
Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Insects
2.2. Plants
2.3. Commercial Chemicals Used as Stimuli
2.4. Electroantennogram (EAG) Recording
2.5. Dissection of Reproductive System
2.6. Moth Copulation
2.7. Effect of Host Plant Flower on Oviposition
2.8. Effect of Components of V. unguiculata Floral Volatiles on Oviposition
2.9. Effect of Concentrations of β-Caryophyllene on Oviposition
2.10. Testing the Roles of Olfaction and Gustation in the Reception of β-Caryophyllene
2.11. Data Analysis
3. Results
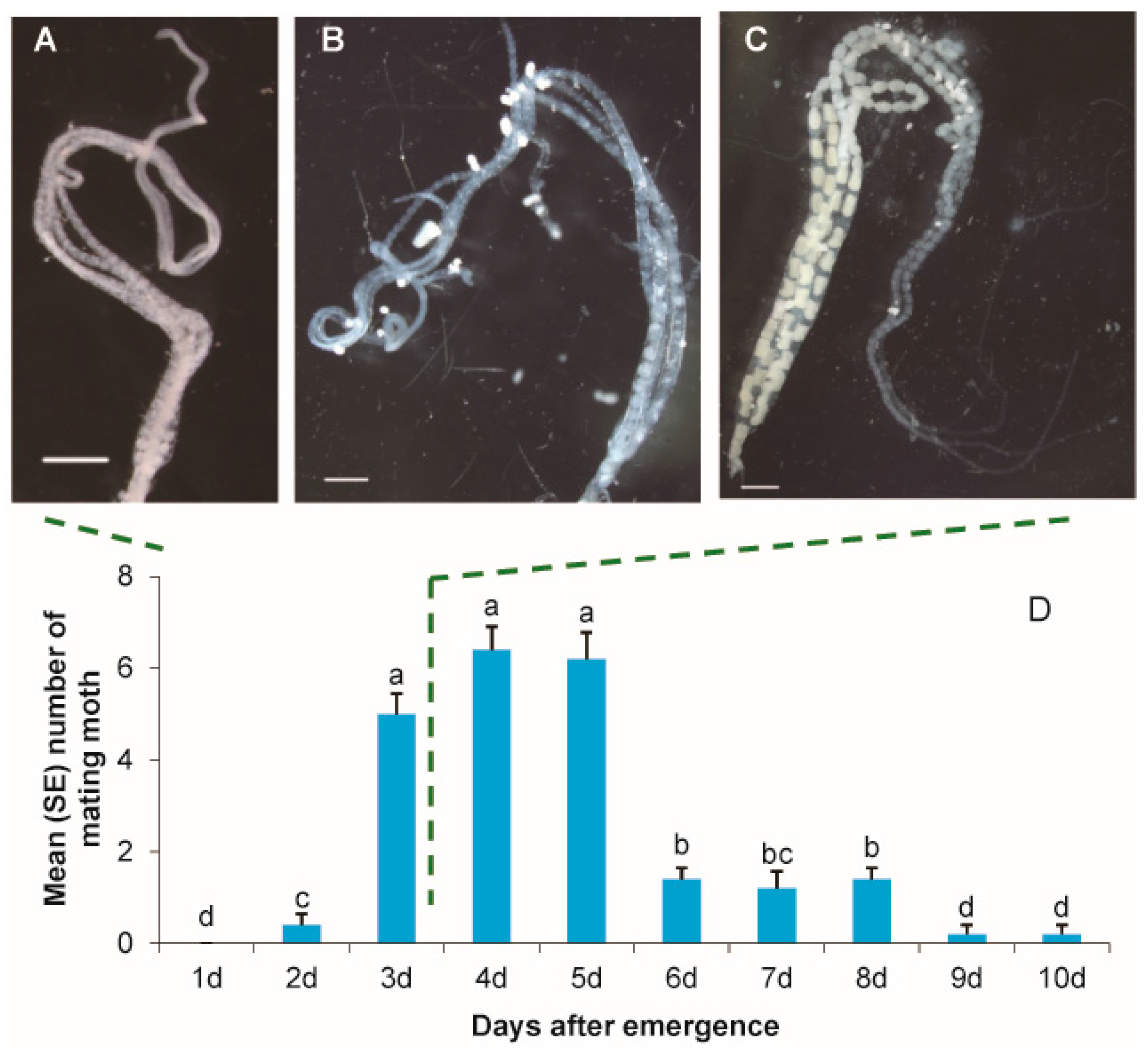
3.1. Egg Maturation and Female Mating Frequency by Age
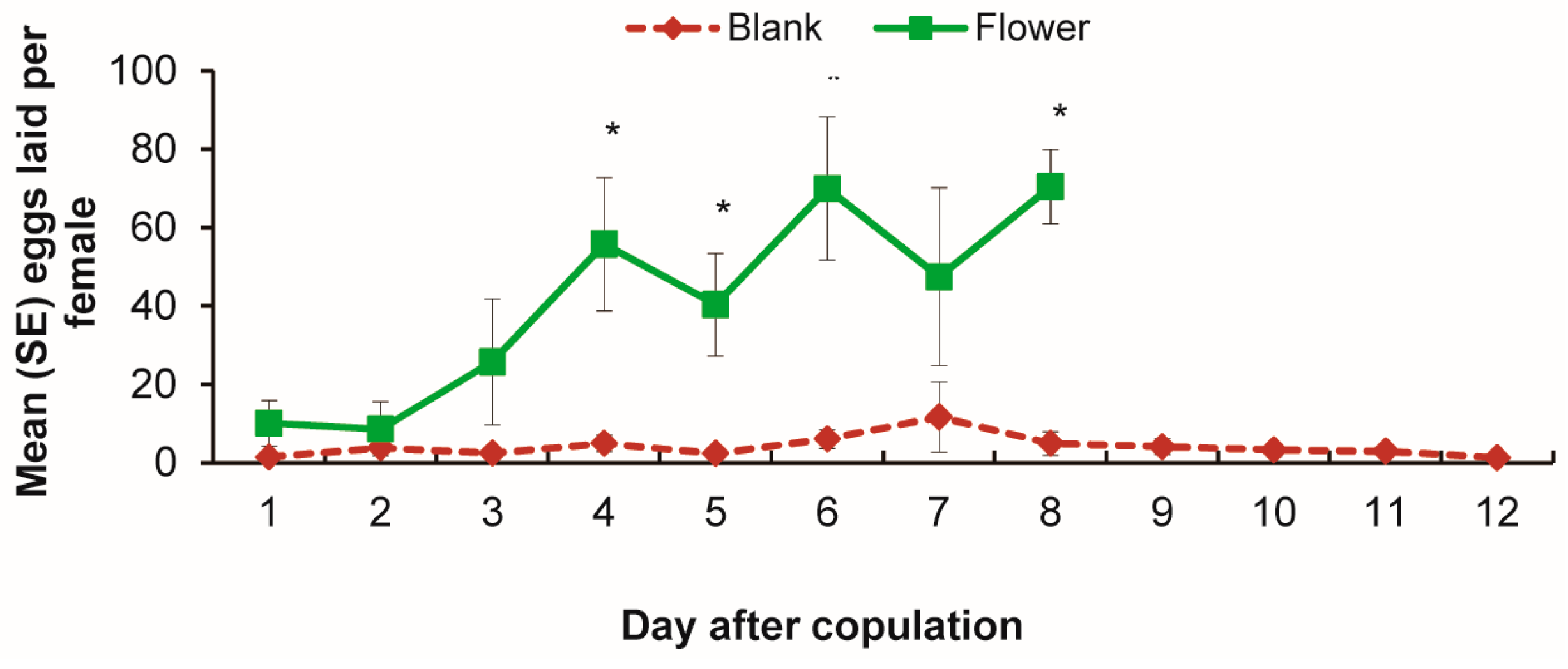
3.2. Oviposition by Age with and without Flowers Present
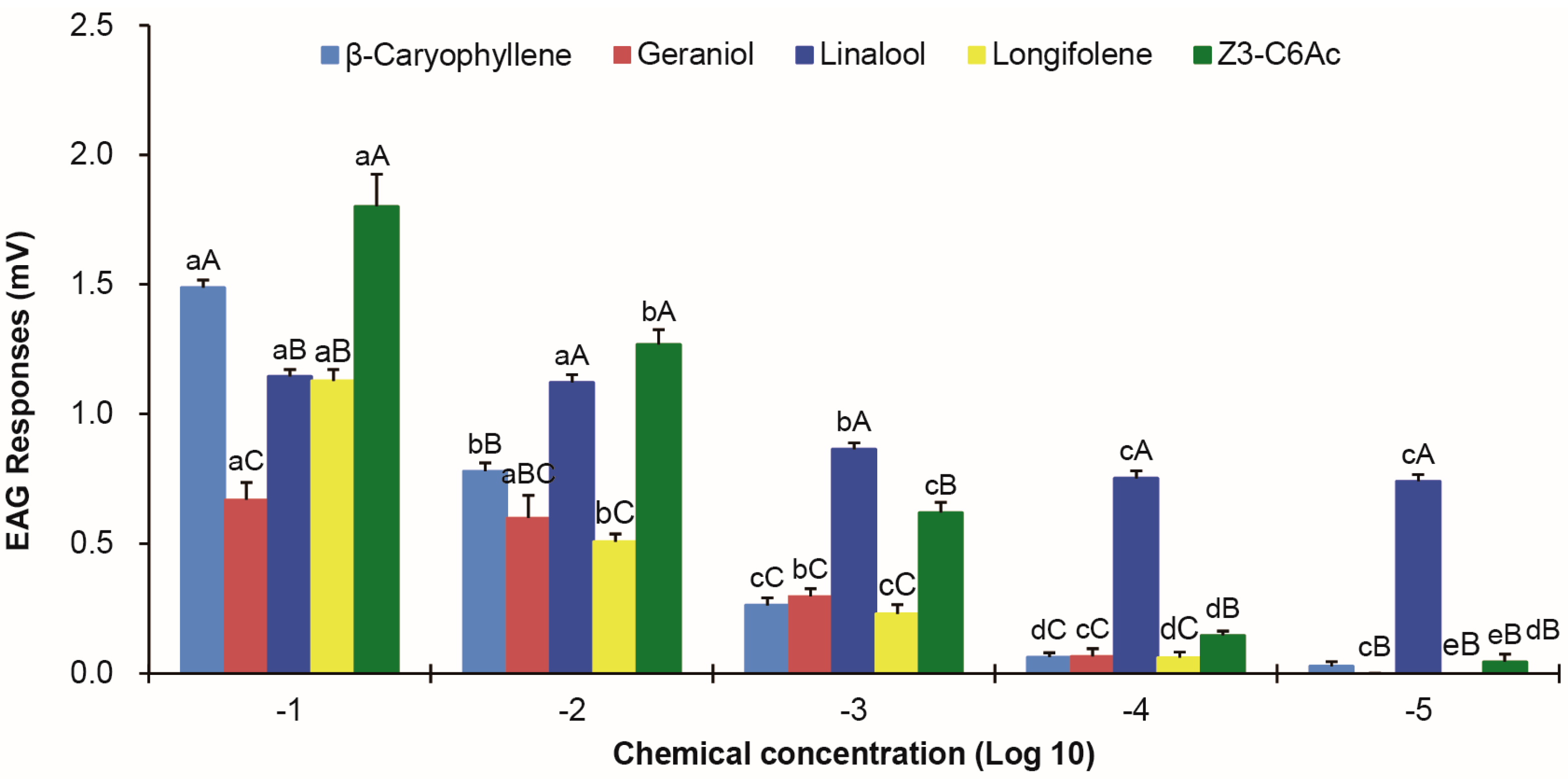
3.3. EAG Responses of Female Antennae to Components of V. unguiculata Floral Volatiles
3.4. V. unguiculata Floral Volatiles as Stimuli on Female Oviposition
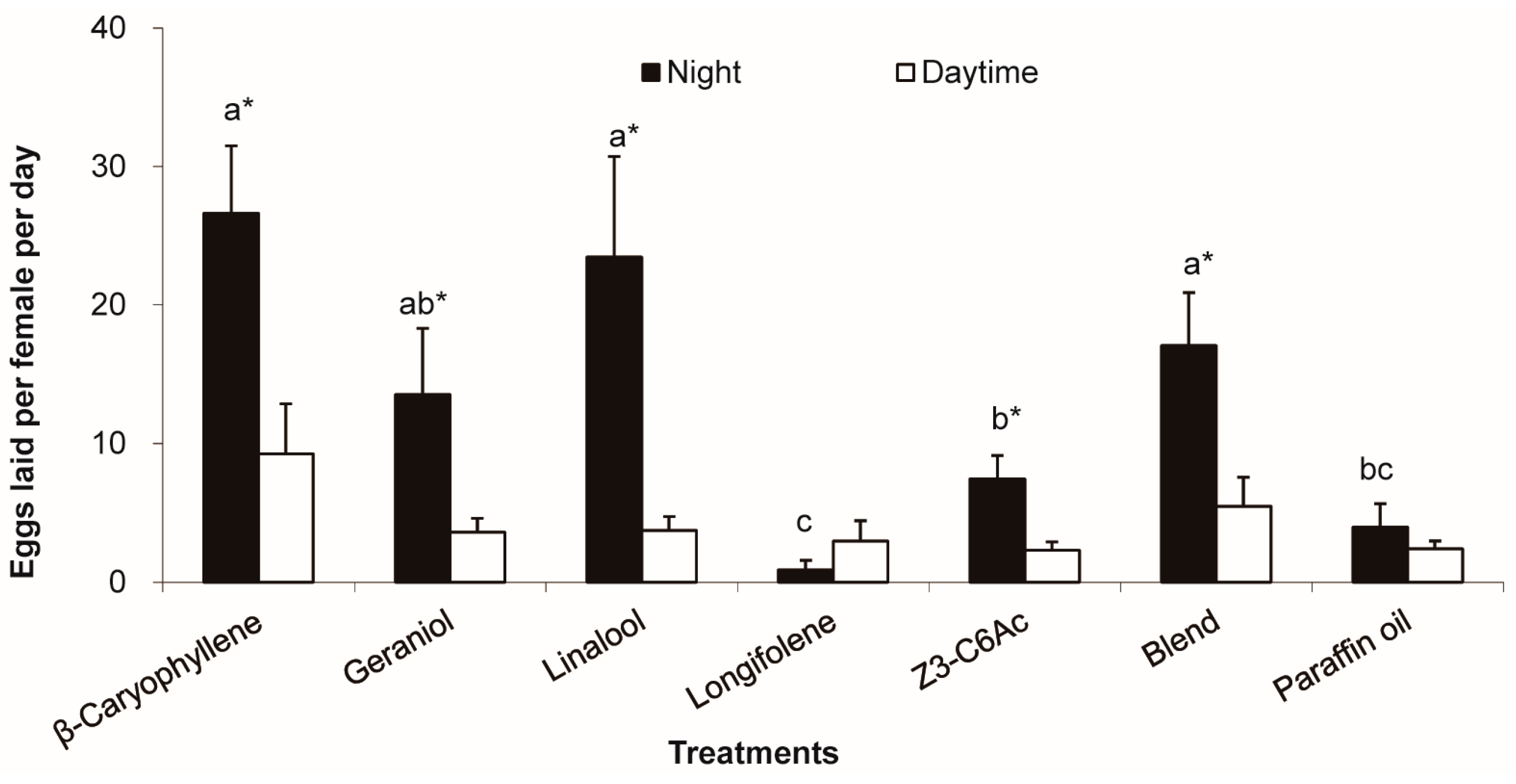
3.5. Stimulation of Host Floral Volatiles on Female Oviposition Restricted in Night
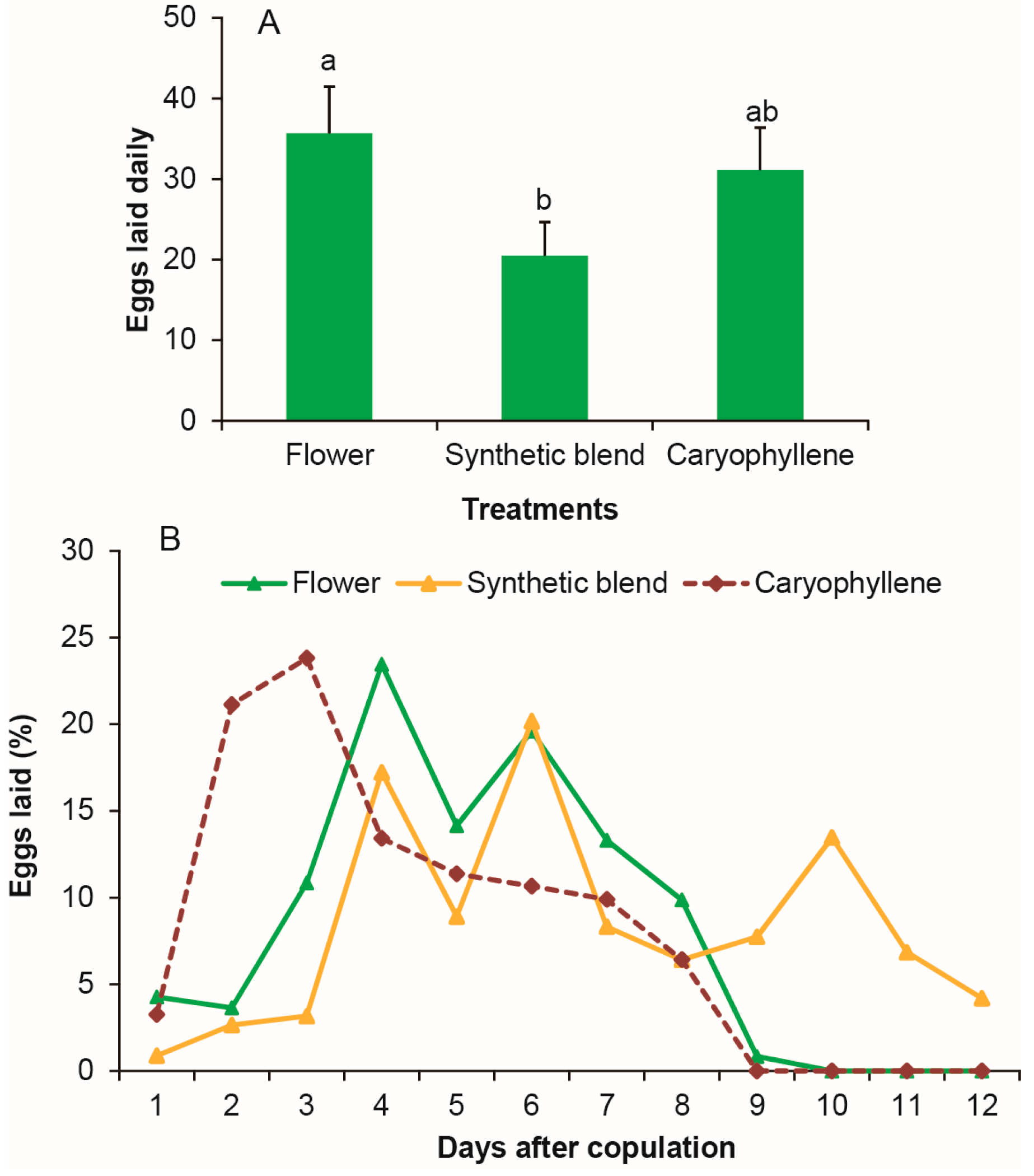
3.6. Stimulation of β-Caryophyllene on Female Oviposition as Strong as Flower
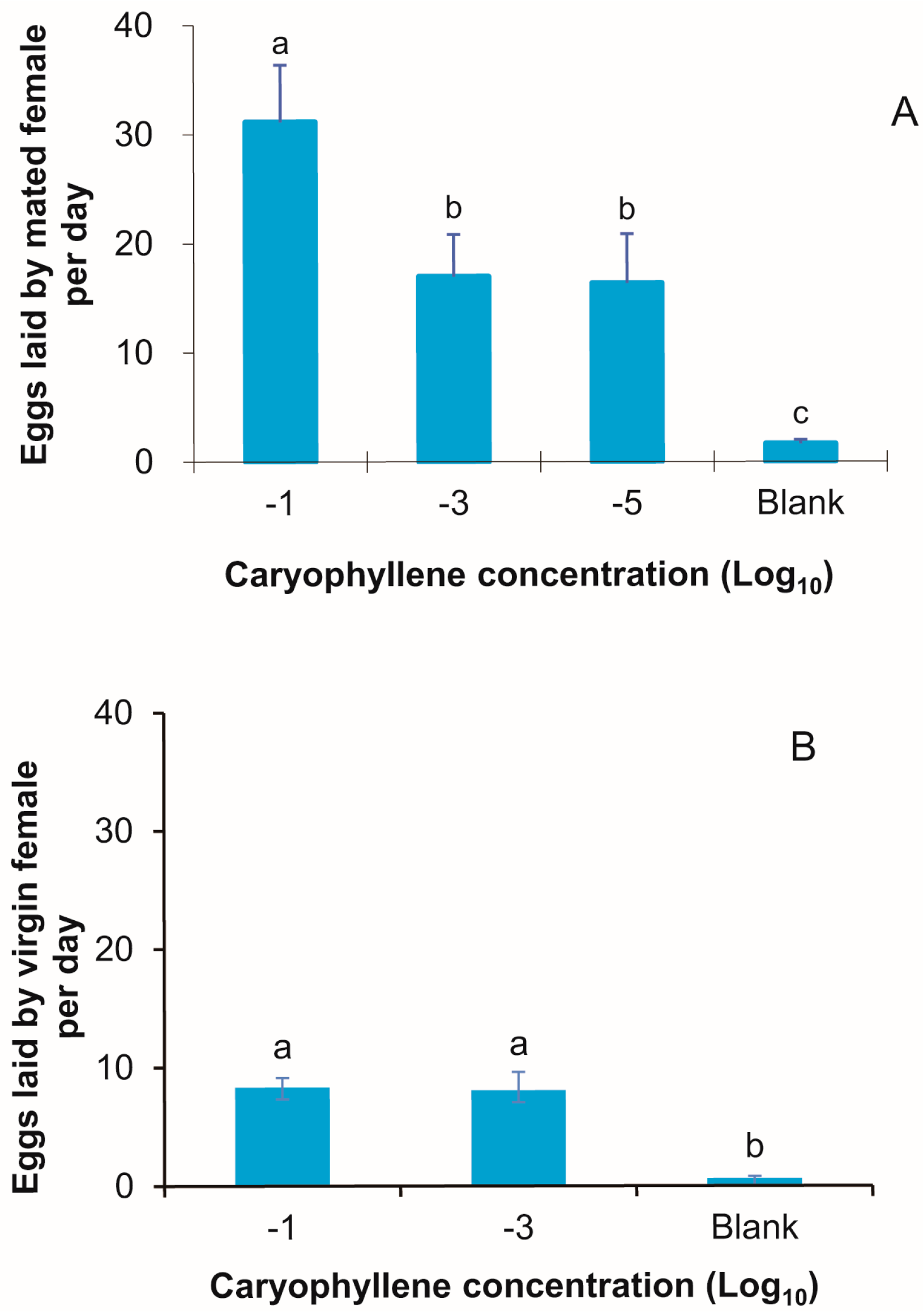
3.7. Dose-Response Relationship for β-Caryophyllene as an Oviposition Stimulus
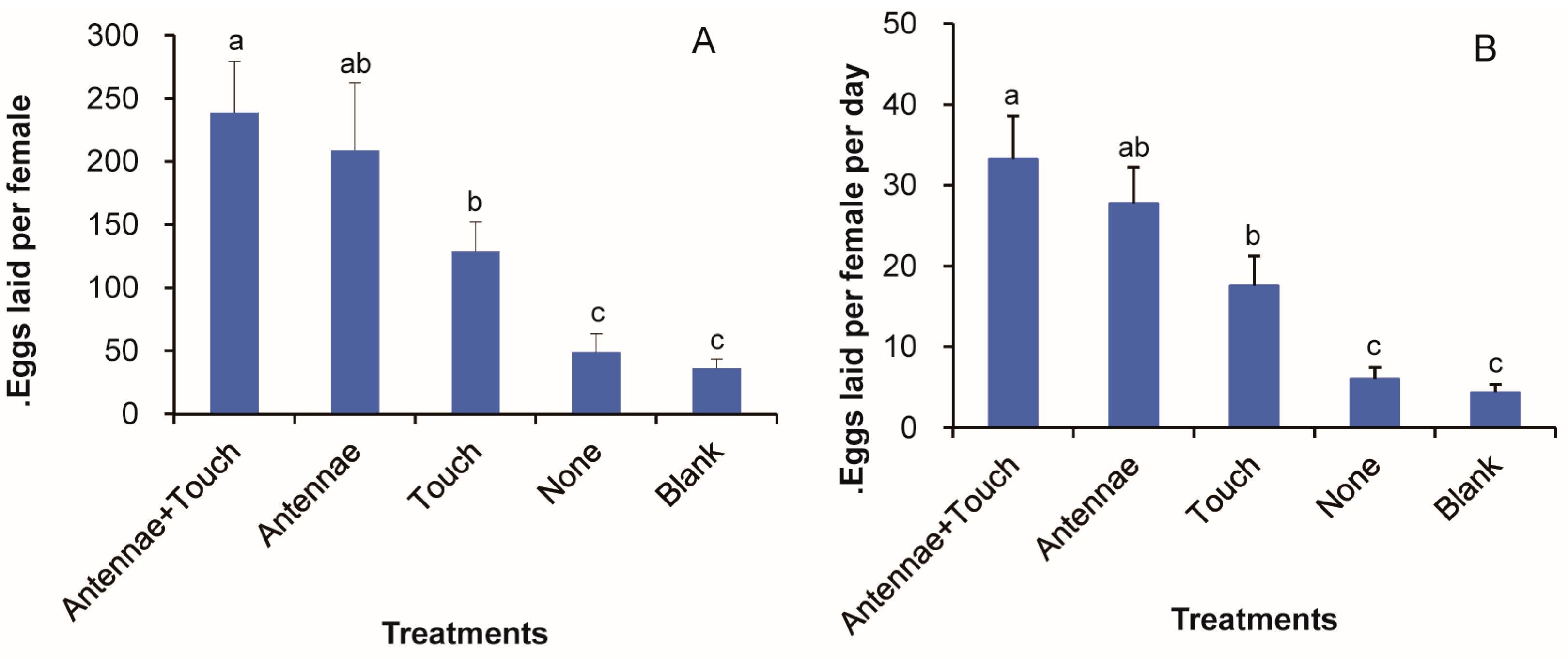
3.8. Roles of Olfaction and Gustation in the Mediation of the Floral Volatile Oviposition Stimulus
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Renwick, J.A.A.; Chew, F.S. Oviposition behavior in Lepidoptera. Annu. Rev. Entomol. 1994, 39, 377–400. [Google Scholar] [CrossRef]
- Rojas, J.C.; Wyatt, T.D.; Birch, M.C. Flight and oviposition behavior toward different host plant species by the cabbage moth, Mamestra brassicae (L.) (Lepidoptera: Noctuidae). J. Insect Behav. 2000, 13, 247–254. [Google Scholar] [CrossRef]
- Thompson, J.N.; Pellmyr, O. Evolution of oviposition behavior and host preference in Lepidoptera. Annu. Rev. Entomol. 1991, 36, 65–89. [Google Scholar] [CrossRef]
- Raguso, R.A. Wake up and smell the roses: The ecology and evolution of floral scent. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 549–569. [Google Scholar] [CrossRef]
