2. Materials and Methods
This work was performed in collaboration with ONC’s VENUS observatory, a cabled underwater observatory based at the University of Victoria on Vancouver Island, BC.
The research sites were at a depth of 170 m in the Strait of Georgia (latitude: 49°02.569′ N, longitude: 123°19.032′ W), part of the Salish Sea. The Strait of Georgia is considered to be Canada’s most important marine waterway and is one of the busiest seaports in North America [
24], shortly to become much busier. It is a large body of water between the mainland coast of BC, Vancouver Island and northern coastal Washington State.
The VENUS observatory includes shore stations which power communication and power hubs or nodes on the seafloor. Each node then connects to a science instrument interface module (SIIM) via fiber optic cable and, from there, to an array of oceanographic sensors, instruments, and cameras, which are continuously collecting physical, chemical, acoustic, and photographic data. The shore station also provides communication links between the instrument arrays and the University of Victoria, which houses a Network’s operation centre (NOC), which manages the functioning of the instruments and a data management and archive system (DMAS), which receives and processes the data which can then be accessed by researchers anywhere in the world via the Internet [
24].
In both spring (7 March 2014) and fall (20 September 2014), two paired, 16–23 kg freshly-killed pig carcasses were utilized. Simon Fraser University Animal Care Committee permission was obtained to purchase dead pigs, Animal Care Permission #1027CR-11. The pigs were humanely euthanized by a licensed butcher and were received immediately after death. They were not exsanguinated or frozen but were refrigerated on the research vessel prior to placement. They were refrigerated from leaving the dock until the vessel was at the research site and ready to deploy, (approximately eight to ten hours). The paired carcasses were placed onto a specially designed platform (designed by Chris Sundstom, ONC) attached to an instrument platform (designed by Paul Macoun, ONC) (
Figure 1).
One carcass was completely exposed, while the other was protected by widely-spaced bars (14–15 cm × 10–12 cm) to conserve at least one carcass in the event that sharks fed on the remains, but still allow arthropod access. This design had been proven successful in previous experiments at 300 m in the Strait of Georgia [
25]. The carcasses were positioned directly below a digital webcam (AXIS Q6034 720 p HD, Axis Communications, Lund, Sweden) with a lighting array (
Figure 1). The camera was continuously powered on, but lights were timed to come on every 15 min for a two minute period during which the camera was programmed to pan over both carcasses and the surrounding area. The camera could also be manually controlled when needed.
The entire pig platform was connected by fiber optic cable to the nearest VENUS instrument platform (VIP) and, from there to the shore station. A number of instruments were deployed with the carcasses to measure oceanic conditions. These included an oxygen optode (SeaBird 43 Oxygen (S/N 1807 (spring) and Aanderaa Optode 4175C (S/N 1795) 23204 (fall) and a CTD (SeaBird 16 Plus 6935 and SeaCat SBE 16plus 4998) which recorded temperature (°C), salinity (psu), pressure (decibar), conductivity (S/m), density, as sigma T (kg/m3), and speed of sound (m/s) at per minute intervals. In the spring experiments, the instruments were placed on the VIP approximately 70 m from the carcasses and in fall they were placed close to the carcasses on the instrument platform attached to the pig platform.
The pigs were tied onto the platform with hemp as, although no bloat or refloat would occur at this depth [
26], past experience has shown that animal activity can move a carcass out of camera view very rapidly [
12,
23]. The platforms were deployed from a research vessel to the seabed using the vessel’s crane, then positioned at the predefined site by a remotely-operated submersible vehicle (ROV). The ROV then connected the instruments and camera to the fibre optic cable, which connected to the VENUS array for remote monitoring. The day of submergence was deemed Day 0. Once deployed, there was no physical access to the carcasses. The platform and carcass remains were recovered approximately six months after deployment (Day 196 in spring and Day 191 in fall).
Videos and instrument data were downloaded from the VENUS website [
24] for analysis. At first all videos were viewed, then videos were viewed every hour. Over time, viewing was reduced to every two hours, then to every 6 h, then gradually reduced to daily, and eventually every five days. At any time, if a dramatic change occurred between viewings, the intermediate videos were observed.
