
Seasonal Effects on the Population, Morphology and Reproductive Behavior of Narnia femorata (Hemiptera: Coreidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Cactus Patch Sampling Study
2.3. Insect Sampling Study
2.4. Readiness to Mate and Beak Length
2.5. Statistical Analyses
3. Results
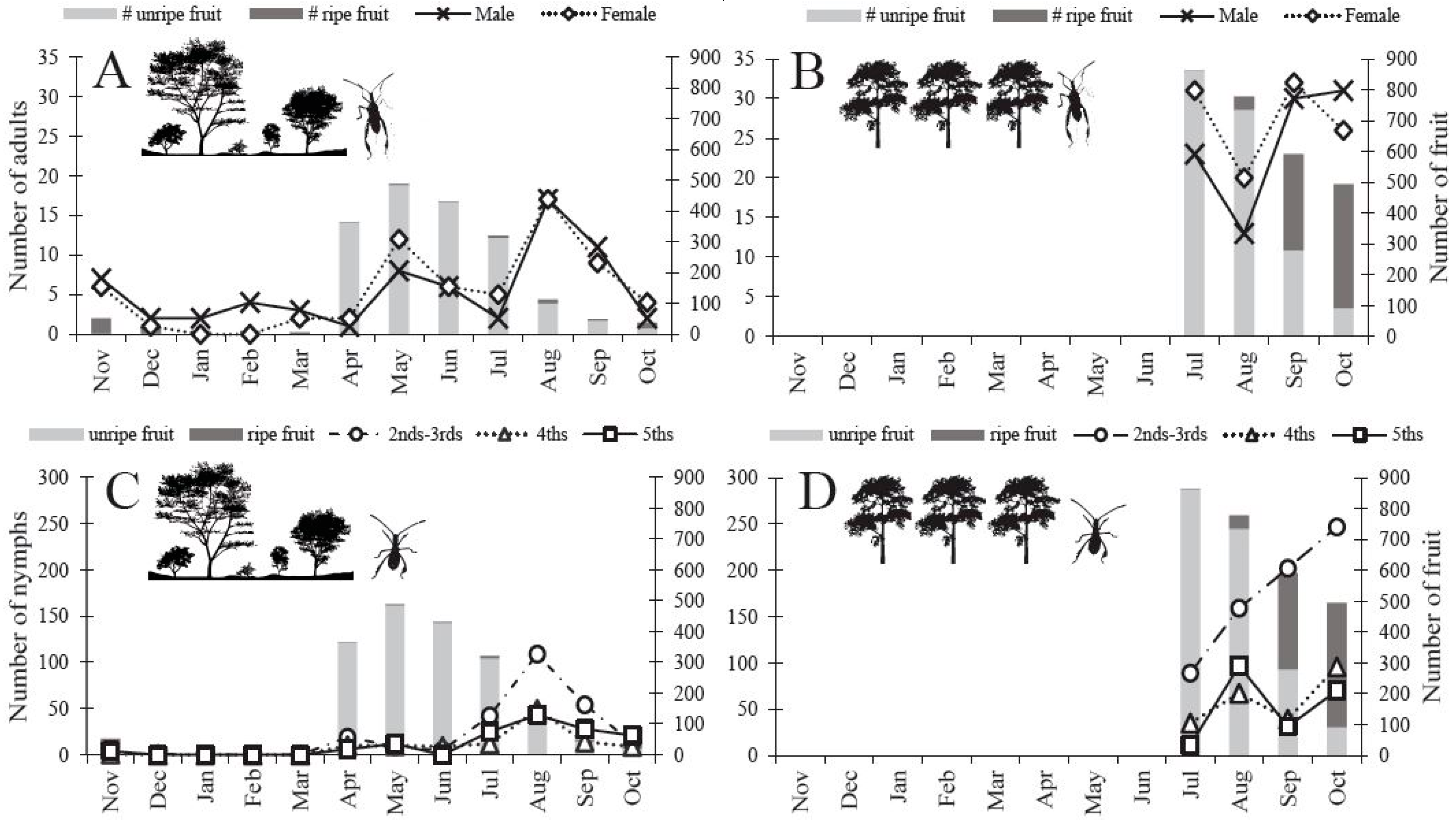
3.1. Fruit Number and Phenology of O. Mesacantha Ssp. Lata
3.2. Abundance of N. femorata
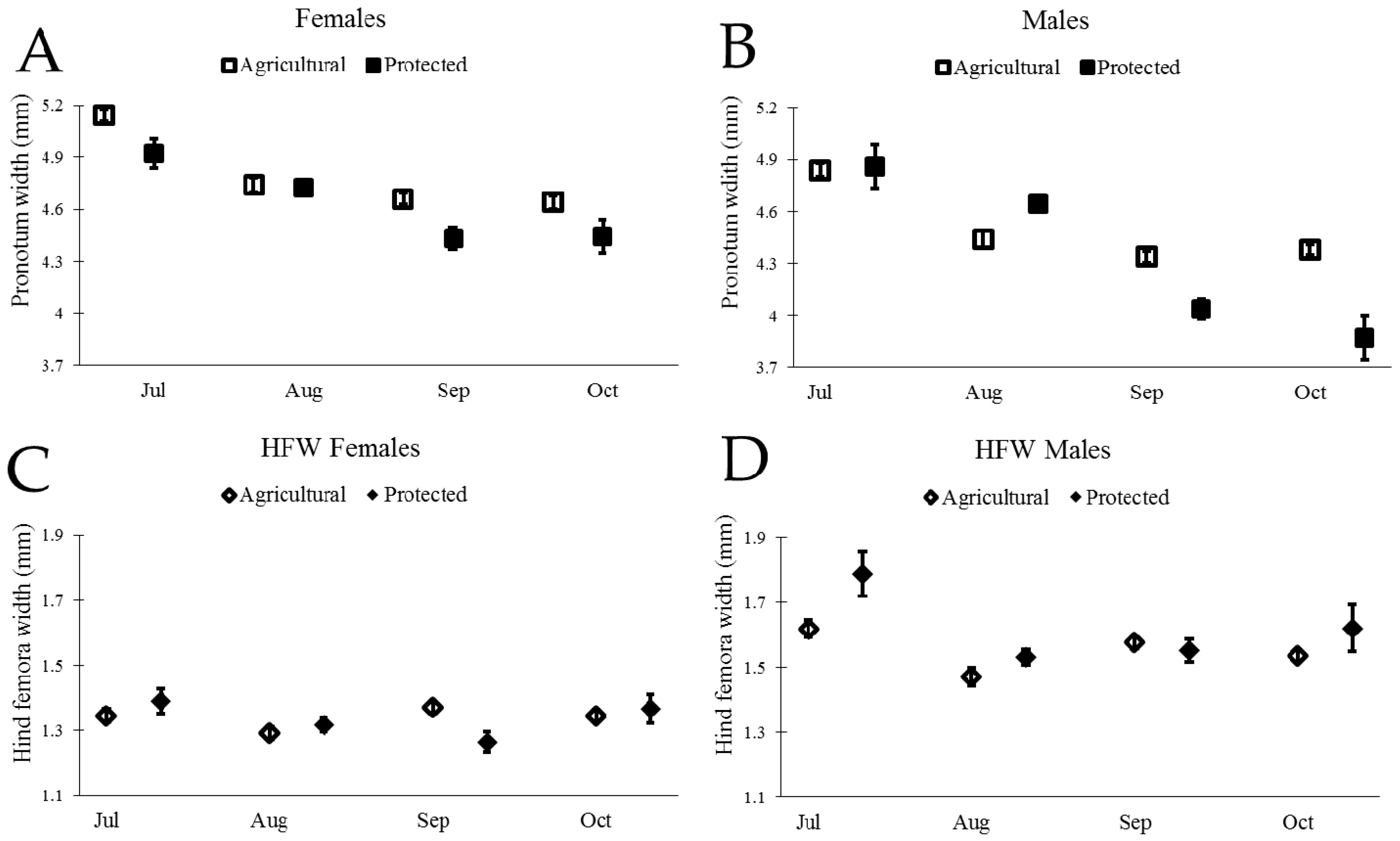
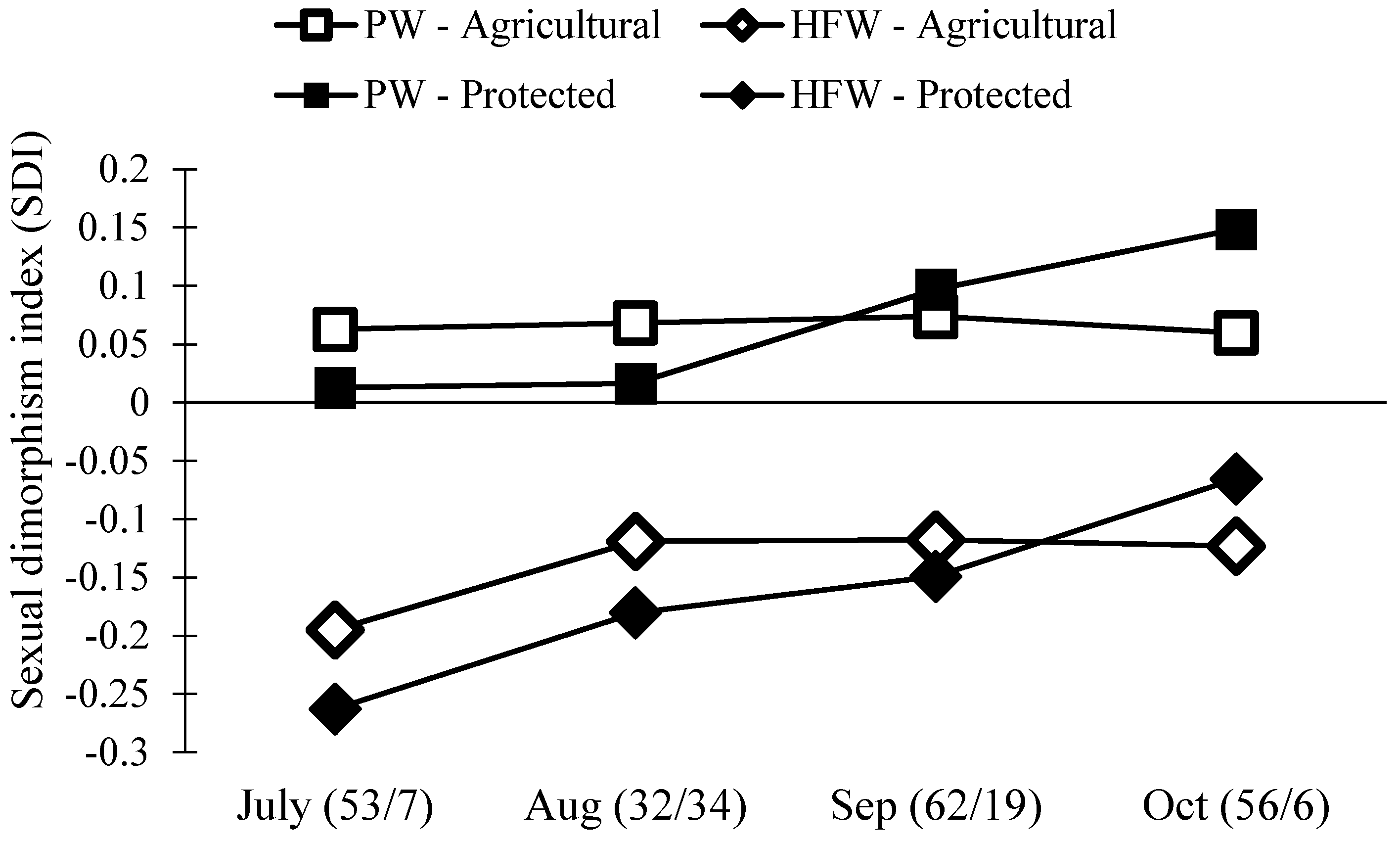
3.3. Size and Sexual Dimorphism of N. Femorata
3.4. Beak Length in N. Femorata
3.5. Female and Male Readiness to Mate in N. Femorata
4. Discussion
4.1. Size and Sexual Dimorphism
4.2. Beak Length
4.3. Readiness to Mate
4.4. Abundance and Natural History
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Natuhara, Y. The influences of food and photoperiod on flight activity and reproduction of the bean bug, Riptortus clavatus Thungberg (Heteroptera: Coreidae). Appl. Entomol. Zool. 1983, 18, 392–400. [Google Scholar]
- Wojcik, V.A.; Frankie, G.W.; Thorp, R.W.; Hernandez, J.L. Seasonality in bees and their floral resource plants at a constructed urban bee habitat in Berkeley, California. J. Kansas Entomol. Soc. 2008, 81, 15–28. [Google Scholar] [CrossRef]
- Ribeiro, D.B.; Freitas, A.V.L. Large-sized insects show stronger seasonality than small-sized ones: A case study of fruit-feeding butterflies. Biol. J. Linn. Soc. 2011, 104, 820–827. [Google Scholar] [CrossRef]
