
Do Refuge Plants Favour Natural Pest Control in Maize Crops?

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
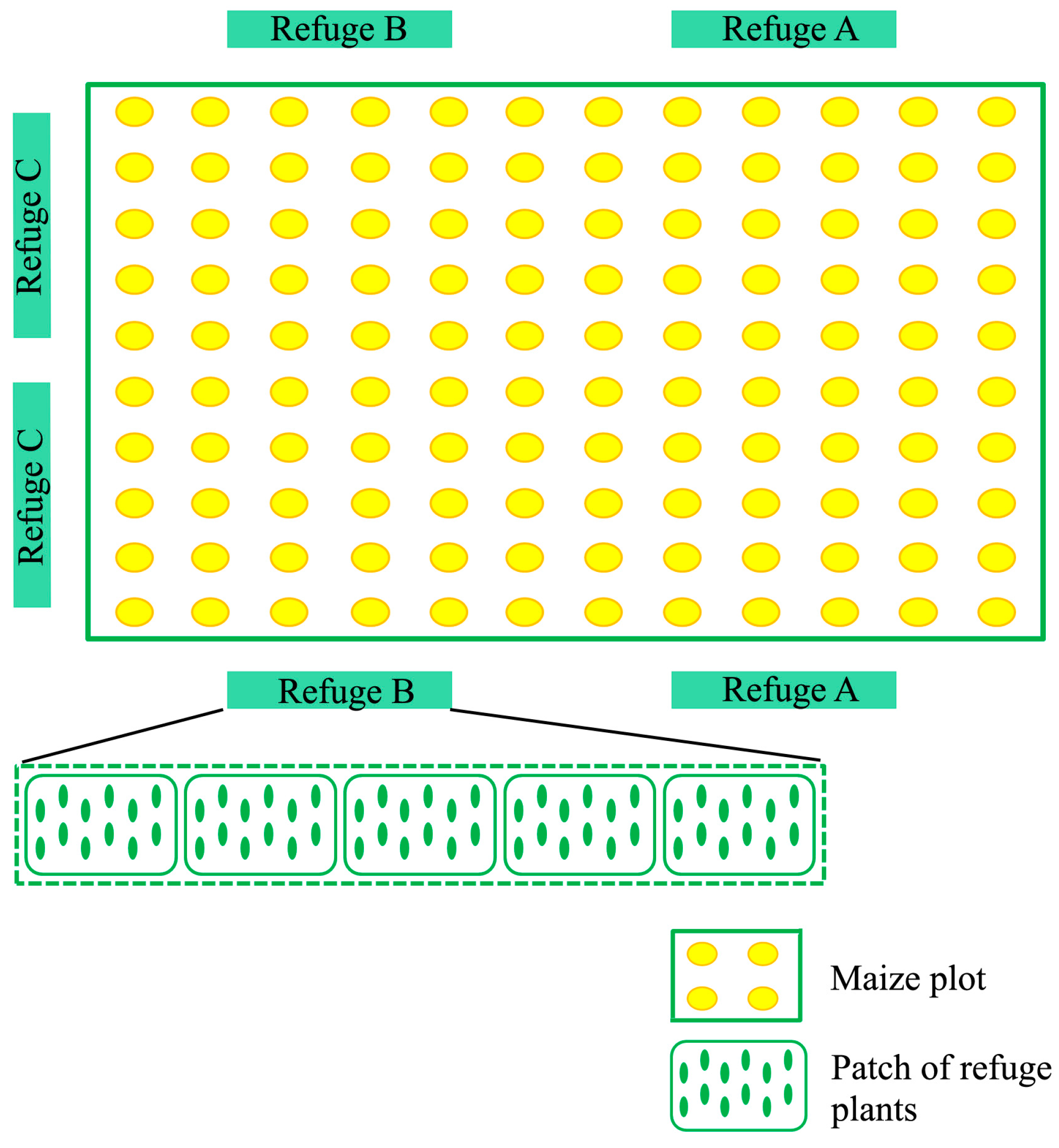
2.2. Field Experiments Setup
2.3. Insect Sampling and Identification
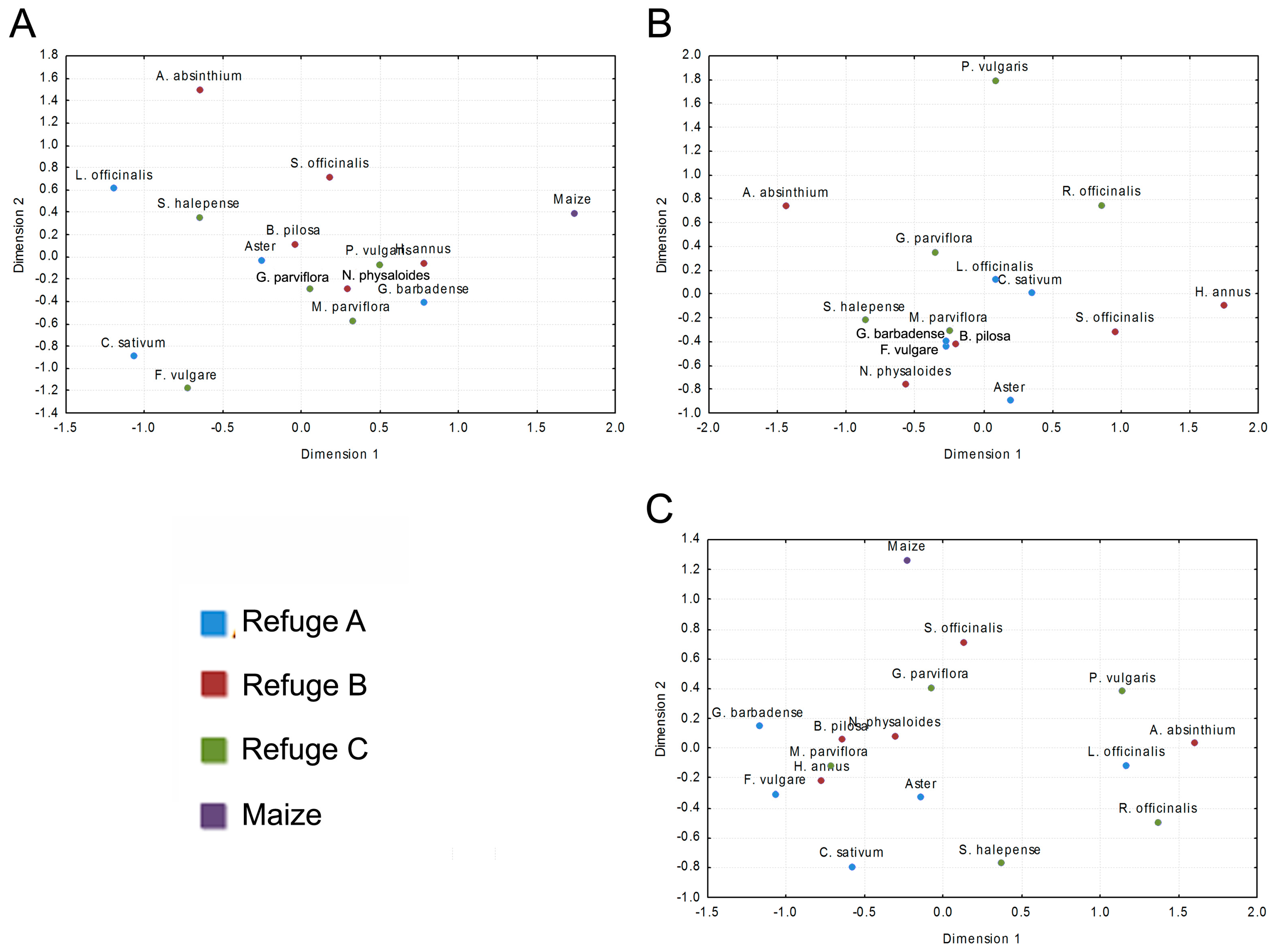
2.4. Data Analysis
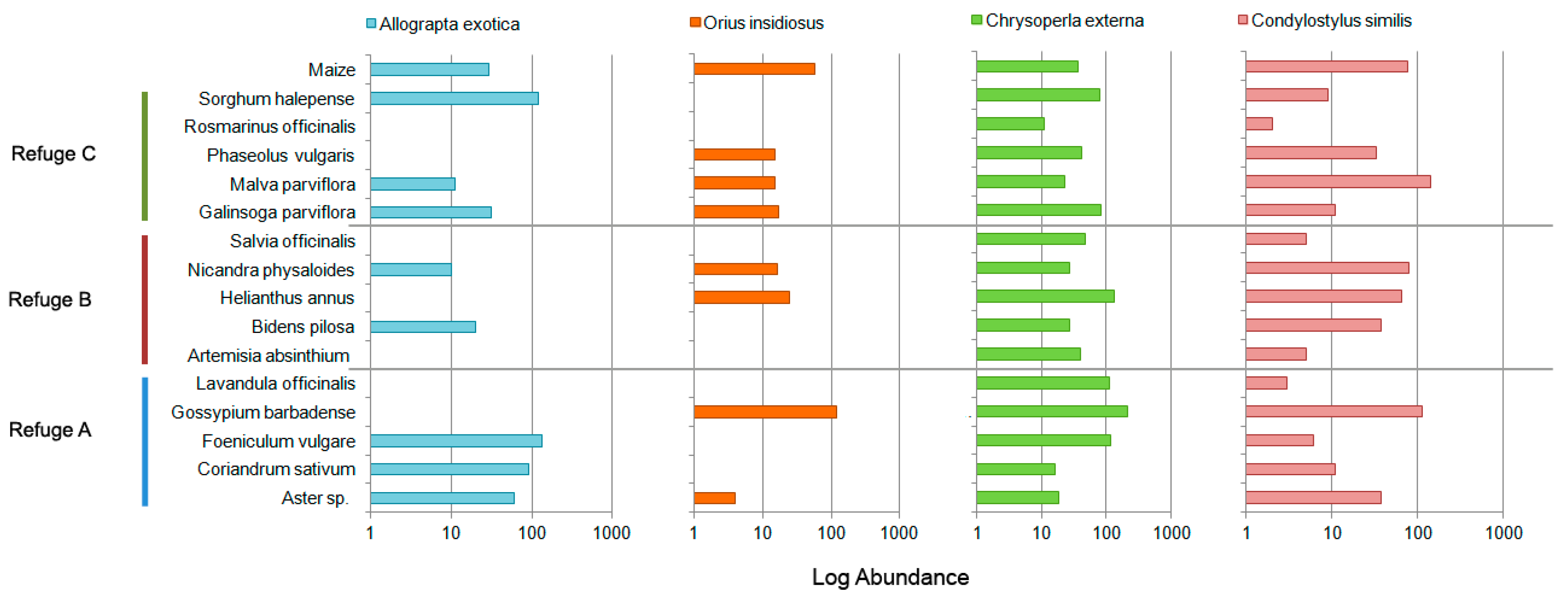
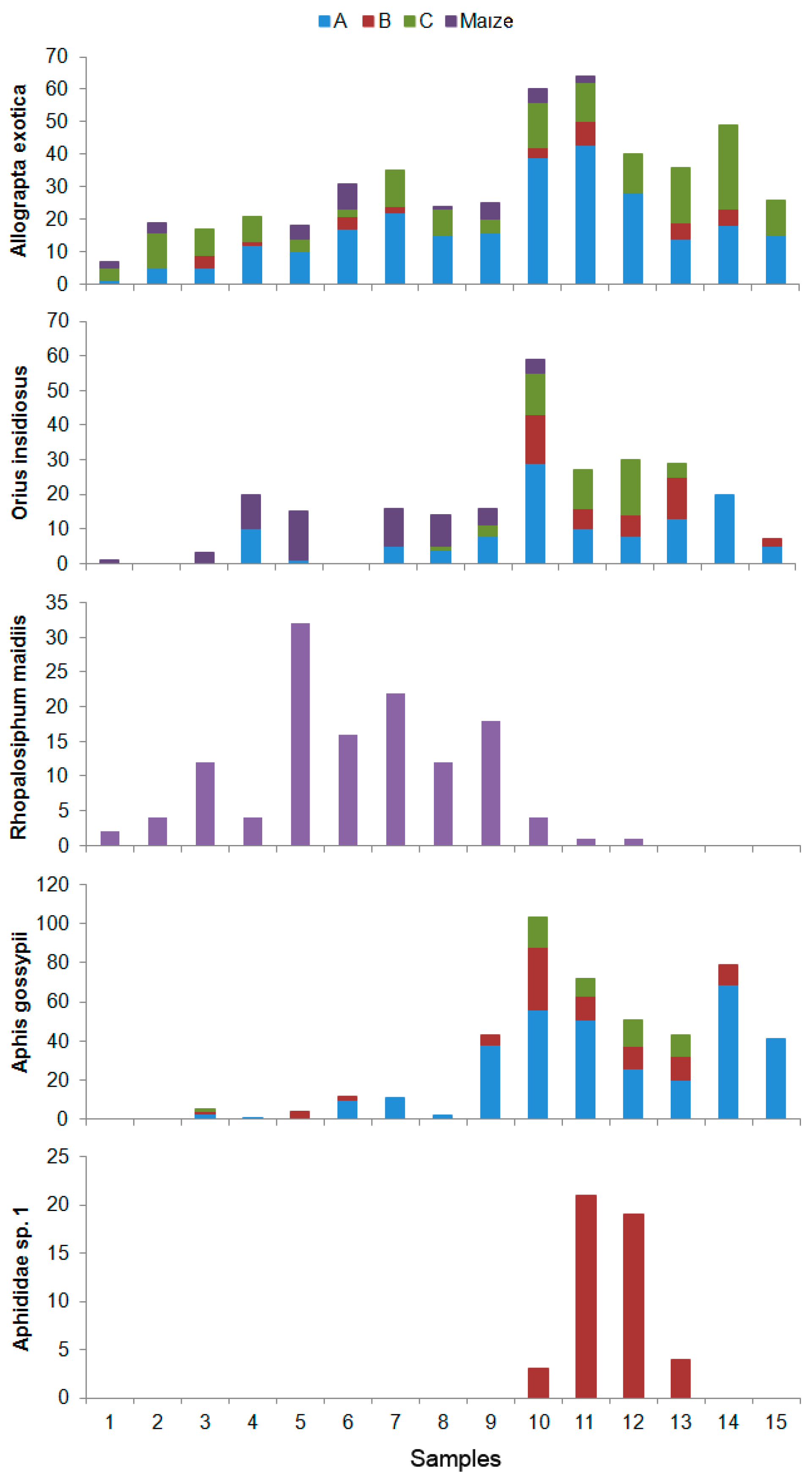
3. Results
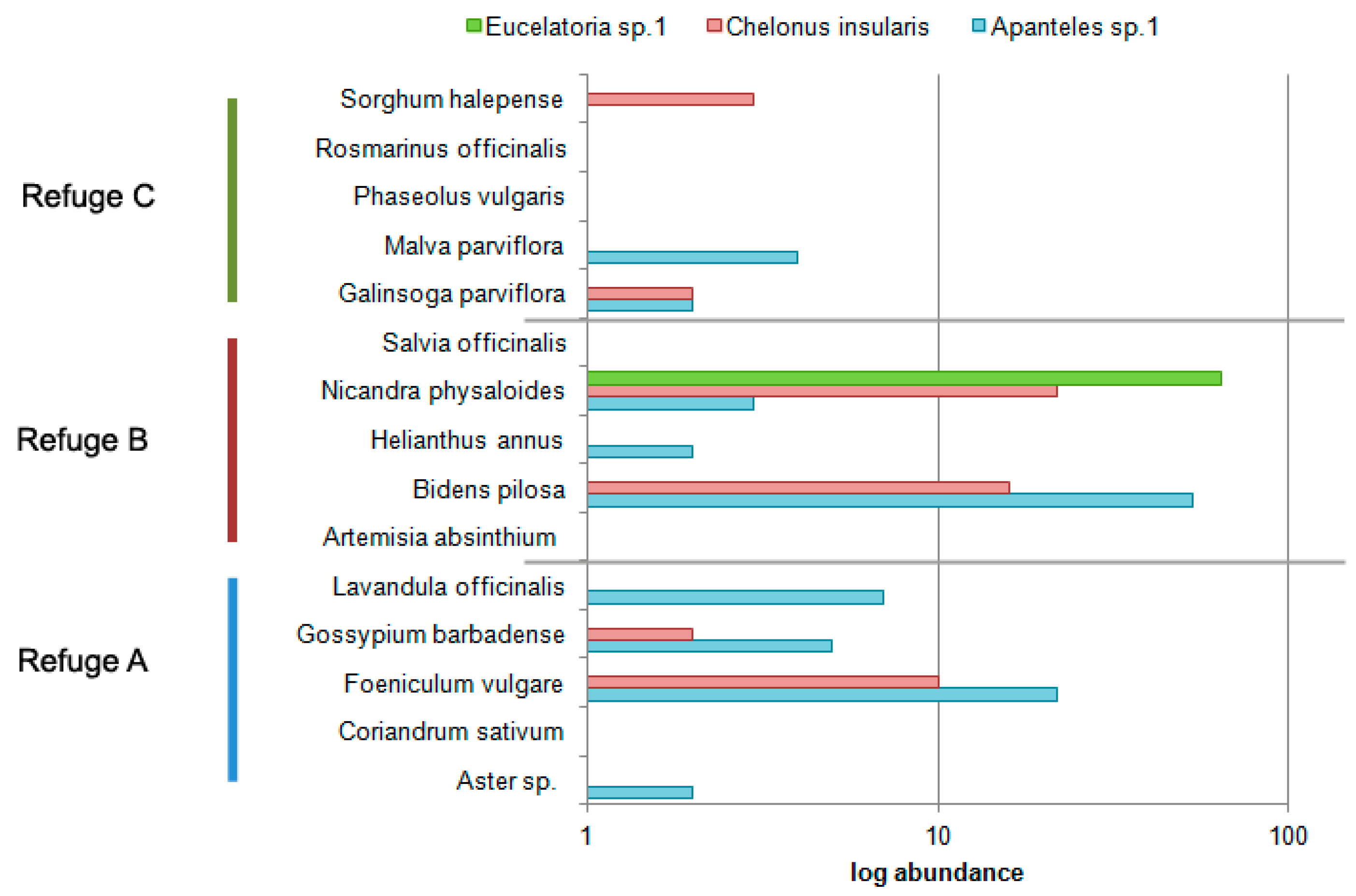
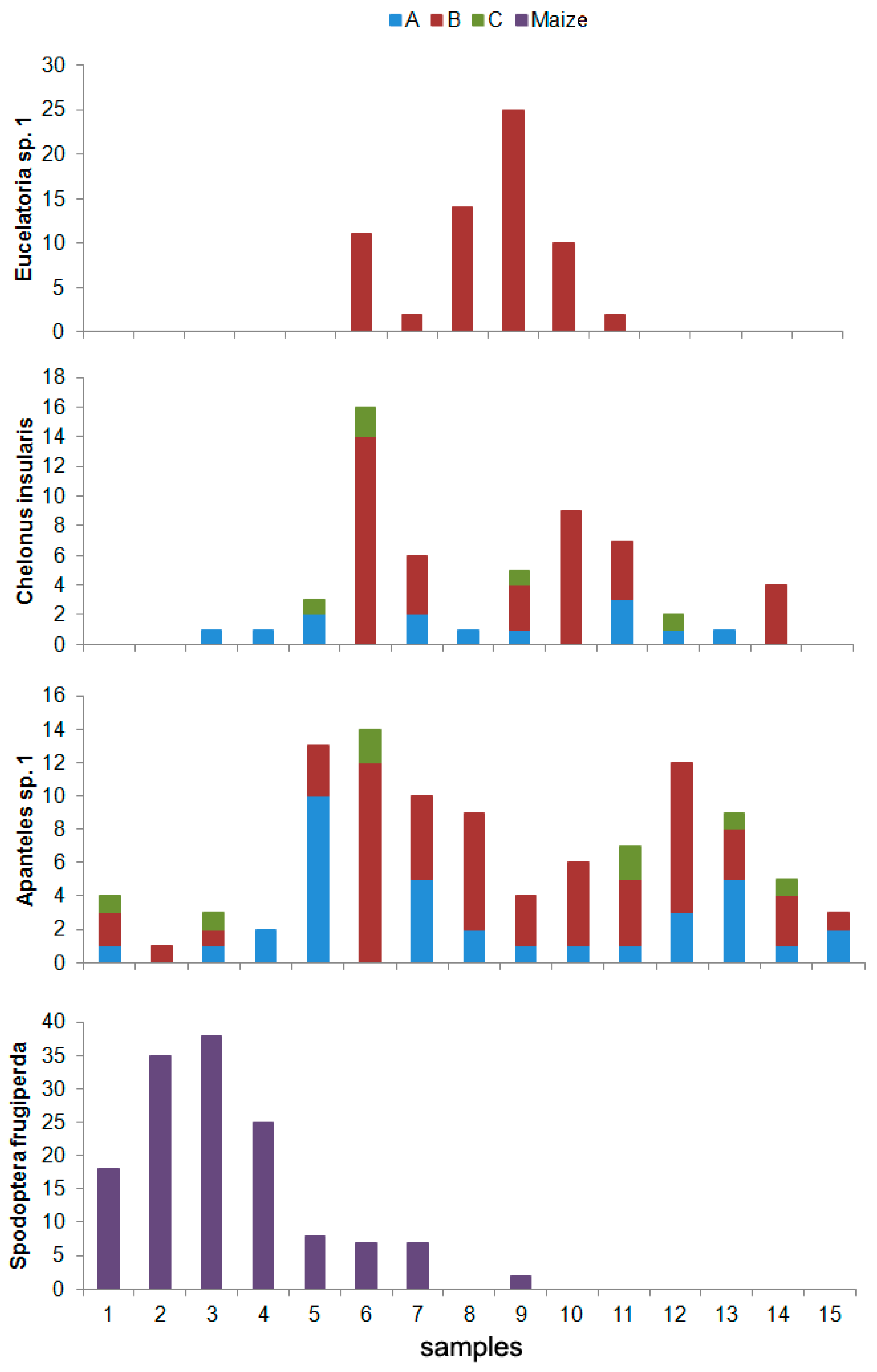
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fiedler, A.K.; Landis, D.A. Attractiveness of michigan native plants to arthropod natural enemies and herbivores. Environ. Entomol. 2007, 36, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Landis, D.; Menalled, F.; Lee, J.; Carmona, D.; Pérez-Valdez, A. Habitat management to enhance biological control in IPM. In Emerging Technologies for Integrated Pest Management: Concepts, Research and Implementation, 1st ed.; Kenedy, G.G., Sutton, T.B., Eds.; APS Press: St. Paul, NC, USA, 2000; pp. 226–239. [Google Scholar]
