Growth of RWA on Selected Wheat Genotypes
The RWA populations did not significantly differ in their survivorship on KRWA9 and Kwale (
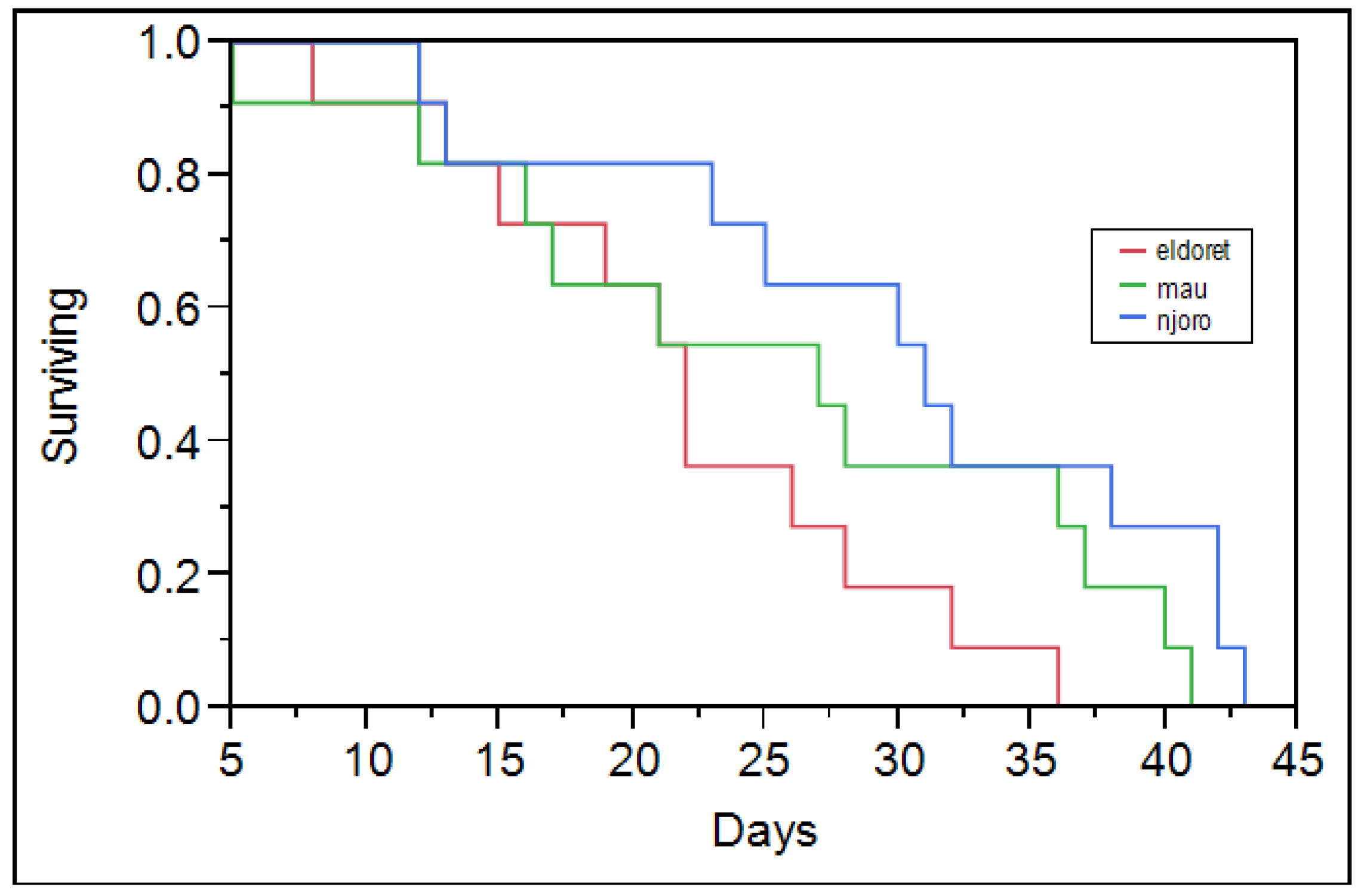
Table 2). Eldoret and Mau Narok populations had a characteristic survivorship curve with constant mortality across all ages (
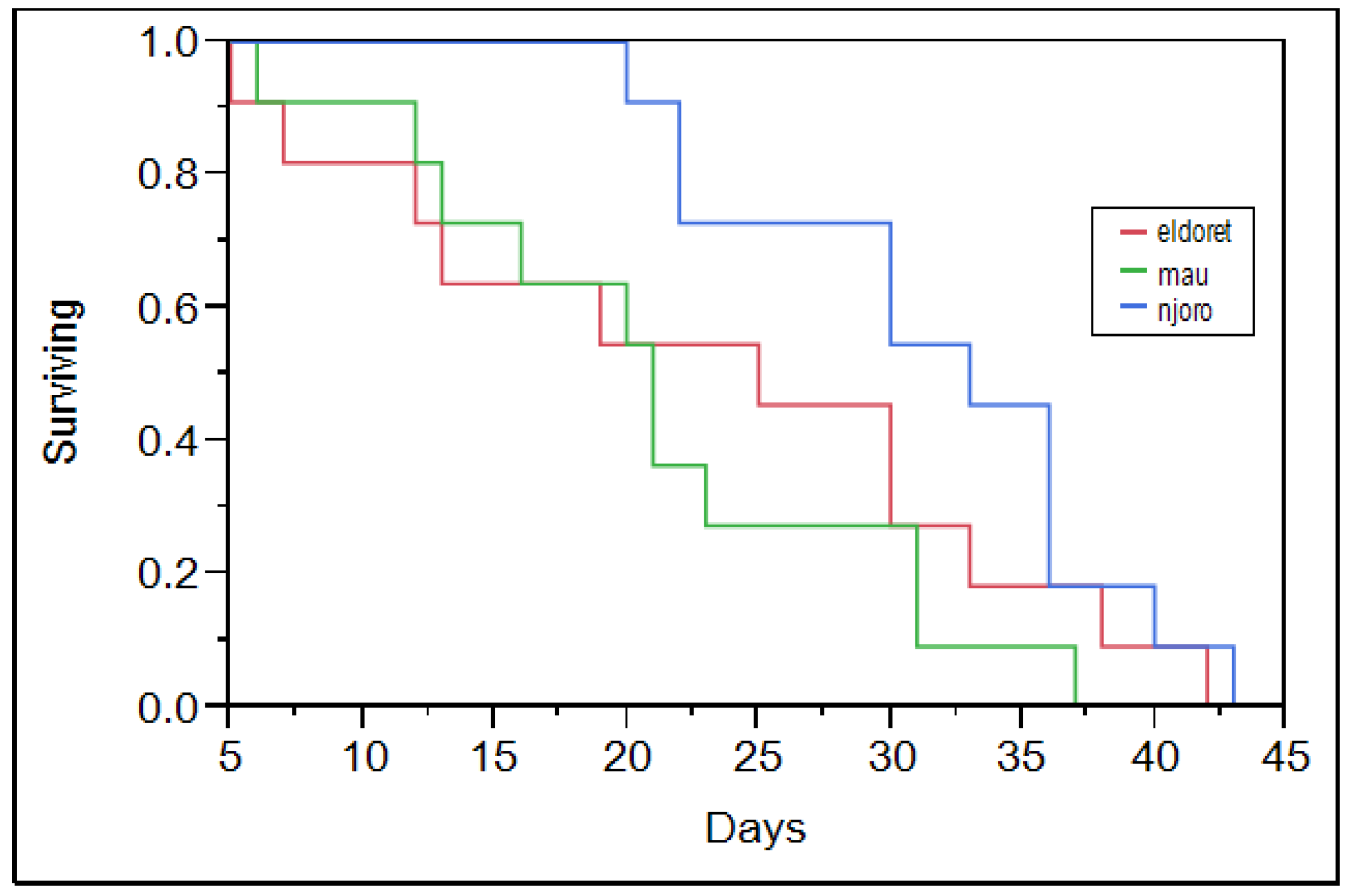
Figure 1 and
Figure 2). The Njoro population however had high mortality towards its maximum lifetime. Maling’a
et al [
27] noted a similar mortality trend for RWA population collected from the Njoro population. The Njoro population had low mortality among its young compared to the Eldoret and Mau Narok populations (
Figure 1 and
Figure 2). The difference is probably because the different populations are different. The prevailing greenhouse temperature was common to all of the RWA populations that were studied. The Njoro population, which has lived under similar conditions for a longer duration, was more adapted compared to other populations that were acclimatized in the greenhouse for three months prior to the start of the study. Njoro has a lower altitude compared to both Mau Narok and Eldoret (
Table 2). Michels and Behle [
28] in their study of RWA populations in Ethiopia found that the mortality of RWA increased with the increase in temperature. This could have caused the increased mortality of the Mau Narok and Eldoret populations since they were collected from regions with lower temperatures and acclimatized in the greenhouse for three months.
The aphid populations did not differ in their development time (the time from birth to first reproduction). Wheat genotypes however had a significant effect on the development time of populations of RWA (
Table 3). Primary and secondary metabolites found in plant phloem exudates influence aphid development and reproduction [
29,
30]. Resistant plants contain higher levels of antibiotic secondary metabolites, such as hydroxamic acids, that reduce insect attack [
31]. Kwale did not have RWA resistance and was more suitable for aphid development compared to KRWA9. Aphid populations took a significantly shorter time to develop on Kwale compared to KRWA9.