- Riffell, J.A.; Alarcón, R.; Abrell, L.; Davidowitz, G.; Bronstein, J.L.; Hildebrand, J.G. Behavioral consequences of innate preferences and olfactory learning in hawkmoth–flower interactions. Proc. Natl. Acad. Sci. USA 2008, 105, 3404–3409. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.J.; Loon, J.J.A.V.; Renwick, J.A.A. Contact chemoreception of oviposition-stimulating glucosinolates and an oviposition-deterrent cardenolide in two subspecies of Pieris napi. Physiol. Entomol. 1995, 20, 164–174. [Google Scholar] [CrossRef]
- Hughes, P.R.; Renwick, J.A.A.; Lopez, K.D. New oviposition stimulants for the diamondback moth in cabbage. Entomol. Exp. Appl. 1997, 85, 281–283. [Google Scholar] [CrossRef]
- Lee, H.S.; Hieu, T.; Ahn, Y.J. Oviposition-stimulating activity of (E)-capsaicin identified in Capsicum annuum fruit and related compounds towards Helicoverpa assulta (Lepidoptera: Noctuidae). Chemoecology 2006, 16, 153–157. [Google Scholar] [CrossRef]
- Maher, N.; Thiery, D. Daphne gnidium, a possible native host plant of the European grapevine moth Lobesia botrana, stimulates its oviposition. Is a host shift relevant? Chemoecology 2006, 16, 135–144. [Google Scholar] [CrossRef]
- Macel, M.; Vrieling, K.; Macel, M.; Vrieling, K. Pyrrolizidine alkaloids as oviposition stimulants for the cinnabar moth, Tyria jacobaeae. J. Chem. Ecol. 2003, 29, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Mozuraitis, R.; Stranden, M.; Ramirez, M.I.; Borg-Karlson, A.K.; Mustaparta, H. (−)-Germacrene D increases attraction and oviposition by the tobacco budworm moth Heliothis virescens. Chem. Senses 2002, 27, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, C.; Shanower, T.G.; Bhagwat, V.R.; Khan, M.I.; Dodia, D.A. Relationship between Helicoverpa armigera Hubner and Maruca vitrata (Geyer) abundance, damage and yield loss in short-duration pigeonpeas. Int. J. Trop. Insect Sci. 2003, 23, 251–258. [Google Scholar] [CrossRef]
- Sharma, H.C.; Franzmann, B.A. Biology of the legume pod borer, Maruca vitrata (Fabricius) and its damage to pigeonpea and Adzuki bean. Int. J. Trop. Insect Sci. 2000, 20, 99–108. [Google Scholar] [CrossRef]
- Atachi, P.; Dannon, E.A.; Arodokoun, Y.D.; Tamo, M. Distribution and sampling of Maruca vitrata (Fabricius) (Lep., Pyralidae) larvae on Lonchocarpus sericeus (Poir) H.B. and K. J. Appl. Entomol. 2002, 126, 188–193. [Google Scholar] [CrossRef]
- Gopali, J.B.; Teggelli, R.; Mannur, D.M.; Yelshetty, S. Web-forming lepidopteran, Maruca vitrata (Geyer): An emerging and destructive pest in pigeonpea. Karnataka J. Agric. Sci. 2010, 23, 35–38. [Google Scholar]