- Frank van Veen, F.J.; Morris, R.J.; Godfray, H.C.J. Apparent competition, quantitative food webs, and the structure of phytophagous insect communities. Annu. Rev. Entomol. 2006, 51, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.K.; Landis, D.A. Habitat diversification in biological control: The role of plant resources. In Plant-Provided Food for Carnivorous Insects, 1st ed.; Wackers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 305–325. [Google Scholar]
- Géneau, C.E.; Wäckers, F.L.; Luka, H.; Daniel, C.; Balmer, O. Selective flowers to enhance biological control of cabbage pests by parasitoids. Basic. App. Ecol. 2012, 13, 85–93. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Wratten, S.D. Improved fitness of aphid parasitoids receiving resource subsidies. Ecology 2004, 85, 658–666. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I. Agroecología. Teoría y Práctica Para una Agricultura Sustentable, 1st ed.; Programa de las Naciones Unidas para el Medio Ambiente: Mexico, Mexico, 2000. (In Spanish) [Google Scholar]
- Fiedler, A.K.; Landis, D.A. Plant characteristics associated with natural enemy abundance at Michigan native plants. Environ. Entomol. 2007, 36, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Vattala, H.D.; Wratten, S.D.; Phillips, C.B.; Wäckers, F.L. The influence of flower morphology and nectar quality on the longevity of a parasitoid biological control agent. Biol. Control 2006, 39, 179–185. [Google Scholar] [CrossRef]
- Haas, J.; Creamer, W.; Huaman, L.; Goldstein, D.; Reinhard, K.; Rodriguez, C.V. Evidence for maize (Zea mays) in the Late Archaic (3000–1800 B.C.) in the Norte Chico region of Peru. Proc. Natl. Acad. Sci. USA 2013, 110, 4945–4949. [Google Scholar] [CrossRef] [PubMed]
- Gonzáles, W.L.; Gianoli, E.; Niemeyer, H.M. Plant quality vs. risc of parasitism: Within-plant distribution and performance of the corn leaf aphid, Rhopalosiphum maidis. Agric. For. Entomol. 2001, 3, 29–33. [Google Scholar] [CrossRef]