4. Discussion
The carcasses deployed in the present study, at 170 m in the Strait of Georgia, were scavenged and skeletonized very differently from those deployed at 300 m in the same Strait. At 300 m, carcasses in both spring and fall were very rapidly covered in Lyssianassidae amphipods [
25]. The present spring carcasses also rapidly attracted amphipods, but in much fewer numbers and the fall carcasses did not attract appreciable numbers of amphipods until Day 15. At 300 m, amphipods rapidly increased in numbers on both carcasses and in each season until, by Day 1, all carcasses, as well as the surrounding cage area, were covered in amphipods many animals deep [
25]. Numbers continued to increase, with amphipods roosting on all surfaces and pooling onto the substrate surrounding the cage area for at least a metre, and in fall, not only covering the cage floor, edges, and ropes, but also completely covering the bars over the carcass briefly. The carcasses at 300 m began to visibly lose biomass by Day 2, with the fall carcasses skeletonized on Day 3, and spring carcasses on Day 4 [
25].
In the present experiments at 170 m, in spring, although amphipods did arrive within hours of placement, they were in much fewer numbers and never reached the density seen at 300 m. In fall, only very few amphipods were present in the first two weeks. The reduction in the numbers of amphipods in both spring and fall at 170 m, meant that other, larger, crustaceans were able to feed on the carcasses. At 300 m, although
P. platyceros were attracted to the spring carcasses immediately, they were rapidly excluded by the increasing numbers of amphipods, and did not return until the amphipods had receded and in fall, both
P. platyceros and
M. magister did not attend the carcasses until after the amphipods were gone. This meant that the 300 m carcasses were almost entirely skeletonized and disarticulated by the actions of the amphipods alone [
25]. In the present experiments, with fewer amphipods in spring and almost none in fall for the first two weeks, the larger crustaceans played a much greater role in skeletonizing the carcasses. In spring,
P. platyceros fed on the carcasses in large numbers, mostly avoiding areas with large masses of amphipods, so feeding primarily around the edges of the carcasses, shredding the skin and tissue. Almost no shrimp were present on the fall carcasses.
Although very few M. magister were present in spring (and those observed were small), in fall, the major scavenger was M. magister for the first two weeks. The large crabs were attracted immediately and ripped open the carcass and fed on all areas of the carcasses. They fed particularly on the exposed carcass which may have been a function of the cage bars which were designed to prevent shark’s from feeding. The bars were wide enough to allow large crabs to access the caged carcass and many did, but the bars may have proven a deterrent. The exposed carcass had consequently lost much of its soft tissue before amphipods began to arrive in larger numbers.
In spring at 170 m, and in both seasons at 300 m, amphipods covered the entire carcasses, but in both sets of experiments, they did not feed on the skin at first. When fish sloughed amphipods from the carcasses, undamaged skin, hair and even silt on the skin could be seen, indicating that the amphipods entered the carcasses through orifices and openings and fed internally and many of those observed were just roosting. This phenomenon is also observed by recovery divers in which human bodies in dive suits are entirely covered in roosting amphipods and, when recovered, amphipods are seen to emerge from orifices over many hours [
26]. When tissue is damaged, as it was by crabs in the fall experiments, the amphipods preferentially covered the areas of exposed muscle tissue and did not cover the areas of remaining skin at first. This was also seen in experiments in nearby Saanich Inlet at 100 m, where pig carcasses were first scavenged by sharks, then
M. magister and
P. platyceros [
12]. Although much fewer amphipods were observed and did not appear until Day 17, they covered only exposed flesh, feeding under the skin. The skin itself was consumed by numerous squat lobsters,
Munida quadrispina Benedict, 1902 [
12].
Amphipods have been shown to eat pig skin, as they were the only scavengers at 300 m, completely skeletonizing the carcass, eating all the soft tissue [
25], but it is clearly a less desired food substance. Previous experiments have shown that amphipods preferentially feed on muscle and fat and can differentiate between food types and texture [
27]. In experiments in which porpoise and dolphin carcasses were submerged at depths of 4000–4800 m in the Atlantic Ocean, the carcasses were skeletonized by a succession of amphipod species within six days, although the skin was not eaten [
28]. Observations of a decomposing Pilot whale (
Globiecephela mela Traill, 1809) and an intact Minke whale (
Balaenoptera acutorostrata Lacépède, 1804) deployed at 30 m and 125 m in a Swedish fjord showed that the bulk of the soft tissue was removed by Atlantic hagfish (
Myxine glutinosa L., 1758), sharks and other scavengers and amphipods (
O. obtusa) were not attracted until the skin was open and bones exposed [
29]. They were then present in dense populations, growing and reproducing on the remaining tissue from five weeks to four months after submersion, although images indicate that they were not present in the densities observed here in the Strait of Georgia at either 300 m or in spring at 170 m [
25,
29] and is closer to that seen in Saanich Inlet, when amphipods were only present after large wounds had been opened [
12]. In these experiments, and those at 300 m, the amphipods clearly avoided the skin until it was the last soft tissue remaining, when it was finally consumed.