- Miller, C.W.; Fletcher, R.J.; Gillespie, S.R. Conspecific and heterospecific cues override resource quality to influence offspring production. PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schaik, C.P.; Terborgh, J.W.; Wright, S.J. The phenology of tropical forests: Adaptive significance and consequences of primary consumers. Annu. Rev. Ecol. Syst. 1993, 24, 353–377. [Google Scholar] [CrossRef]
- Numata, H.; Hidaka, T. Photoperiodic control of adult diapause in the bean bug, Riptortus clavatus Thunberg (Heteroptera: Coreidae) IV.: Food and post-diapause development. Appl. Entomol. Zool. 1984, 19, 443–447. [Google Scholar]
- McLain, D.K.; Burnette, L.B.; Deeds, D.A. Within season variation in the intensity of sexual selection on body size in the bug Margus obscurator (Hemiptera: Coreidae). Ethol. Ecol Evol. 1993, 5, 75–86. [Google Scholar] [CrossRef]
- Sasson, D.A.; Munoz, P.R.; Gezan, S.A.; Miller, C.W. Resource quality affects weapon and testis size and the ability of these traits to respond to selection in the leaf-footed cactus bug, Narnia femorata. Ecol. Evol. 2016, 6, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- Dingle, H. Migration strategies of insects. Science 1972, 175, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Beck, S.D.; Reese, J.C. Insect-plant interactions: Nutrition and metabolism. In Biochemical Interaction between Plants and Insects; Wallace, W. J., Mansell, R.L., Eds.; Springer: New York, NY, USA, 1976; pp. 41–92. [Google Scholar]
- Dixon, A.F.G.; Wellings, P.W. Seasonality and reproduction in aphids. Int. J. Invertebr. Reprod. 1982, 5, 83–89. [Google Scholar] [CrossRef]
- Janzen, D.H. Ecological characterization of a costa rican dry forest caterpillar fauna. Biotropica 1988, 20, 120–135. [Google Scholar] [CrossRef]
