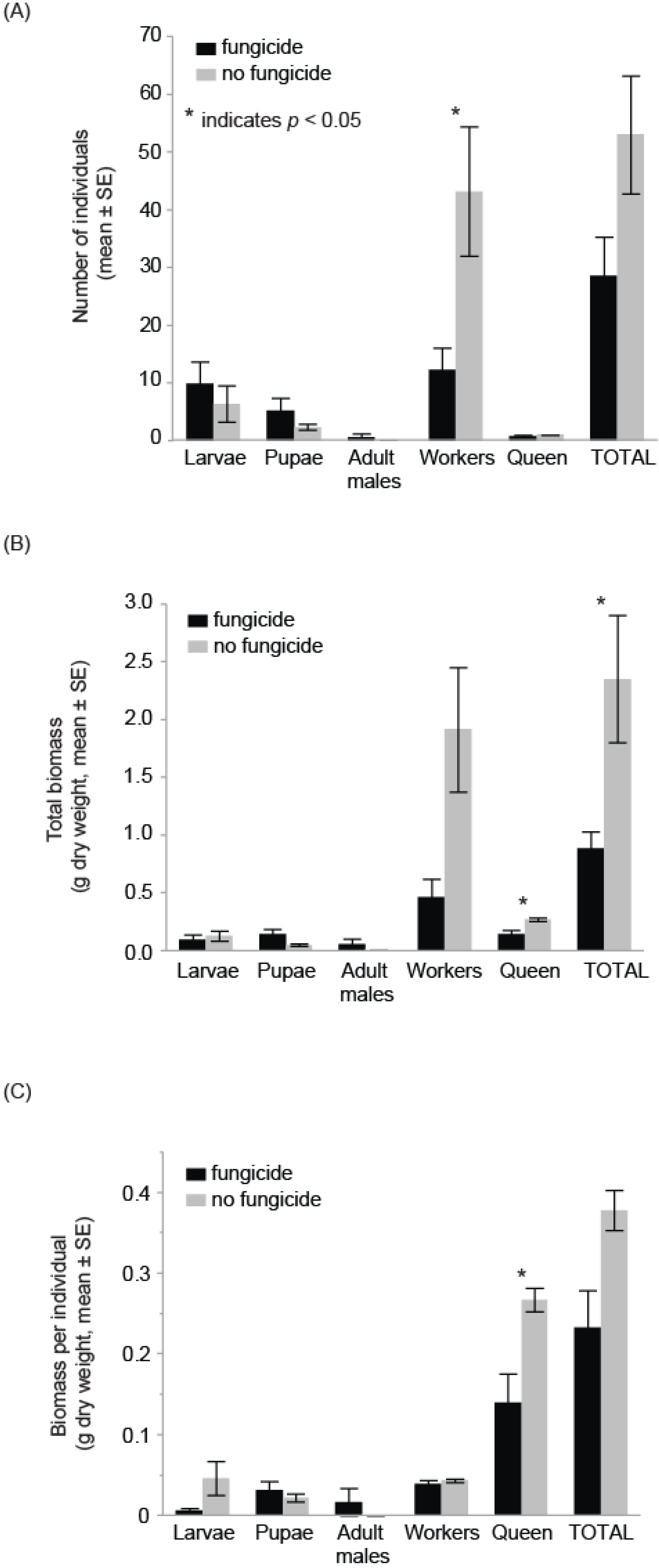
3. Results
4. Discussion

{kind=link}
{kind=link}
| Y-variable | Shapiro-Wilk † | F-test, 2-sided ‡ | ANOVA | Wilcoxon Rank Sum | |
|---|---|---|---|---|---|
| Number | Larvae | W = 0.92, p = 0.37 | F4,4 = 1.5, p = 0.71 | F1,9 = 0.47, p = 0.51 | |
| Pupae | W = 0.96, p = 0.84 | F4,4 = 17.8, p = 0.02 * | Z = −0.74, p = 0.46 | ||
| Workers | W = 0.97, p = 0.92 | F4,4 = 8.5, p = 0.06 | F1,9 = 6.8, p = 0.03 * | ||
| Adult males | W = 0.62, p = 0.0001 *** | insufficient data | Z = −0.8, p = 0.42 | ||
| Total | W = 0.96, p = 0.80 | F4,4 = 2.3, p = 0.43 | F1,9 = 4.0, p = 0.08 | ||
| Biomass | Larvae | W = 0.95, p = 0.67 | F4,4 = 1.1, p = 0.93 | F1,9 = 0.27, p = 0.62 | |
| Pupae | W = 0.89, p = 0.17 | F4,4 = 13.1, p = 0.03 * | Z = −1.5, p = 0.14 | ||
| Workers | W = 0.94, p = 0.58 | F4,4 = 12.1, p = 0.03 * | Z = 1.7, p = 0.09 | ||
| Adult males | W = 0.62, p = 0.0001 *** | insufficient data | Z = −0.8, p = 0.42 | ||
| √(total) | W = 0.96, p = 0.79 | F4,4 = 5.6, p = 0.12 | F1,9 = 8.3, p = 0.02* | ||
| Biomass per individual | Larva | W = 0.85, p = 0.08 | F4,4 = 101.5, p = 0.003 ** | Z = −1.1, p = 0.62 | |
| Pupa | W = 0.94, p = 0.63 | F3,4 = 3.9, p = 0.22 | F1,8 = 2.2, p = 0.18 | ||
| Workers | W = 0.96, p = 0.78 | F4,4 = 5.1, p = 0.14 | F1,9 = 0.80, p = 0.40 | ||
| Adult male | insufficient data | insufficient data | insufficient data | ||
| Queen | W = 0.81, p = 0.02 * | F4,4 = 6.2, p = 0.11 | Z = 2.5, p = 0.01* | ||

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Cane, J.H.; Schiffhauer, D. Dose-response relationships between pollination and fruiting refine pollinator comparisons for cranberry (Vaccinium macrocarpon [Ericaceae]). Am. J. Bot. 2003, 90, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Vicens, N.; Bosch, J. Pollinating efficacy of Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae, Apidae) on “red Delicious” apple. Environ. Entomol. 2000, 29, 235–240. [Google Scholar] [CrossRef]