- Wäckers, F.L.; Romeis, J.; van Rijn, P. Nectar and pollen feeding by insect herbivores and implications for multitrophic interactions. Annu. Rev. Entomol. 2006, 52, 301–323. [Google Scholar] [CrossRef] [PubMed]
- Bottenberg, H.; Tam, M.; Singh, B.B. Occurrence of phytophagous insects on wild Vigna sp. and cultivated cowpea: Comparing the relative importance of host-plant resistance and millet intercropping. Agric. Ecosyst. Environ. 1998, 70, 217–229. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S.D. A key role for floral scent in a wasp-pollination system in Eucomis (Hyacinthaceae). Ann. Bot. 2009, 103, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Beerwinkle, K.R.; Shaver, T.N.; Lopez, J.D. Field observations of adult emergence and feeding behavior of Helicoverpa zea (Lepidoptera: Noctuidae) on dallisgrass ergot honeydew. Environ. Entomol. 1993, 22, 554–558. [Google Scholar] [CrossRef]
- Gabel, B.; Thiéry, D.; Suchy, V.; Marion-Poll, F.; Hradsky, P.; Farkas, P. Floral volatiles of Tanacetum vulgare L. attractive to Lobesia botrana den. et schiff. females. J. Chem. Ecol. 1992, 18, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, C.N.K.; Walter, G.H. Polyphagy and primary host plants: Oviposition preference versus larval performance in the lepidopteran pest Helicoverpa armigera. Arthropod-Plant Interact. 2007, 1, 17–26. [Google Scholar] [CrossRef]
- Jallow, M.F.A.; Zalucki, M.P.; Fitt, G.P. Role of chemical cues from cotton in mediating host selection and oviposition behaviour in Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 1999, 38, 359–366. [Google Scholar] [CrossRef]
- Mitchell, E.R.; Tingle, F.C.; Heath, R.R. Flight activity of Heliothis virescens (F.) females (Lepidoptera, Noctuidae) with reference to host-plant volatiles. J. Chem. Ecol. 1991, 17, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Rasmussen, H.B.; Birkett, M.A.; Murray, D.A.; Pye, B.J.; Watts, N.P.; Williams, I.H. Behavioural and chemical ecology underlying the success of turnip rape (Brassica rapa) trap crops in protecting oilseed rape (Brassica napus) from the pollen beetle (Meligethes aeneus). Arthropod-Plant Interact. 2007, 1, 57–67. [Google Scholar] [CrossRef]
- Atachi, P.; Hountondji, F.C.C. Olfactory responses of Maruca vitrata (Fabricius) larvae to reproductive parts of different varieties of cowpea, Vigna unguiculata (L.) Walp. Insect Sci. Appl. 2000, 20, 117–121. [Google Scholar] [CrossRef]