- Hunter, M.D.; McNeil, J.N. Host-plant quality influences diapause and voltinism in a polyphagous insect herbivore. Ecology 1997, 78, 977–986. [Google Scholar] [CrossRef]
- Mopper, S. Adaptive genetic: Structure in phytophagous insect populations. Trends Ecol. Evol. 1996, 11, 235–238. [Google Scholar] [CrossRef]
- Wellings, P.W.; Leather, S.R.; Dixon, A.F.G. Seasonal-variation in reproductive potential—A programmed feature of aphid life-cycles. J. Anim. Ecol. 1980, 49, 975–985. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.; Endler, J.A. The impact of predation on life-history evolution in Trinidadian guppies (Poecilia reticulata). Evolution 1982, 36, 160–177. [Google Scholar] [CrossRef]
- Schluter, D. Ecology and the origin of species. Trends Ecol. Evol. 2001, 16, 372–380. [Google Scholar] [CrossRef]
- Grant, B.R.; Grant, P.R. Lack of premating isolation at the base of a phylogenetic tree. Am. Nat. 2002, 160, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Stål, C. Narnia femorata. Stettin. Entomol. Zeit. 1862, 23, 296. [Google Scholar]
- Brailovsky, H. Distribucion de las especies de Narnia Stål (Coreidae-Coreinae-Anisoscelini) y descripcion de una nueva especie. Revista Sociedad Mexicana Historia Natural 1975, 36, 169–176. (In Spanish) [Google Scholar]
- Baranowski, R.M.; Slater, J.A. Coreidae of Florida (Hemiptera: Heteroptera). In Arthropods of Florida and Neighboring Land Areas; Florida Department of Agriculture and Consumer Services, Division of Plant Industry: Gainesville, FL, USA, 1986. [Google Scholar]