Large openings in the skin had little impact on the time it took amphipods to skeletonize carcasses. In the earlier experiments at 300 m, Bluntnose sixgill sharks took large bites from the exposed carcass and the caged carcass did not display any damage, yet both exposed and caged carcasses were skeletonized at the same time [
25], showing that openings in the skin did not accelerate the amphipods’ ability to enter the carcasses. In these fall experiments, only a few amphipods were present from Day 8, with much larger numbers appearing on both carcasses on Day 16. By this time, much of the exposed carcass biomass had been removed by
M. magister and the caged carcass was still mostly intact. Despite these differences, when the amphipods arrived in large numbers, both carcasses were skeletonized within 24 h of each other, showing the exceptional feeding efficiency of the amphipods’ in comparison with that of the large crabs.
A disadvantage of remote monitoring is that it is not possible to collect samples of fauna, so the actual species of amphipods are not known, although earlier sampling from this area suggests the dominant species is
Orchomenella aff.
obtusa [
25]. However, it is possible that more than one species fed. Reports of amphipods in the
Orchomene group have suggested that some species are nocturnal [
30,
31,
32], however, in both of these experiments, and those at 300 m [
25], once present, the amphipods did not exhibit any diel activity, although at 170 m, the numbers of amphipods did appear to wax and wane, but this did not appear to follow a diel pattern and may have been more influenced by tides and currents.
As there were fewer amphipods in spring at 170 m and amphipods were delayed in fall, the scavenging patterns were quite different from those seen when only amphipods were feeding. When only amphipods were present, carcasses appeared to ‘deflate’, losing biomass and gradually collapsing under the amphipods, which then receded to reveal complete skeletonization. This occurred at 300 m [
25] and in the caged carcass in fall in these experiments. In contrast, with fewer amphipods present, the larger crustaceans picked at the skin and inner soft tissue with chelicerae, shredding the skin, with small pieces drifting in the water current. This presented a very different taphonomic pattern. Additionally, at 300 m, once the carcasses were skeletonized, the majority of the amphipods dispersed, with very low numbers returning briefly [
25]. However, in both spring and fall at 170 m, amphipods remained in very large numbers for some days after skeletonization.
The exclusion of the larger crustaceans by large masses of amphipods and the repeatedly observed avoidance of amphipods by
P. platyceros and
M. magister could suggest that the smaller crustaceans were competitively excluding the larger crustaceans, or simply proved physically irritating. However, in the experiments at 300 m in the Strait of Georgia, optodes were placed very close to the carcasses in one deployment and it was shown that extremely large numbers of amphipods caused a dramatic, but very localized, drawdown of oxygen [
25]. The experiment was repeated to confirm this result and a similar drawdown was noted [
33]. In experiments which exposed
Orchomene sp. to bait odors, increased respiration rates and elevated oxygen consumption was recorded for up to eight hours [
34], so it is very probable that the amphipods create a localized anoxia, repelling larger crustaceans. This is unlikely to affect the feeding amphipods as
O. obtusa has been experimentally shown to survive 10–33 h of anoxia [
35]. In these experiments, the optodes were not close enough to the carcasses to measure a localized drawdown and the numbers of amphipods were much fewer than those seen at 300 m, so may not have produced the same levels of hypoxia. This may be why the larger crustaceans remained at the carcasses and fed around the edges of the amphipod masses.