- Bosch, J.; Kemp, W.P.; Trostle, G.E. Bee population returns and cherry yields in an orchard pollinated with Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2006, 99, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Pitts-Singer, T.L.; Cane, J.H. The Alfalfa Leafcutting Bee, Megachile rotundata: The World’s Most Intensively Managed Solitary Bee. Annu. Rev. Entomol. 2011, 56, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.D.; Evans, J.D.; Pettis, J. Colony losses, managed colony population decline, and Colony Collapse Disorder in the United States. J. Apic. Res. 2010, 49, 134–136. [Google Scholar] [CrossRef]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. In Annual Review of Entomology; Annual Reviews: Palo Alto, CA, USA, 2008; Volume 53, pp. 191–208. [Google Scholar]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyoergyi, H.; Woyciechowski, M.; Vila, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and honey bee toxicity—USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; vanEngelsdorp, D.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Schmehl, D.R.; Mullin, C.A.; Frazier, J.L. Four common pesticides, their mixtures and a formulation solvent in the hive environment have high oral toxicity to honey bee larvae. PLoS ONE 2014, 9, e77547. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Acaricide, fungicide and drug interactions in honey bees (Apis mellifera). PLoS ONE 2013, 8, e54092. [Google Scholar] [CrossRef] [PubMed]
- Morandin, L.A.; Winston, M.L.; Franklin, M.T.; Abbott, V.A. Lethal and sub-lethal effects of spinosad on bumble bees (Bombus impatiens Cresson). Pest Manag. Sci. 2005, 61, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Beeing there: The search for pesticides’ effect on declining bee colonies moves to the fields. Available online: http://www.scientificamerican.com/article/beeing-there-the-search-for-pesticides-effect-on-declining-bee-colonies-moves-to-the-fields/ (accessed on 17 March 2015).
- Sterk, G.; Heuts, F.; Merck, N.; Bock, J. Sensitivity of non-target arthropods and beneficial fungal species to chemical and biological plant protection products: Results of laboratory and semi-field trials. In Proceedings of the International Symposium on Biolgocial Control of Arthropods, Pucón, Chile, 4–8 March 2013.