- Brailovsky, H.; Barrera, E.; Mayorga, C.; Ortega-León, G. Estudios ninfales de los coreidos del valle de tehuacán, puebla. (Hemiptera-Heteroptera) I. Chelinidea saffilesi, C. Tabulata y Narnia femorata. Anales Instituto Biología Serie Zoologia 1994, 65, 241–264. (In Spanish) [Google Scholar]
- Palomares-Perez, M.; Galeana-de la Cruz, M.; Carrillo-Fonseca, C.; Sancen-Plaza, A. Report Narnia femorata Stål (Hemiptera: Coreidae) on Opuntia ficus-indica l. (Miller) in Milpa Alta, Mexico City, Mexico. Southwest. Entomol. 2012, 37, 415–418. (In Corsican) [Google Scholar] [CrossRef]
- Mann, J. Cactus-feeding insects and mites. Bull. US Natn. Mus. 1969, 256, 1–158. [Google Scholar] [CrossRef]
- Majure, L.C. Typifications and a nomenclatural change in some eastern north american Opuntia (Cactaceae). Phytoneuron 2014, 106, 1–2. [Google Scholar]
- Gibson, E.H.; Holdridge, A. The genus Narnia Stål, and a key to the genera of Anisoscelini a. and s. (Coreidae: Heteroptera). Psyche 1918, 25, 1–4. [Google Scholar] [CrossRef]
- Mitchell, P.L. Leaf-footed bugs (Coreidae). In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 337–403. [Google Scholar]
- Cirino, L.A.; Entomology & Nematology Department, University of Florida, Gainesville, FL, USA. Personal observation. 2015.
- Miller, T.E.X.; Tyre, A.J.; Louda, S.M. Plant reproductive allocation predicts herbivore dynamics across spatial and temporal scales. Am. Nat. 2006, 168, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Vessels, H.K.; Bundy, C.S.; McPherson, J.E. Life history and laboratory rearing of Narnia femorata (Hemiptera: Heteroptera: Coreidae) with descriptions of immature stages. Ann. Entomol. Soc. Am. 2013, 106, 575–585. [Google Scholar] [CrossRef]