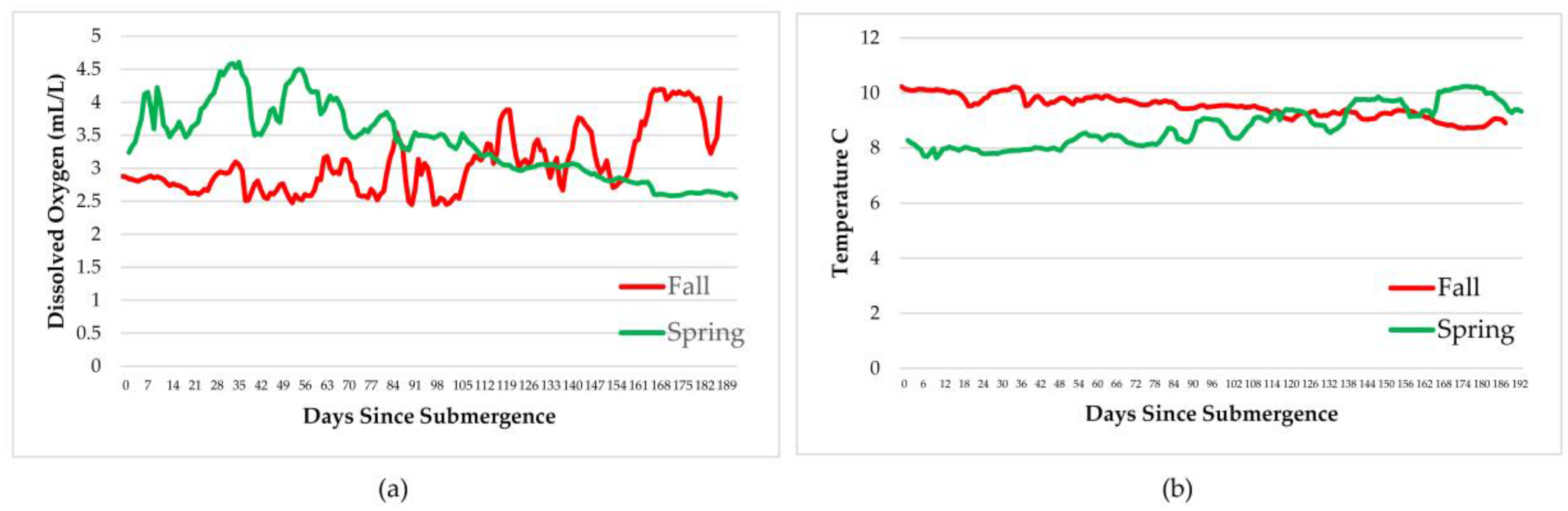
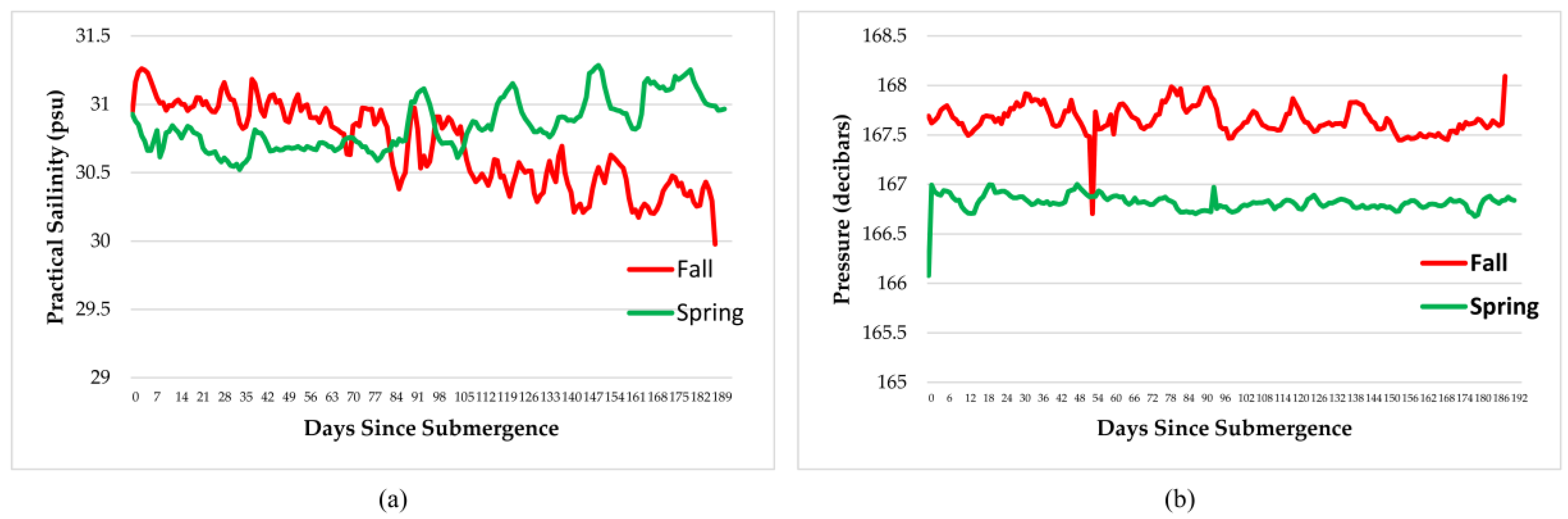
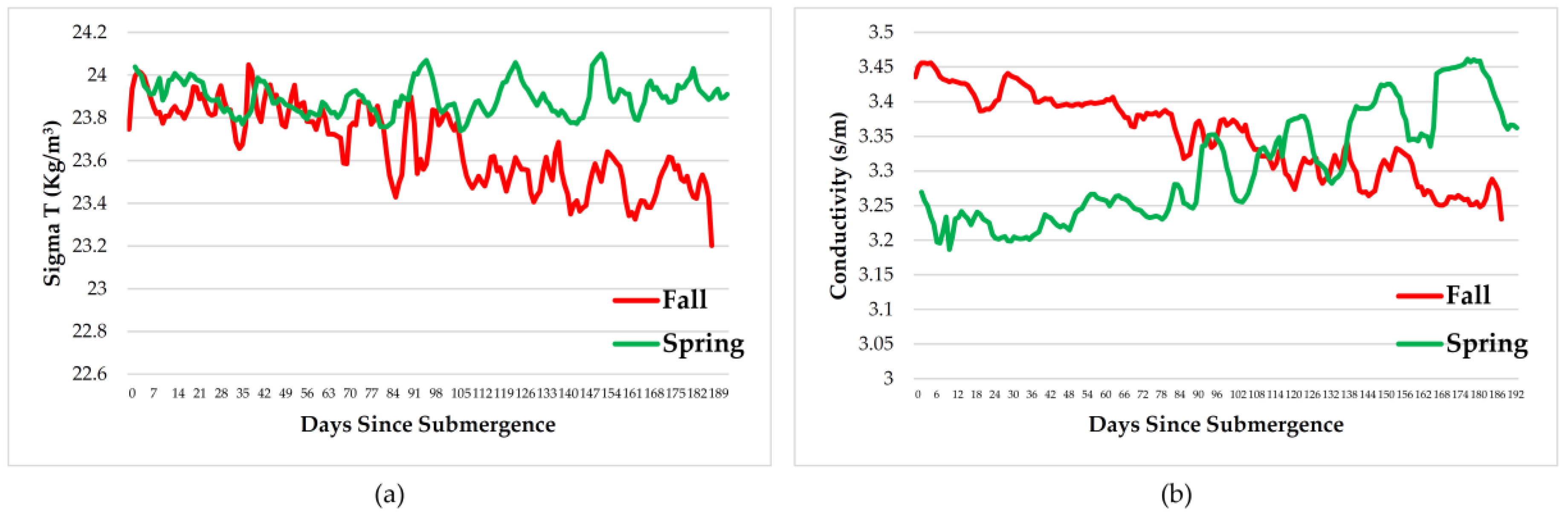
Measurements of dissolved oxygen, temperature, salinity, pressure, density, and conductivity varied over the time period and with season but remained within normal ranges for the species involved therefore, had little impact on the faunal scavenging, unlike that seen in Saanich Inlet, where dissolved oxygen levels drove the scavenging [
12].
Very few fish were associated with the carcasses. At 300 m in this Strait, bluntnose sixgill sharks were rapidly attracted to the spring carcasses in both a test run, in which a single deployed carcass was entirely consumed by a number of sharks, and in the actual experiments, where large pieces of tissue were removed by several sharks in the first 24 h after deployment until large masses of amphipods covered the carcasses [
25]. However, at 170 m, sharks did not feed on any of the carcasses and only one was seen in the vicinity briefly in fall. At 100 m in Saanich Inlet, on two single occasions, a shark removed part of the carcasses in fall [
12]. The bluntnose sixgill shark is typically found in deep waters with a few adults and more commonly, juveniles, in shallower waters [
36]. The Strait of Georgia is part of this very widely-distributed demersal species’ range, but local research using time lapse cameras has shown that this species is mostly present in the Strait in summer, with numbers increasing in June and peaking from mid-June to mid-July, and no specimens observed between October and May [
37]. These carcasses were deployed in March and late September so the spring carcasses would have already been skeletonized by the summer and the fall carcasses were deployed just at the end of this range. However, the spring carcasses at 300 m, as well as the preliminary test carcass were deployed in February and all attracted several sharks, so obviously some sharks are still present at this time of year, albeit perhaps in much lower numbers.