- Casmuz, A.; Juarez, M.L.; Socías, G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastamin, G. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 2010, 69, 209–231. [Google Scholar]
- Ashley, T.R. Geographical distributions and parasitization Levels for parasitoids of the fall armyworm, Spodoptera frugiperda. Fla. Entomol. 1986, 69, 516. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean basin: An inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Degen, T.; Bergvinson, D.; Savidan, A.; Tamo, C.; Turlings, T.C. Occurrence and direct control potential of parasitoids and predators of the fall armyworm (Lepidoptera: Noctuidae) on maize in the subtropical lowlands of Mexico. Agric. For. Entomol. 2004, 6, 83–88. [Google Scholar] [CrossRef]
- Vilaseca, C.J.; Baptiste, L.G.; López-Ávila, A. Incidencia de los márgenes sobre el control biológico natural de Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) en cultivos de arroz. Rev. Corpoica-Cienc. Tecnol. Agropecu. 2008, 9, 45–54. (In Spanish) [Google Scholar] [CrossRef]
- Holdridge, L.R. Life Zone Ecology, 1st ed.; Tropical Science Center: San Jose, Costa Rica, 1967. [Google Scholar]
- Brack, A. Ecología del Perú, 3rd ed.; Bruño: Lima, Peru, 2010. [Google Scholar]
- Lopez, O.; Salto, C.; Luiselli, S. Foeniculum vulgare miller como hospedera de pulgones y sus enemigos naturales en otoño. FAVE Secc. Cienc. Agrar. 2003, 2, 55–65. [Google Scholar] [CrossRef]
- Sánchez, G.; Sarmiento, J. Plagas del Cultivo del Maíz, 1st ed.; Departamento de Entomología y Fitopatología, UNALM: Lima, Peru, 2003. (In Spanish) [Google Scholar]
- Mazón, M.; Bordera, S. Effectiveness of two sampling methods used for collecting Ichneumonidae (Hymenoptera) in the Cabañeros National Park (Spain). Eur. J. Entomol. 2008, 105, 879–888. [Google Scholar] [CrossRef]
- Solow, A.R. A simple test for change in community structure. J. Anim. Ecol. 1993, 62, 191–193. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. 2006. Available online: http://viceroy.eeb.uconn.edu/EstimateS/ (accessed on 1 August 2010).