- Hunter, W.D.; Pratt, F.C.; Mitchell, J.D. Narnia pallidicornis stål. In The Principal Cactus Insects Of The United States; Department of Agriculture, Bureau of Entomology: Washington, DC, USA, 1912; pp. 32–33. [Google Scholar]
- Allen, P.E.; Miller, C.W. Complex diet-by-temperature interactions on sexually selected traits and sexual dimorphism. 2016; In preparation. [Google Scholar]
- Proctor, D.S.; Moore, A.J.; Miller, C.W. The form of sexual selection arising from male-male competition depends on the presence of females in the social environment. J. Evolution. Biol. 2012, 25, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, S.R.; Tudor, M.S.; Moore, A.J.; Miller, C.W. Sexual selection is influenced by both developmental and adult environments. Evolution 2014, 68, 3421–3432. [Google Scholar] [CrossRef] [PubMed]
- Nageon-De Lestang, F.; Miller, C. Effects of diet on development and survivorship of Narnia femorata nymphs (Hemiptera: Coreidae). Fla. Entomol. 2009, 92, 511–512. [Google Scholar] [CrossRef]
- Gonzalez-Espinosa, M.; Quintana-Ascencio, P. Seed predation and dispersal in a dominant desert plant: Opuntia, ants, birds and mammals. In Frugivores and Seed Dispersal; Estrada, A., Fleming, T.H., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 273–284. [Google Scholar]
- Janzen, D.H. Chihuahuan desert nopaleras–Defaunated big mammal vegetation. Annu. Rev. Ecol. Syst. 1986, 17, 595–636. [Google Scholar] [CrossRef]
- Hellgren, E.C. Prickly-pear cactus (Opuntia spp.) and its use by wildlife. In Proceedings of the 5th Annual Prickly Pear Council, Kingsville, TX, USA, 1994; pp. 87–93.