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, W.F.; de Meyer, L.; Guedes, R.N.C.; Smagghe, G. Lethal and sublethal effects of azadirachtin on the bumblebee Bombus terrestris (Hymenoptera: Apidae). Ecotoxicology 2012, 24, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Ladurner, E.; Bosch, J.; Kemp, W.P.; Maini, S. Assassing delayed and acute toxicity of five formulated fungicides to Osmia lignaria Say and Apis mellifera. Apidologie 2005, 36, 449–460. [Google Scholar] [CrossRef]
- Sprayberry, J.D.H.; Ritter, K.A.; Riffell, J.A. The Effect of olfactory exposure to non-insecticidal agrochemicals on bumblebee foraging behavior. PLoS ONE 2013, 8, e76273. [Google Scholar] [CrossRef] [PubMed]
- vanEngelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J., Jr.; Pettis, J.S. “Entombed Pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Mussen, E.C.; Lopez, J.E.; Peng, C.Y.S. Effects of selected fungicides on growth and development of larval honey bees, Apis mellifera L. (Hymenoptera: Apidae). Environ. Entomol. 2004, 33, 1151–1154. [Google Scholar] [CrossRef]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; vanEngelsdorp, D. Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, S.S.; Kremen, C. Wild bee species increase tomato production and respond differently to surrounding land use in Northern California. Biol. Conserv. 2006, 133, 81–87. [Google Scholar] [CrossRef]
- Scott-Dupree, C.D.; Conroy, L.; Harris, C.R. Impact of currently used or potentially useful insecticides for canola agroecosystems on Bombus impatiens (Hymenoptera: Apidae), Megachile rotundata (Hymentoptera: Megachilidae), and Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2009, 102, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Malone, L.A.; Scott-Dupree, C.D.; Todd, J.H.; Ramankutty, P. No sub-lethal toxicity to bumblebees, Bombus terrestris, exposed to Bt-corn pollen, captan and novaluron. N. Z. J. Crop Hortic. Sci. 2007, 35, 435–439. [Google Scholar] [CrossRef]
- Survilene, E.; Raudonis, L.; Jankauskiene, J. Investigation of pesticides effect on pollination of bumblebees in greenhouse tomatoes. Sci. Works Lith. Inst. Hortic. Lith. Univ. Agric. 2009, 28, 235–241. [Google Scholar]
- Elston, C.; Thompson, H.M.; Walters, K.F.A. Sub-lethal effects of thiamethoxam, a neonicotinoid pesticide, and propiconazole, a DMI fungicide, on colony initiation in bumblebee (Bombus terrestris) micro-colonies. Apidologie 2013, 44, 563–574. [Google Scholar] [CrossRef]
- Gianessi, L.; Reigner, N. The value of fungicides in U.S. crop production. Available online: http://croplife.intraspin.com/Pesticides/paper.asp?id=442 (accessed on 5 March 2015).
- Van Scoy, A.R.; Tjeerdema, R.S. Environmental fate and Toxicology of chlorothalonil. Rev. Environ. Contam. Toxicol. 2014, 232, 89–105. [Google Scholar] [PubMed]
- Koch, P.L.; Stier, J.C.; Senseman, S.A.; Sobek, S.; Kerns, J.P. Modification of a commercially-available ELISA kit to determine chlorothalonil and iprodione concentration on golf course turfgrass. Crop Prot. 2013, 54, 35–42. [Google Scholar] [CrossRef]
- De Ibarra, N.H.; Philippides, A.; Riabinina, O.; Collett, T.S. Preferred viewing directions of bumblebees (Bombus terrestris L.) when learning and approaching their nest site. J. Exp. Biol. 2009, 212, 3193–3204. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. JMP Pro 10, SAS Institute Inc.: Cary, NC, USA, 2007.
- Everich, R.; Schiller, C.; Whitehead, J.; Beavers, M.; Barrett, K. Effects of Captan on Apis mellifera brood development under field conditions in California almond orchards. J. Econ. Entomol. 2009, 102, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, B. Bumblebee Economics; Harvard University Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Pomeroy, N.; Plowright, R.C. The relation between worker numbers and the production of males and queens in the bumble bee Bombus perplexus. Can. J. Zool. 1982, 60, 954–957. [Google Scholar] [CrossRef]
- Mattila, H.R.; Rios, D.; Walker-Sperling, V.E.; Roeselers, G.; Newton, I.L.G. Characterization of the active microbiotas associated with honey bees reveals healthier and broader communities when colonies are genetically diverse. PLoS ONE 2012, 7, e32962. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, M. Identification and roles of non-pathogenic microflora associated with honey bees1. FEMS Microbiol. Lett. 1997, 155, 1–10. [Google Scholar]
- Kariluoto, S.; Aittamaa, M.; Korhola, M.; Salovaara, H.; Vahteristo, L.; Piironen, V. Effects of yeasts and bacteria on the levels of folates in rye sourdoughs. Int. J. Food Microbiol. 2006, 106, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.A.; Jajack, A.J.; Rosselot, A.E.; Smith, T.J.; Yerke, M.C.; Sammataro, D. Fungicide contamination reduces beneficial fungi in bee bread based on an area-wide field study in honey bee, Apis mellifera, colonies. J. Toxicol. Environ. Health A 2013, 76, 587–600. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernauer, O.M.; Gaines-Day, H.R.; Steffan, S.A. Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure. Insects 2015, 6, 478-488. https://doi.org/10.3390/insects6020478
Bernauer OM, Gaines-Day HR, Steffan SA. Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure. Insects. 2015; 6(2):478-488. https://doi.org/10.3390/insects6020478
Chicago/Turabian StyleBernauer, Olivia M., Hannah R. Gaines-Day, and Shawn A. Steffan. 2015. "Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure" Insects 6, no. 2: 478-488. https://doi.org/10.3390/insects6020478
APA StyleBernauer, O. M., Gaines-Day, H. R., & Steffan, S. A. (2015). Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure. Insects, 6(2), 478-488. https://doi.org/10.3390/insects6020478