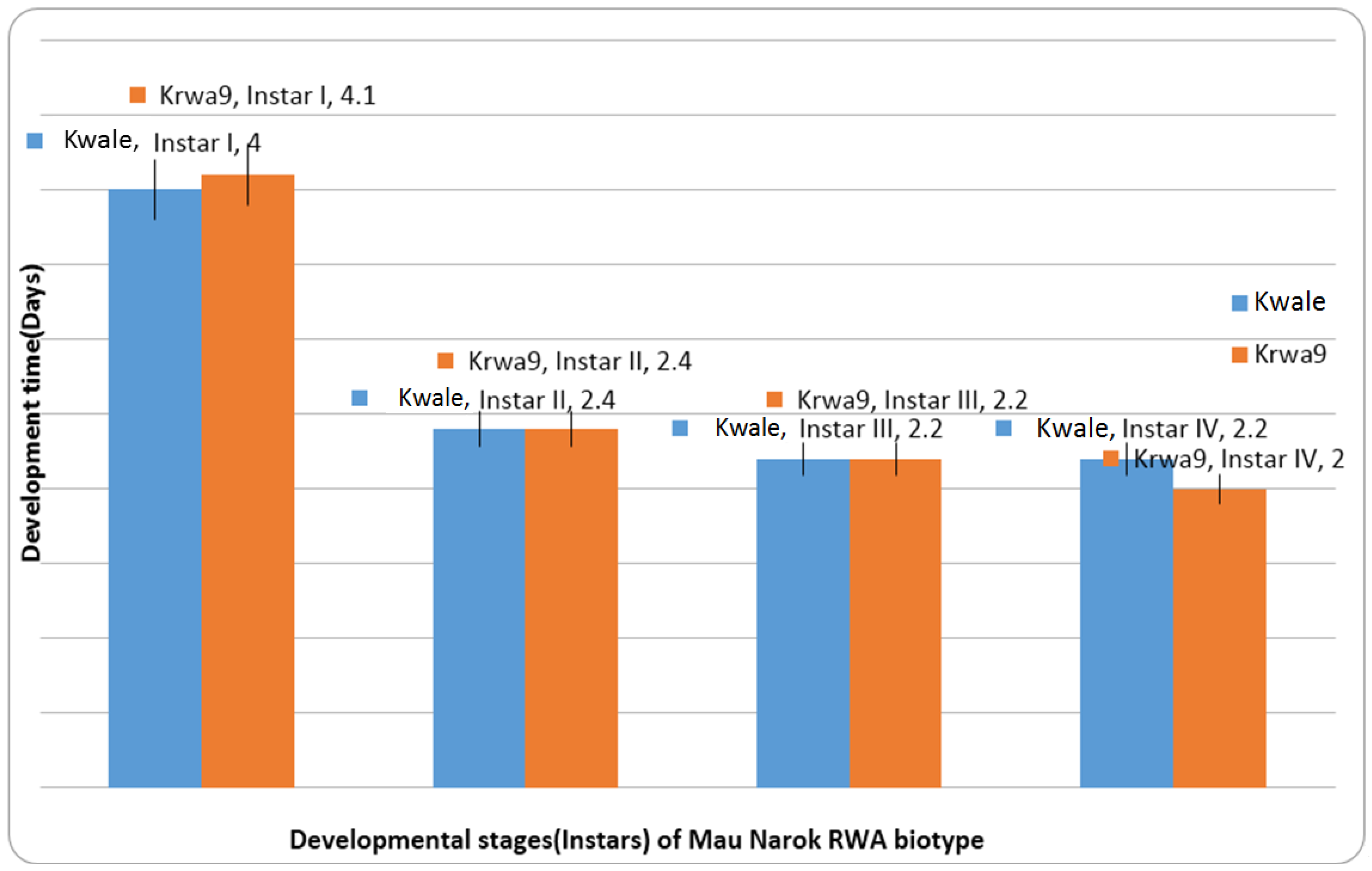
There was significant host-aphid population interaction on the development time of the first instar of RWAs (
Table 4). The development time of the first instar of the Mau Narok and Njoro populations was significantly different from the Eldoret population. The development time of the second instar of the Mau Narok RWA population was significantly longer compared to the Eldoret and Njoro RWA populations, which had similar development time on Kwale. Overall, the development differed significantly among RWA populations on Kwale. The Eldoret RWA population had the shortest development time on Kwale compared to Mau Narok and Njoro populations, whose development time was similar on Kwale. There were no differences in the development time of RWA populations on KRWA9 (
Table 5). Second and third instars of the Mau Narok population were observed to be indifferent to the effect of variety on development time (
Figure 3).This result indicates that the Mau Narok population is virulent, since the resistance of KRWA9 does not affect the development time of its instars (
Table 4). There was no host by aphid population interaction on the development time of the third and fourth instars of all tested RWA populations (
Table 4). The results observed are consistent with the findings by Aalbersberg
et al. [
32], Kazemi
et al. [
33] and Maling’a
et al. [
19].