- Miller, C.W.; Entomology & Nematology Department, University of Florida, Gainesville, FL, USA. Personal observation. 2014.
- Miller, T.E.X. Bottom-up, top-down, and within-trophic level pressures on a cactus-feeding insect. Ecol. Entomol. 2008, 33, 261–268. [Google Scholar] [CrossRef]
- Miller, C.W.; Emlen, D.J. Dynamic effects of oviposition site on offspring sexually-selected traits and scaling relationships. Ecol. Entomol. 2010, 24, 375–390. [Google Scholar] [CrossRef]
- Addesso, K.R.; Short, K.A.; Moore, A.J.; Miller, C.W. Context-dependent female mate preferences in leaf-footed cactus bugs. Behaviour 2014, 151, 479–492. [Google Scholar] [CrossRef]
- Somjee, U.; Allen, P.E.; Miller, C.W. Different environments lead to a reversal in the expression of weapons and testes in the heliconia bug, Leptoscelis tricolor (Hemiptera: Coreidae). Biol. J. Linn. Soc. 2015, 115, 802–809. [Google Scholar] [CrossRef]
- Miller, C.W.; McDonald, G.C.; Moore, A.J. The tale of the shrinking weapon: Seasonal changes in nutrition affect weapon size and sexual dimorphism, but not contemporary evolution. J. Evolution. Biol. 2016, 29, 2266–2275. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.E.; University of Florida, Gainesville, FL, USA. Unpublished data. 2014.
- Barbera, G.; Carimi, F.; Inglese, P.; Panno, M. Physical, morphological and chemical-changes during fruit-development and ripening in 3 cultivars of prickly pear, Opuntia-ficus-indica (L.) miller. J. Hortic. Sci. 1992, 67, 307–312. [Google Scholar] [CrossRef]
- Miyatake, T. Seasonal abundance of the bamboo bug, Notobitus meleagris Fabricus (Heteroptera, Coreidae) in Okinawa Island. Appl. Entomol. Zool. 1994, 29, 601–603. [Google Scholar]
- Tuten, T.; Suwannee County High School, Live Oak, FL, USA. Personal communication, 2016.
- Schooley, R.L.; Wiens, J.A. Movements of cactus bugs: Patch transfers, matrix resistance, and edge permeability. Landscape Ecol. 2004, 19, 801–810. [Google Scholar] [CrossRef]
- Nolen, Z.J.; Allen, P.E.; Miller, C.W. Seasonal resource value and male size influence male aggressive interactions in the leaf footed cactus bug, Narnia femorata. Behav. Process. 2017, in press. [Google Scholar]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? In Annual Review of Entomology; Berenbaum, M.R., Ed.; Annual Reviews: Palo Alto, CA, USA, 2016; pp. 317–333. [Google Scholar]
- Miller, C.W.; University of Florida, Gainesville, FL, USA. Unpublished data. 2012.