- Vázquez, L.L.; Matienzo, Y.; Veitía, M.; Alfonso, J. Conservación y Manejo de Enemigos Naturales de Insectos Fitófagos en los Sistemas Agrícolas de Cuba, 1st ed.; Instituto de Investigaciones de Sanidad Vegetal: La Habana, Cuba, 2008. (In Spanish) [Google Scholar]
- Maingay, H.M.; Bugg, R.L.; Carlson, R.W.; Davidson, N.A. Predatory and parasitic wasps (Hymenoptera) feeding at flowers of sweet fennel (Foeniculum vulgare miller var. dulce Battandier & Trabut, Apiaceae) and Spearmint (Mentha spicata L., Lamiaceae) in Massachusetts. Biol. Agric. Hortic. 1991, 7, 363–383. [Google Scholar]
- Weber, M.G.; Porturas, L.D.; Keeler, K.H. World List of Plants with Extrafloral Nectaries. 2015. Available online: www.extrafloralnectaries.org (accessed on 6 May 2017).
- Matienzo, Y.; Veitía, M.; Alayón, G. Las plantas florecidas: Un componente básico para la conservación de atrópodos benéficos en fincas de la agricultura urbana y suburbana. Agricultura Orgánica 2010, 3, 26–28. (In Spanish) [Google Scholar]
- Heil, M.; Hilpert, A.; Krüger, R.; Linsenmair, K.E. Competition among visitors to extrafloral nectaries as a source of ecological costs of an indirect defence. J. Trop. Ecol. 2004, 20, 201–208. [Google Scholar] [CrossRef]
- Fernández, J.L.; Garcés, G.; Portuondo, E.; Valdés, P.; Expósito, I. Insectos asociados con flores de malezas del Jardín Botánico de Santiago de Cuba, con énfasis en Hymenoptera. Rev. Biol. Trop. 2001, 49, 1013–1026. [Google Scholar] [PubMed]
- Amat, I.; Bernard, S.; Foray, V.; Pelosse, P.; Bernstein, C.; Desouhant, E. Fuelling flight in a parasitic wasp: Which energetic substrate to use? Ecol. Entomol. 2012, 37, 480–489. [Google Scholar] [CrossRef]
- Pemberton, R.W.; Lee, J.-H. The influence of extrafloral nectaries on parasitism of an insect herbivore. Am. J. Bot. 1996, 83, 1187–1194. [Google Scholar] [CrossRef]
- Stapel, J.O.; Cortesero, A.-M.; De Moraes, C.M.; Tumlinson, J.H.; Lewis, W.J. Extrafloral nectar, honeydew, and sucrose effects on searching behavior and efficiency of Microplitis croceipes (Hymenoptera: Braconidae) in cotton. Environ. Entomol. 1997, 36, 617–623. [Google Scholar] [CrossRef]
- Faria, C.A.; Wäckers, F.L.; Turlings, T.C.J. The nutritional value of aphid honeydew for non-aphid parasitoids. Basic. Appl. Ecol. 2008, 9, 286–297. [Google Scholar] [CrossRef]
- Heil, M. Extrafloral Nectar at the Plant-Insect Interface: A spotlight on chemical ecology, phenotypic plasticity, and food webs. Annu. Rev. Entomol. 2015, 60, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Wäckers, F.L.; Bonifay, C. How to be sweet? Extrafloral nectar allocation by Gossypium hirsutum fits optimal defense theory predictions. Ecology 2004, 85, 1512–1518. [Google Scholar] [CrossRef]
- Koptur, S.; Jones, I.M.; Peña, J.E. The influence of host plant extrafloral nectaries on multitrophic Interactions: An experimental investigation. PLoS ONE 2015, 10, e0138157. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Weis, J.J.; Forbes, A.E.; Tilmon, K.J.; Ives, A.R. Biodiversity as both a cause and consequence of resource availability: A study of reciprocal causality in a predator-prey system: Bidirectional diversity-resource relationships. J. Anim. Ecol. 2006, 75, 497–505. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Frost, C.J.; Mescher, M.C.; Carlson, J.E.; De Moraes, C.M. Plant defense priming against herbivores: Getting ready for a different battle. Plant Physiol. 2008, 146, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Poelman, E.H.; Van Loon, J.J.A.; Van Dam, N.M.; Vet, L.E.M.; Dicke, M. Herbivore-induced plant responses in Brassica oleracea prevail over effects of constitutive resistance and result in enhanced herbivore attack. Ecol. Entomol. 2010, 35, 240–247. [Google Scholar] [CrossRef]
