
A Geometric Analysis of the Regulation of Inorganic Nutrient Intake by the Subterranean Termite Reticulitermes flavipes Kollar


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collections
2.2. Feeding Trials
2.3. Data Analysis
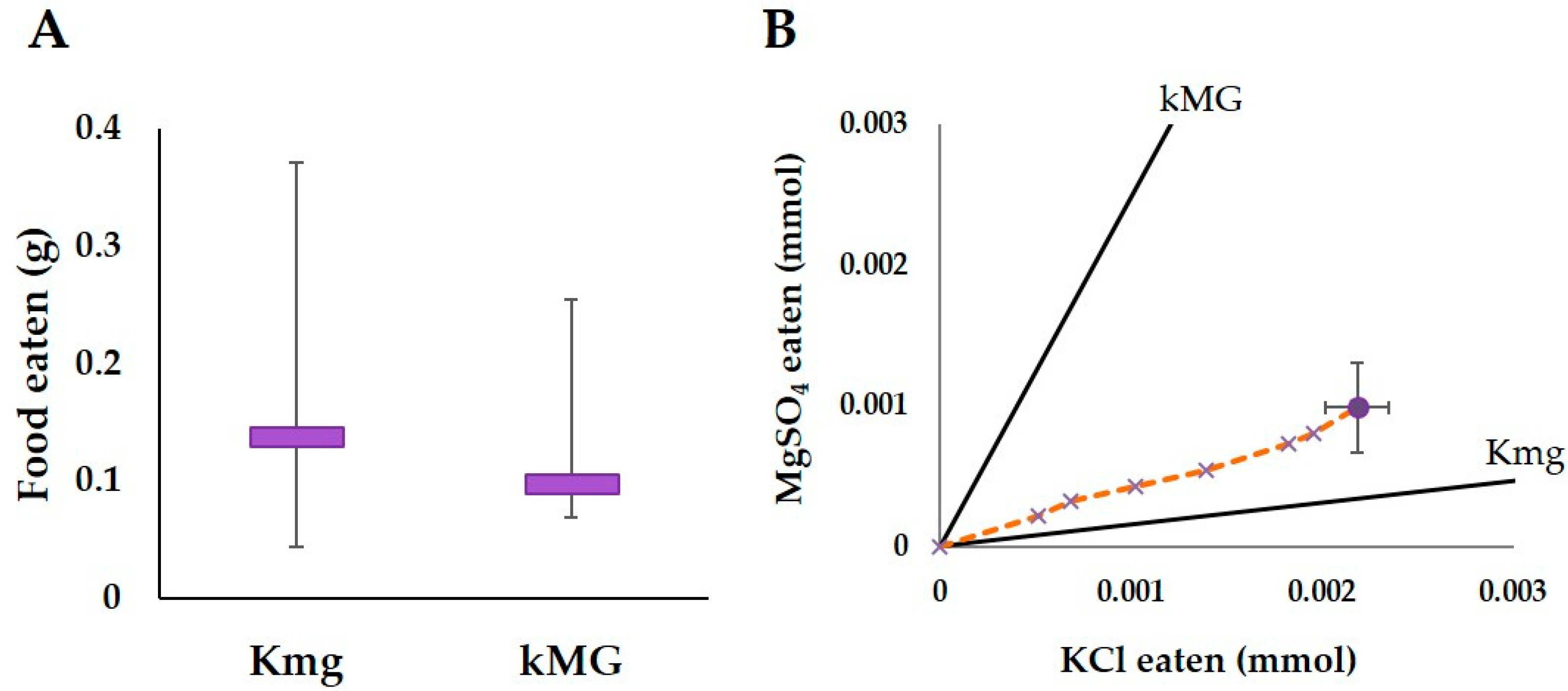
3. Results
3.1. Results of Individual Tests
3.1.1. Kfe vs. kFE Trial
3.1.2. MGfe vs. mgFE Trial
3.1.3. Kmg/kMG Trial
3.2. Combined Analysis and Intake Target
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Seeley, T.D.; Camazine, S.; Sneyd, J. Collective decision-making in honey bees: How colonies choose among nectar sources. Behav. Ecol. Sociobiol. 1991, 28, 277–290. [Google Scholar] [CrossRef]
- Sudd, J.H.; Sudd, M.E. Seasonal changes in the response of wood-ants (Formica lugubris) to sucrose baits. Ecol. Entmol. 1985, 10, 89–97. [Google Scholar] [CrossRef]
- Cook, S.C.; Eubanks, M.D.; Gold, R.E.; Behmer, S.T. Colony-level macronutrient regulation in ants: mechanisms, hoarding and associated costs. Anim. Behav. 2010, 79, 429–437. [Google Scholar] [CrossRef]
- Cook, S.C.; Eubanks, M.D.; Gold, R.E.; Behmer, S.T. Seasonality directs contrasting food collection behavior and nutrient regulation strategies in ants. PLoS ONE 2011, 6, e25407. [Google Scholar] [CrossRef] [PubMed]
- Judd, T.M. Relationship between food stores and foraging behavior of Pheidole ceres (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 2006, 99, 398–406. [Google Scholar] [CrossRef]
- Sorensen, A.A.; Busch, T.M.; Vinson, S.B. Behavioral flexibility of temporal subcastes in the fire ant, Solenopsis invicta in response to food. Psyche 1985, 91, 316–331. [Google Scholar] [CrossRef]
- Dussutour, A.; Simpson, S.J. Carbohydrate regulation in relation to colony growth in ants. J. Exp. Biol. 2008, 211, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.E.; Eggleton, P.; Bignell, D.E. Gut content analysis and a new feeding group classification of termites. Ecol. Entmol. 2001, 26, 356–366. [Google Scholar] [CrossRef]