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with imagej. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Hutcheson, G.D.; Sofroniou, N. The Multivariate Social Scientist: Introductory Statistics Using Generalized Linear Models; Sage Publications: London, UK, 1999. [Google Scholar]
- Lovich, J.E.; Gibbons, J.W. A review of techniques for quantifying sexual size dimorphism. Growth Develop. Aging 1992, 56, 269–281. [Google Scholar]
- Joseph, P.N.; Sasson, D.A.; Allen, P.E.; Somjee, U.; Miller, C.W. Adult nutrition, but not inbreeding, affects male primary sexual traits in the leaf-footed cactus bug Narnia femorata (Hemiptera: Coreidae). Ecol. Evol. 2016, 6, 4792–4799. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Calsbeek, R. Sex-specific selection and intraspecific variation in sexual size dimorphism. Evolution 2010, 64, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Stillwell, R.C.; Blanckenhorn, W.U.; Teder, T.; Davidowitz, G.; Fox, C.W. Sex differences in phenotypic plasticity affect variation in sexual size dimorphism in insects: From physiology to evolution. Annu. Rev. Entomol. 2010, 55, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Puniamoorthy, N.; Schafer, M.A.; Blanckenhorn, W.U. Sexual selection accounts for the geographic reversal of sexual size dimorphism in the dung fly, Sepsis punctum (Diptera: Sepsidae). Evolution 2012, 66, 2117–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manicom, C.; Alford, R.; Schoener, T.W.; Schwarzkopf, L. Mechanisms causing variation in sexual size dimorphism in three sympatric, congeneric lizards. Ecology 2014, 95, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.W.; Yuan, X.; Lei, C.L. Sexual size dimorphism decreases with temperature in a blowfly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae). Ecol. Entomol. 2011, 36, 111–115. [Google Scholar] [CrossRef]
- Miller, C.W.; Svensson, E.I. Sexual selection in complex environments. In Annual Review of Entomology; Berenbaum, M.R., Ed.; Annual Reviews: Palo Alto, CA, USA, 2014; pp. 427–445. [Google Scholar]
- Thompson, D.B. Genotype-environment interaction and the ontogeny of diet-induced phenotypic plasticity in size and shape of Melanoplus femurrubrum (Orthoptera: Acrididae). J. Evolution. Biol. 1999, 12, 38–48. [Google Scholar] [CrossRef]
- Carroll, S.P.; Boyd, C. Host race radiation in the soapberry bug: Natural-history with the history. Evolution 1992, 46, 1052–1069. [Google Scholar] [CrossRef]
- Relyea, R.A.; Auld, J.R. Predator- and competitor-induced plasticity: How changes in foraging morphology affect phenotypic trade-offs. Ecology 2005, 86, 1723–1729. [Google Scholar] [CrossRef]
- Gil, D.; Bulmer, E.; Celis, P.; Lopez-Rull, I. Adaptive developmental plasticity in growing nestlings: Sibling competition induces differential gape growth. P. Roy. Soc. B 2008, 275, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Stoler, A.B.; Relyea, R.A. Leaf litter quality induces morphological and developmental changes in larval amphibians. Ecology 2013, 94, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Greenway, E.V.; Doughtery, L.R.; Shucker, D.M. Mating failure. Curr. Biol. 2015, 25, R534–R536. [Google Scholar] [CrossRef] [PubMed]
- Milner, R.N.C.; Detto, T.; Jennions, M.D.; Backwell, P.R.Y. Experimental evidence for a seasonal shift in the strength of a female mating preference. Behav. Ecol. 2010, 21, 311–316. [Google Scholar] [CrossRef]