- De Rijk, M.; Dicke, M.; Poelman, E.H. Foraging behaviour by parasitoids in multiherbivore communities. Anim. Behav. 2013, 85, 1517–1528. [Google Scholar] [CrossRef]
- Li, Y.; Weldegergis, B.T.; Chamontri, S.; Dicke, M.; Gols, R. Does aphid infestation interfere with indirect plant defense against lepidopteran caterpillars in wild cabbage? J. Chem. Ecol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Clissold, F.J.; Sanson, G.D.; Read, J.; Simpson, S.J. Gross vs. net income: How plant toughness affects performance of an insect herbivore. Ecology 2009, 90, 3393–3405. [Google Scholar] [CrossRef] [PubMed]
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Hunter, M.D. Effects of plant quality on the population ecology of parasitoids. Agr. Forest Entomol. 2003, 5, 1–8. [Google Scholar] [CrossRef]
- Bukovinszky, T.; van Veen, F.J.; Jongema, Y.; Dicke, M. Direct and indirect effects of resource quality on food web structure. Science 2008, 319, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, M.C.; Powrie, L.W.; Husted, L.B. Plant diversity consequences of a herbivore-driven bioma switch from Grassland to Nama-Karoo shrub steppe in South Africa. Appl. Veg. Sci. 2012, 15, 14–25. [Google Scholar] [CrossRef]
- Carmona, D.; Lajeunesse, M.J.; Johnson, M.T.J. Plant traits that predict resistance to herbivores. Func. Ecol. 2011, 25, 358–367. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Ann. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.; Cisneros, F.H. Ocurrencia estacional de plagas del maíz en la costa central del Perú y sus enemigos naturales. Rev. Per. Entomol. 1981, 24, 39–54. (In Spanish) [Google Scholar]
- Quispe, R.; Fundación PROINPA, La Paz, Bolivia. Unpublished work. 2011.
- Yu, D.S.; van Achterberg, C.; Horstmann, K. World Ichneumonoidea 2011. Taxapad 2012 Database. Available online: www.taxapad.com (accessed on 6 May 2017).
- Snyder, W.E.; Snyder, G.B.; Finke, D.L.; Straub, C.S. Predator biodiversity strengthens herbivore suppression. Ecol. Lett. 2006, 9, 789–796. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Refuge | Species | Type | Life Form | Foliage Density | Extrafloral Nectaries |
|---|---|---|---|---|---|
| A | Gossypium barbadense (M) | Cultivated | Herb | High | Yes (bracts) |
| Aster sp (As) | Ornamental | Herb | Medium | No | |
| Foeniculum vulgare (Ap) | Aromatic | Herb | Medium | Yes (bracts) | |
| Coriandrum sativum (Ap) | Aromatic | Herb | Medium | No | |
| Lavandula officinalis (L) | Aromatic | Herb | Medium | No | |
| B | Helianthus annus (As) | Cultivated | Herb | Medium | Yes (bracts) |
| Nicandra physaloides (S) | Weed | Shrub | High | No | |
| Salvia officinalis (L) | Aromatic | Woody | Medium | No | |
| Bidens pilosa (As) | Weed | Shrub | Medium | Yes (stem, nodes, bracts) | |
| Artemisia absinthium (As) | Weed | Shrub | Medium | No | |
| C | Malva parviflora (M) | Weed | Herb | High | No |
| Rosmarinus officinalis (L) | Aromatic | Shrub | Medium | No | |
| Phaseolus vulgaris (F) | Cultivated | Herb | Medium | Yes (stipules) | |
| Galinsoga parviflora (As) | Weed | Herb | Medium | No | |
| Sorghum halepense (P) | Cultivated | Herb | Medium | No |
| Herbivores | Parasitoids | Predators | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Refuge | Plant Species | N | S | N | S | N | S | N | S |
| A | Aster sp. | 22 | 9 | 41 | 27 | 142 | 15 | 214 | 54 |
| Coriandrum sativum | 72 | 4 | 75 | 31 | 260 | 17 | 407 | 52 | |
| Foeniculum vulgare | 222 | 5 | 276 | 54 | 901 | 30 | 1403 | 90 | |
| Gossypium barbadense | 671 | 15 | 229 | 52 | 715 | 27 | 1627 | 97 | |
| Lavandula officinalis | 6 | 2 | 54 | 26 | 117 | 4 | 183 | 34 | |
| Total Refuge A | 993 | 21 | 675 | 71 | 2135 | 41 | 3803 | 133 | |
| B | Artemisia absinthium | 1 | 1 | 25 | 17 | 44 | 2 | 70 | 20 |