- Halder, J.; Srinivasan, S.; Muralikrishna, T. Biochemical basis of resistance to spotted pod borer, Maruca vitrata (Geyer) in Mungbean. J. Entomol. Res. 2006, 30, 313–316. [Google Scholar]
- Morris, B.D.; Foster, S.P.; Grugel, S.; Charlet, L.D. Isolation of the diterpenoids, ent-kauran-16alpha-ol and ent-atisan-16alpha-ol, from sunflowers, as oviposition stimulants for the banded sunflower moth, Cochylis hospes. J. Chem. Ecol. 2005, 31, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.D.; Charlet, L.D.; Foster, S.P. Isolation of three diterpenoid acids from sunflowers, as oviposition stimulants for the banded sunflower moth, Cochylis hospes. J. Chem. Ecol. 2009, 35, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Bendera, M.; Ekesi, S.; Ndung’u, M.; Srinivasan, R.; Torto, B. A major host plant volatile, 1-octen-3-ol, contributes to mating in the legume pod borer, Maruca vitrata (Fabricius) (Lepidoptera: Crambidae). Naturwissenschaften 2015, 102, 47. [Google Scholar] [CrossRef] [PubMed]
- Ager, C.A. Analysis of Chemical Composition of Cowpea Floral Volatiles and Nectar. Master’s Thesis, Kenyatta University, Kahawa, November 2009. [Google Scholar]
- Knudsen, J.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Eltz, T.; Ayasse, M.; Lunau, K. Species-specific antennal responses to tibial fragrances by male orchid bees. J. Chem. Ecol. 2006, 32, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Parachnowitsch, A.L.; Raguso, R.A.; Kessler, A. Phenotypic selection to increase floral scent emission, but not flower size or colour in bee-pollinated Penstemon digitalis. New Phytol. 2012, 195, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Galizia, C.G.; Kunze, J.; Gumbert, A.; Borg-Karlson, A.-K.; Sachse, S.; Markl, C.; Menzel, R. Relationship of visual and olfactory signal parameters in a food-deceptive flower mimicry system. Behav. Ecol. 2005, 16, 159–168. [Google Scholar] [CrossRef]
- Koschier, E.; De Kogel, W.; Visser, J.H. Assessing the attractiveness of volatile plant compounds to western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 2000, 26, 2643–2655. [Google Scholar] [CrossRef]
- Ruther, J. Male-biassed response of garden chafer, Phyllopertha horticola L., to leaf alcohol and attraction of both sexes to floral plant volatiles. Chemoecology 2004, 14, 187–192. [Google Scholar] [CrossRef]