- Kahn, A.T.; Dolstra, T.; Jennions, M.D.; Backwell, P.R.Y. Strategic male courtship effort varies in concert with adaptive shifts in female mating preferences. Behav. Ecol. 2013, 24, 906–913. [Google Scholar] [CrossRef]
- Jennions, M.D.; Petrie, M. Variation in mate choice and mating preferences: A review of causes and consequences. Biol. Rev. 1997, 72, 283–327. [Google Scholar] [CrossRef] [PubMed]
- Sauby, K.E.; Marsico, T.D.; Ervin, G.N.; Baooks, C.P. The role of host identity in determining the distribution of the invasive moth Cactoblastis cactorum (Lepidoptera: Pyralidae) in Florida. Fla. Entomol. 2012, 95, 561–568. [Google Scholar] [CrossRef]
- Stiling, P. Death and decline of a rare cactus in Florida. Castanea 2010, 75, 190–197. [Google Scholar] [CrossRef]
- Jezorek, H.; Baker, A.J.; Stiling, P. Effects of Cactoblastis cactorum on the survival and growth of North American Opuntia. Biol. Invasions 2012, 14, 2355–2367. [Google Scholar] [CrossRef]
- Baker, A.J.; Stiling, P. Comparing the effects of the exotic cactus-feeding moth, Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae) and the native cactus-feeding moth, Melitara prodenialis (Walker) (Lepidoptera: Pyralidae) on two species of Florida Opuntia. Biol. Invasions 2009, 11, 619–624. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Male PW | Male HFW | Female PW | Female HFW | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | df | p | χ2 | df | p | χ2 | df | p | χ2 | df | p | |
| Site | 7.868 | 1 | 0.005 | 0.458 | 1 | 0.499 | 15.814 | 1 | <0.001 | 2.964 | 1 | 0.085 |
| Month | 123.178 | 3 | <0.001 | 67.216 | 3 | <0.001 | 81.086 | 3 | <0.001 | 27.591 | 3 | <0.001 |
| Site × Month | 40.652 | 3 | <0.001 | 25.322 | 3 | <0.001 | 6.213 | 3 | 0.102 | 15.238 | 3 | 0.002 |
| Source | Male HFW | Female HFW | ||||
|---|---|---|---|---|---|---|
| χ2 | df | p | χ2 | df | p | |
| PW | 56.663 | 1 | <0.001 | 25.239 | 1 | <0.001 |
| Site | 6.801 | 1 | 0.009 | 0.033 | 1 | 0.856 |
| Month | 28.132 | 3 | <0.001 | 7.950 | 3 | 0.047 |
| Site × Month | 7.408 | 3 | 0.060 | 12.828 | 3 | 0.005 |
| Source | Female BL | Male BL | ||||
|---|---|---|---|---|---|---|
| χ2 | df | p | χ2 | df | p | |
| Site | 0.945 | 1 | 0.331 | 0.002 | 1 | 0.964 |
| Month | 13.900 | 3 | 0.003 | 37.527 | 3 | <0.001 |
| Site × Month | 2.708 | 3 | 0.439 | 7.911 | 3 | 0.048 |
| Source | Female BL | Male BL | ||||
|---|---|---|---|---|---|---|
| χ2 | df | p | χ2 | df | P | |
| PW | 88.376 | 1 | <0.001 | 201.434 | 1 | <0.001 |
| Site | 0.190 | 1 | 0.663 | 0.745 | 1 | 0.388 |
| Month | 54.593 | 3 | <0.001 | 42.213 | 3 | <0.001 |
| Site × Month | 1.041 | 3 | 0.791 | 9.162 | 3 | 0.027 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirino, L.A.; Miller, C.W. Seasonal Effects on the Population, Morphology and Reproductive Behavior of Narnia femorata (Hemiptera: Coreidae). Insects 2017, 8, 13. https://doi.org/10.3390/insects8010013
Cirino LA, Miller CW. Seasonal Effects on the Population, Morphology and Reproductive Behavior of Narnia femorata (Hemiptera: Coreidae). Insects. 2017; 8(1):13. https://doi.org/10.3390/insects8010013
Chicago/Turabian StyleCirino, Lauren A., and Christine W. Miller. 2017. "Seasonal Effects on the Population, Morphology and Reproductive Behavior of Narnia femorata (Hemiptera: Coreidae)" Insects 8, no. 1: 13. https://doi.org/10.3390/insects8010013
APA StyleCirino, L. A., & Miller, C. W. (2017). Seasonal Effects on the Population, Morphology and Reproductive Behavior of Narnia femorata (Hemiptera: Coreidae). Insects, 8(1), 13. https://doi.org/10.3390/insects8010013