More shortspine thornyhead fish (
S. alascanus) were present in spring than in fall, and this was also observed at 300 m in Strait of Georgia [
25], in both these and the previous experiments, they frequently rested on the seabed for hours or days at a time, which is typical of these rockfish [
38]. However, in the experiments at 300 m, the rockfish seemed to just rest near the carcasses, but did not feed either on the invertebrates on the carcasses or on the carcasses themselves [
25]. In the present experiments, the rockfish did mostly rest near the carcasses, but on several occasions they were observed to attempt to catch a shrimp and were seen to feed on the carcass, although rarely, and at one point picked up a mouthful of bones and removed them from camera view (
Video S2). It is surprising they did not feed more on the arthropods as although their diet more commonly includes cephalopods, echinoderms, and copepods, they do eat some arthropods [
39]. At 300 m the carcasses were disarticulated almost entirely by the amphipods, but at 170 m, the spring carcasses remained articulated after skeletonization by both amphipods and shrimp, and were disarticulated by the action of these fish.
As was observed at 300 m in Strait of Georgia [
25] and in Saanich Inlet [
12], the carcasses were scavenged and consumed rather than progressing through the expected aquatic decomposition stages seen in other areas such as nearby Howe Sound [
11,
40], as well as Puget Sound in Washington State [
13], California’s Monterey Bay [
18], and in much warmer waters near Italy [
41]. However, in those experiments, the carcasses floated for a period of time due to bloat and, in the human case studies, the bodies were recovered after being washed ashore, thus, presumably had also floated for a period of time. In the experiments reported here the carcasses were at too great a depth for bloat to occur [
26], thus remaining in contact with the seafloor at all times, making them much more accessible to non-swimming or more weakly swimming arthropods.
The data generated here and in the previous series of experiments are valuable as they provide experimental evidence of the fate of mammalian carcasses (as human proxies) on the seafloor under these conditions. Most previous work relates to cadavers recovered from shallow waters or washed ashore which means that the body was floating in the water column. The majority of experimental or observational data from carcasses on the seabed pertains to extremely large carcasses, such as whale falls, which are not forensically valid proxies for human bodies. These data will be very valuable for recovery divers, coast guard, police, forensic pathologists, coroners and medical examiners in understanding the fate of a body in similar marine environments where recovery decisions need to be assessed.
Past case studies indicate the importance of understanding the impact of marine submergence. A body was recovered in the Gulf of Maine by a trawler after having been submerged for over 32 years. However, there was a distinct difference between the decomposition of the upper part of the body and the lower half which suggested that the two contiguous parts of the body had been exposed to different environments [
15]. The upper body was highly degraded, with the majority of bones lost and octopus eggs within the flight suit, yet the lower part of the body was well preserved, which led the authors to conclude that the upper part of the body had been in a well-oxygenated environment and the lower part was well preserved due to having been exposed to an anoxic environment suggestive of burial in silt [
15]. This is consistent with what we have earlier observed in low-oxygen environments [
12]. In another case, a body was recovered from clam flats at the mouth of the Damariscotta River, where it enters the Gulf of Maine, after being submerged for four weeks. A sea urchin spine and marine animal shell fragments showed that the body had entered the water in the ocean, and had been washed into the flats, and had, therefore, not drowned in the flats [
15].
Pigs are often used as human proxies [
22], so the results presented here indicate that it is probable that human bodies would be skeletonized in a similar manner in these waters at this depth and these seasons. This is valuable information for family members and recovery divers. Divers need to be aware of what to expect when searching for remains and understand that, at certain times of year, the body might be so thickly covered in amphipods that they may no longer be recognizable as a cadaver.
Although at greater depths in the Strait of Georgia, a cadaver can be expected to be skeletonized in three to four days [
25], at 170 m, soft tissue can be expected to remain on the carcasses for much longer. Season also had an impact, with only crab damage at first in fall, before amphipods arrived. Therefore, a body at this depth and in this habitat can be expected to be intact for several days or even weeks, and skin and other tissue might be recovered from the surrounding area. Fewer amphipods were present than at 300 m, but they could still prove a diving hazard, as they can be quite dangerous to divers when in close proximity as they are extremely irritating when they swarm over the divers’ face, biting and attempting to get under the mask and can cause a severe panic reaction and so pose a drowning risk [
26]. It is, therefore, important that recovery divers are fore-warned of the expected presence of amphipods and the expected conditions of the remains they are trying to locate.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}