- Kessler, D.; Baldwin, I.T. Making sense of nectar scents: The effects of nectar secondary metabolites on floral visitors of Nicotiana attenuata. Plant J. 2007, 49, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.-D.; Fang, J.-L.; Li, Z.-J. Bionomics and control of the legume pod borer Maruca testulalis (Geyer). Acta Entomol. Sin. 1985, 28, 51–59. [Google Scholar]
- Yan, F.; Du, Y.; Han, X. A comparative study on the electroantenogram responses of three aphid species to plant volatiles. Insect Sci. 1994, 1, 53–66. [Google Scholar] [CrossRef]
- Oghiakhe, S. Effect of cowpea cultivar, plant part and growth phenology on oviposition behaviour of the legume pod borer, Maruca testulalis Geyer (Lep., Pyralidae). J. Appl. Entomol. 1996, 120, 549–553. [Google Scholar] [CrossRef]
- Sharma, H.C. Bionomics, host plant resistance, and management of the legume pod borer, Maruca vitrata—A review. Crop Prot. 1998, 17, 373–386. [Google Scholar] [CrossRef]
- Imosanen; Singh, H.K.B. Incidence of Helicoverpa armigera (Hub.) and Maruca vitrata (Geyer) on pigeonpea under Medzephema conditions of Nagaland. J. Appl. Zool. Res. 2005, 16, 85–86. [Google Scholar]
- Liao, C.T.; Lin, C.S. Occurrence of the legume pod borer, Maruca testulalis Geyer (Lepidoptera: Pyralidae) on cowpea (Vigna nguiculata Walp) and its insecticides application trial. Plant Prot. Bull. 2000, 42, 213–222. [Google Scholar]
- Sonune, V.R.; Bharodia, R.K.; Jethva, D.M.; Dabhade, P.L. Seasonal incidence of spotted pod borer, Maruca testulalis (Geyer) on blackgram. Legum. Res. 2010, 33, 61–63. [Google Scholar]
- Porter, A.E.A.; Griffiths, D.W.; Robertson, G.W.; Sexton, R. Floral volatiles of the sweet pea Lathyrus odoratus. Phytochemistry 1999, 51, 211–214. [Google Scholar] [CrossRef]
- Light, D.M.; Kamm, J.A.; Buttery, R.G. Electroantennogram response of alfalfa seed chalcid, Bruchophagus roddi (Hymenoptera: Eurytomidae) to host- and nonhost-plant volatiles. J. Chem. Ecol. 1992, 18, 333–352. [Google Scholar] [CrossRef] [PubMed]
- Kamdem, D.P.; Gruber, K.; Barkman, T.; Gage, D.A. Characterization of black locust floral fragrance. J. Essent. Oil Res. 1994, 6, 199–200. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Groth, I.; Bergström, G. Pollen advertisement: Chemical contrasts between wholeflower and pollen odors. Am. J. Bot. 1996, 83, 877–885. [Google Scholar] [CrossRef]
- Turlings, T.; Benrey, B. Effects of plant metabolites on the behavior and development of parasitic wasps. Ecoscience 1998, 5, 321–333. [Google Scholar] [CrossRef]