Wheat genotypes did not significantly affect the reproduction time and lifespan of aphids. Host genotype, however, significantly affected the M
d (fecundity in a time equivalent to the aphid development time, M
d) and total aphid fecundity. Genotype Kwale was the most suitable host, as shown by the high number of progeny produced on it (
Table 5). Qing-Nian
et al. [
34] similarly noted that resistant wheat genotypes significantly decreased the population growth of the grain aphid
Sitobion avenae and therefore noted that the oviposition behavior of herbivorous insects responds to host quality and availability [
20,
35,
36]. All populations had significantly higher total fecundity and M
d (Fecundity at a time equivalent to the development time) on Kwale, a susceptible host compared to resistant genotype KRWA9. This indicates that KRWA9 has antibiosis as the mode of resistance, since it affects aphid biology. The high increase in population during a time equivalent to the development time has been cited as critical to determining the individual contribution to the population of an aphid species, because aphids contribute almost 90% of the progeny to population during this period [
25].
There were significant differences in aphid reproductive time, total aphid lifespan, M
d (fecundity in a time equivalent to the aphid development time) and total aphid fecundity between aphid populations (
Table 6). The Njoro population produced a significantly higher number of progeny and had the longest reproduction time compared to the Mau Narok and Eldoret populations. The Njoro population had a higher reproductive longevity and fecundity than the Eldoret and Mau Narok populations (
Table 6). The Njoro and Eldoret populations had a higher number of progeny at a duration equivalent to the development time (M
d), compared to Mau Narok. The differences in total aphid fecundity can be attributed to differences in aphid populations, wheat genotypes and the longevity of the reproductive time.
There was no host by population interaction on reproduction time, total lifespan, daily fecundity and fecundity of a time equivalent to the development time of RWA. However, there was a host by population interaction on total fecundity. The Eldoret and Njoro populations had the highest fecundity on Kwale, while the Njoro and Mau Narok populations had the highest progeny on KRWA9. The Eldoret population, however, had the lowest progeny on KRWA9, indicating that this genotype may be effective in managing the population buildup of the Eldoret population of RWA (
Table 6). Diehl and Bush [
14] defined conspecific, sympatric populations, which differ in some biological traits, as biotypes. Longevity of reproductive time, aphid lifespan and total aphid fecundity of aphid populations clearly show the Njoro population to be a distinct biotype of RWA in Kenya.
All aphid populations had a positive intrinsic rate of population increase, indicating their ability to build up populations. There was a significant effect of wheat genotype and aphid population on the intrinsic rate of the natural increase of the populations; the aphid populations were, however, not different from each other when the intrinsic rate of natural increase was compared and when cohort generation time was compared (
Table 6 and
Table 7). The interaction between host genotype and aphid population was not significant on the intrinsic rate of the natural increase of RWA populations and the cohort generation time of RWA. Genotype Kwale had the biggest effect on the intrinsic rate of natural increase compared to KRWA9 and was the best host for increasing the aphid population of RWA. Maling’a [
19] reported that the aphid populations developed faster and higher populations on susceptible wheat genotypes without
Dn genes as compared to resistant wheat genotypes, like KRWA9, that contained
Dn genes and the progenies. This shows that resistant wheat genotypes that contain
Dn genes can be used to manage RWA populations in Kenya.
Pearson’s correlation coefficients for aphid development time, aphid reproductive time (reproduction longevity), aphid lifespan, M
d, total fecundity, intrinsic rate of natural increase and cohort generation time are presented in
Table 8. A significant negative relationship was observed between aphid development time and M
d (
r = 0.299,
p < 0.05), indicating that populations with a shorter development time had higher progenies during the initial reproduction period. Development time was also negatively correlated with total fecundity and the intrinsic rate of natural increase. This means that the aphid population with the shortest development time has a higher intrinsic rate of natural increase and total fecundity. Cohort generation time was negatively correlated with M
d, total fecundity and the intrinsic rate of natural increase, meaning that these parameters cannot be used to predict the cohort generation time of RWA.
Reproductive time had a significant positive correlation with aphid lifespan, Md, total fecundity and the intrinsic rate of natural increase. Aphid lifespan was positively correlated with Md, total fecundity and the intrinsic rate of natural increase, whereas Md was strongly correlated with total fecundity and the intrinsic rate of natural increase. There was no correlation between development time, reproductive time, aphid lifespan and cohort generation time.






{kind=link}
{kind=link}
{kind=link}