| Bidens pilosa | 97 | 8 | 390 | 60 | 149 | 23 | 687 | 94 | |
| Helianthus annus | 336 | 13 | 182 | 13 | 429 | 21 | 957 | 50 | |
| Nicandra physaloides | 499 | 10 | 177 | 33 | 165 | 14 | 856 | 59 | |
| Salvia officinalis | 11 | 5 | 25 | 19 | 76 | 9 | 118 | 34 | |
| Total Refuge B | 944 | 17 | 799 | 72 | 863 | 31 | 2606 | 120 | |
| C | Galinsoga parviflora | 23 | 9 | 104 | 26 | 189 | 11 | 316 | 46 |
| Malva parviflora | 376 | 11 | 145 | 41 | 348 | 21 | 878 | 77 | |
| Phaseolus vulgaris | 159 | 7 | 15 | 9 | 90 | 3 | 265 | 20 | |
| Rosmarinus officinalis | 24 | 1 | 17 | 14 | 15 | 4 | 56 | 19 | |
| Sorghum halepense | 43 | 7 | 148 | 24 | 240 | 8 | 431 | 39 | |
| Total Refuge C | 625 | 19 | 429 | 59 | 882 | 27 | 1936 | 105 | |
| Maize | 2138 | 15 | 207 | 5 | 1769 | 18 | 4144 | 38 | |
| Total | 4700 | 38 | 2110 | 89 | 5649 | 52 | 12,459 | 179 | |
| Effectiveness | ||||||
|---|---|---|---|---|---|---|
| Refuge | Plant Species | S obs | S ICE | S Jack1 | ICE | Jack 1 |
| A | Aster sp. | 61 | 132 | 97 | 46.26% | 62.96% |
| Coriandrum sativum | 60 | 98 | 86 | 61.21% | 69.41% | |
| Foeniculum vulgare | 99 | 121 | 123 | 81.71% | 80.74% | |
| Gossypium barbadense | 106 | 142 | 142 | 74.82% | 74.71% | |
| Lavandula officinalis | 36 | 55 | 54 | 65.71% | 66.82% | |
| Total Refuge A | 133 | 150 | 159 | 88.93% | 83.52% | |
| B | Artemisia absinthium | 22 | 74 | 36 | 29.61% | 61.23% |
| Bidens pilosa | 100 | 123 | 128 | 81.06% | 77.98% | |
| Helianthus annus | 57 | 89 | 80 | 64.15% | 70.86% | |
| Nicandra physaloides | 68 | 112 | 100 | 60.92% | 70.15% | |
| Salvia officinalis | 36 | 116 | 58 | 31.05% | 61.76% | |
| Total Refuge B | 120 | 132 | 140 | 90.93% | 85.48% | |
| C | Galinsoga parviflora | 52 | 99 | 77 | 52.64% | 67.17% |
| Malva parviflora | 83 | 123 | 114 | 67.57% | 72.70% | |
| Phaseolus vulgaris | 23 | 36 | 31 | 63.33% | 73.46% | |
| Rosmarinus officinalis | 20 | 64 | 32 | 31.19% | 62.25% | |
| Sorghum halepense | 42 | 65 | 60 | 64.90% | 70.07% | |
| Total Refuge C | 105 | 142 | 138 | 73.94% | 76.05% | |
| Maize | 40 | 46 | 48 | 86.58% | 34.21% | |
| Plant attributes | Functional group | Insect attributes | Refuge A | Refuge B | Refuge C |
|---|---|---|---|---|---|
| Extrafloral nectaries | Parasitoids | Abundance | 25.96 (0.000) | 9.38 (0.002) | 8.98 (0.003) |
| Richness | 23.78 (0.000) | 4.82 (0.028) | 8.06 (0.005) | ||
| Predators | Abundance | 75.72 (0.000) | 11.05 (0.001) | 0.38 (0.539) | |
| Richness | 83.30 (0.000) | 24.98 (0.000) | 0.22 (0.639) | ||
| Foliage density | Parasitoids | Abundance | 5.97 (0.015) | 2.33 (0.127) | 0.28 (0.594) |
| Richness | 5.11 (0.024) | 0.63 (0.427) | 0.63 (0.427) | ||
| Predators | Abundance | 29.29 (0.000) | 1.22 (0.269) | 10.71 (0.001) | |
| Richness | 26.56 (0.000) | 0.03 (0.874) | 6.18 (0.013) | ||
| Life form | Parasitoids | Abundance | - | 13.34 (0.001) | 1.06 (0.304) |
| Richness | - | 16.19 (0.000) | 0.43 (0.514) | ||
| Predators | Abundance | - | 23.71 (0.000) | 34.78 (0.000) | |
| Richness | - | 24.97 (0.000) | 28.98 (0.000) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quispe, R.; Mazón, M.; Rodríguez-Berrío, A. Do Refuge Plants Favour Natural Pest Control in Maize Crops? Insects 2017, 8, 71. https://doi.org/10.3390/insects8030071
Quispe R, Mazón M, Rodríguez-Berrío A. Do Refuge Plants Favour Natural Pest Control in Maize Crops? Insects. 2017; 8(3):71. https://doi.org/10.3390/insects8030071
Chicago/Turabian StyleQuispe, Reinaldo, Marina Mazón, and Alexander Rodríguez-Berrío. 2017. "Do Refuge Plants Favour Natural Pest Control in Maize Crops?" Insects 8, no. 3: 71. https://doi.org/10.3390/insects8030071
APA StyleQuispe, R., Mazón, M., & Rodríguez-Berrío, A. (2017). Do Refuge Plants Favour Natural Pest Control in Maize Crops? Insects, 8(3), 71. https://doi.org/10.3390/insects8030071