- Verheggen, F.; Arnaud, L.; Bartram, S.; Gohy, M.; Haubruge, E. Aphid and plant volatiles induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol. 2008, 34, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Colazza, S.; McElfresh, J.S.; Millar, J. Identification of volatile synomones, induced by Nezara viridula feeding and oviposition on bean spp., that attract the egg parasitoid Trissolcus basalis. J. Chem. Ecol. 2004, 30, 945–964. [Google Scholar] [CrossRef] [PubMed]
- Wegener, R.; Schulz, S.; Meiners, T.; Hadwich, K.; Hilker, M. Analysis of volatiles induced by oviposition of elm leaf beetle Xanthogaleruca luteola on Ulmus minor. J. Chem. Ecol. 2001, 27, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Alhmedi, A.; Haubruge, E.; Francis, F. Identification of limonene as a potential kairomone of the harlequin ladybird Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 2010, 107, 541–548. [Google Scholar] [CrossRef]
- Verheggen, F.; Fagel, Q.; Heuskin, S.; Lognay, G.; Francis, F.; Haubruge, E. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis pallas, to sesquiterpene semiochemicals. J. Chem. Ecol. 2007, 33, 2148–2155. [Google Scholar] [CrossRef] [PubMed]
- Bichão, H.; Borg-Karlson, A.K.; Araújo, J.; Mustaparta, H. Identification of plant odours activating receptor neurones in the weevil Pissodes notatus F. (Coleoptera, Curculionidae). J. Comp. Physiol. A 2003, 189, 203–212. [Google Scholar]
- Campbell, C.A.M.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Spring migration of damson-hop aphid, Phorodon humuli (Homoptera, Aphididae), and summer host plant-derived semiochemicals released on feeding. J. Chem. Ecol. 1993, 19, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Johne, A.B.; Sprauer, S.; Weißbecker, B.; Schütz, S. Influence of flower odor compounds on oviposition of the horse chestnut leaf miner Cameraria ohridella (Deschka and Dimic). Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2006, 14, 355–360. [Google Scholar]
- Furtado, R.; Baptista, J.; Lima, E.; Paiva, L.; Barroso, J.G.; Rosa, J.S.; Oliveira, L. Chemical composition and biological activities of Laurus essential oils from different Macaronesian Islands. Biochem. Syst. Ecol. 2014, 55, 333–341. [Google Scholar] [CrossRef]
- Ioannou, C.S.; Papadopoulos, N.T.; Kouloussis, N.A.; Tananaki, C.I.; Katsoyannos, B.I. Essential oils of citrus fruit stimulate oviposition in the Mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae). Physiol. Entomol. 2012, 37, 330–339. [Google Scholar] [CrossRef]
- Koul, O.; Singh, R.; Kaur, B.; Kanda, D. Comparative study on the behavioral response and acute toxicity of some essential oil compounds and their binary mixtures to larvae of Helicoverpa armigera, Spodoptera litura and Chilo partellus. Ind. Crop. Prod. 2013, 49, 428–436. [Google Scholar] [CrossRef]
- McCallum, E.J.; Cunningham, J.P.; Lucker, J.; Zalucki, M.P.; De Voss, J.J.; Botella, J.R. Increased plant volatile production affects oviposition, but not larval development, in the moth Helicoverpa armigera. J. Exp. Biol. 2011, 214, 3672–3677. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.X.; Cai, X.M.; Bian, L.; Luo, Z.X.; Xin, Z.J.; Chen, Z.M. Electrophysiological and behavioral responses of Chrysopa phyllochroma (Neuroptera: Chrysopidae) to plant volatiles. Environ. Entomol. 2015, 44, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Reisenman, C.E.; Riffell, J.A.; Bernays, E.A.; Hildebrand, J.G. Antagonistic effects of floral scent in an insect-plant interaction. Proc. R. Soc. B Biol. Sci. 2010, 277, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Lwande, W.; McDowell, P.G.; Amiani, H.; Amoke, P. Analysis of airborne volatiles of cowpea. Phytochemistry 1989, 28, 421–423. [Google Scholar] [CrossRef]
- Van Den Boom, C.E.M.; Van Beek, T.A.; Posthumus, M.A.; De Groot, A.; Dicke, M. Qualitative and quantitative variation among volatile profiles induced by Tetranychus urticae feeding on plants from various families. J. Chem. Ecol. 2004, 30, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.F.; Qiao, H.L.; Wang, X.P.; Zhou, X.M.; Wang, X.Q.; Lei, C.L. Adult behavior and circadian rhythm of sex pheromone production and release of the legume pod borer, Maruca vitrata (Fabricius) (Lepidoptera: Pyralidae). Acta Entomol. Sin. 2007, 50, 335–342. [Google Scholar]
- Ige, O.E.; Olotuah, O.F.; Akerele, V. Floral biology and pollination ecology of cowpea (Vigna unguiculata L. Walp). Mod. Appl. Sci. 2011, 5, 74–82. [Google Scholar]
- Uematsu, H.; Yoshikawa, K. Changes in copulation and oviposition time of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Jpn. J. Appl. Entomol. Zool. 2002, 46, 81–87. [Google Scholar] [CrossRef]
- Iwaizumi, R.; Arakawa, K.; Koshio, C. Nocturnal flight activities of the female Asian gypsy moth, Lymantria dispar (Linnaeus) (Lepidoptera: Lymantriidae). Appl. Entomol. Zool. 2010, 45, 121–128. [Google Scholar] [CrossRef]
- Wan, X.L.; Qian, K.; Du, Y.J. Synthetic pheromones and plant volatiles alter the expression of chemosensory genes in Spodoptera exigua. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Meiners, T. Plants and insect eggs: How do they affect each other? Phytochemistry 2011, 72, 1612–1623. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, S.T.; Guedes, R.N.; Demuner, A.J.; Lima, E.R. Effect of coffee alkaloids and phenolics on egg-laying by the coffee leaf miner Leucoptera coffeella. Bull. Entomol. Res. 2008, 98, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Fatzinger, C.W.; Merkel, E.P. Oviposition and feeding preferences of the southern pine coneworm (Lepidoptera: Pyralidae) for different host-plant materials and observations on monoterpenes as an oviposition stimulant. J. Chem. Ecol. 1985, 11, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, J.; Shen, Y.-L.; Liu, F.-Y.; Du, Y.-J. Electroantennogram responses of Maruca testulalis (Lepidoptera: Pyralidae) to plant volatiles and sex pheromone. Yingyong Shengtai Xuebao 2009, 20, 1973–1979. [Google Scholar] [PubMed]
- Tsuchihara, K.; Hisatomi, O.; Tokunaga, F.; Asaoka, K. An oviposition stimulant binding protein in a butterfly: Immunohistochemical localization and electrophysiological responses to plant compounds. Commun. Integr. Biol. 2009, 2, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Valmalette, J.C.; Raad, H.; Qiu, N.; Ohara, S.; Capovilla, M.; Robichon, A. Nano-architecture of gustatory chemosensory bristles and trachea in Drosophila wings. Sci. Rep. 2015, 5, 14198. [Google Scholar] [CrossRef] [PubMed]
- Justus, K.A.; Mitchell, B.K. Oviposition site selection by the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae). J. Insect Behav. 1996, 9, 887–898. [Google Scholar] [CrossRef]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, B.; Qian, K.; Du, Y.-J. Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects 2017, 8, 60. https://doi.org/10.3390/insects8020060
Feng B, Qian K, Du Y-J. Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects. 2017; 8(2):60. https://doi.org/10.3390/insects8020060
Chicago/Turabian StyleFeng, Bo, Kai Qian, and Yong-Jun Du. 2017. "Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata" Insects 8, no. 2: 60. https://doi.org/10.3390/insects8020060
APA StyleFeng, B., Qian, K., & Du, Y. -J. (2017). Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects, 8(2), 60. https://doi.org/10.3390/insects8020060