- Inward, D.J.G.; Vogler, A.P.; Eggleton, P.A. Comprehensive phylogenetic analysis of termites (Isoptera) illuminates key aspects of their evolutionary biology. Mol. Phylogenet. Evol. 2007, 44, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P.; Tayasu, I. Feeding groups, lifetypes and the global ecology of termites. Ecol. Res. 2001, 16, 941–960. [Google Scholar] [CrossRef]
- Reinhard, J.; Kaib, M. Trail communication during foraging and recruitment in the subterranean termite Reticulitermes santonensis De Feytaud (Isoptera, Rhinotermitidae). J. Insect Behav. 2001, 14, 157–171. [Google Scholar] [CrossRef]
- La Fage, J.P.; Nutting, W.L. Nutrient dynamics of termites. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 165–232. [Google Scholar]
- Suárez, M.E.; Thorne, B.L. Rate, amount, and distribution pattern of alimentary fluid transfer via trophallaxis in three species of termites (Isoptera: Rhinotermitidae, Termopsidae). Ann. Entomol. Soc. Am. 2000, 93, 145–155. [Google Scholar] [CrossRef]
- Judd, T.M.; Fasnacht, M.P. Distribution of micronutrients in social insects: A test in the termite Reticulitermes flavipes (Isoptera: Rhinotermitidae) and the ant Myrmica punctiventris (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 2007, 100, 893–899. [Google Scholar] [CrossRef]
- Waller, D.A.; La Fage, J.P. Nutritional ecology of termites. In Nutritional Ecology of Insects, Mites, Spiders and Related Invertebrates; Slansky, F., Jr., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1987; pp. 487–532. [Google Scholar]
- Abushama, F.T.; Kamal, M.A. The role of sugars in the food-selection of termite Microtermes traegardhi (Sjostedt). J. Appl. Entomol. 1977, 84, 250–255. [Google Scholar]
- Botch, P.S.; Brennan, C.L.; Judd, T.M. Seasonal effects of calcium and phosphate on the feeding preference of the termite Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2010, 55, 42–56. [Google Scholar]
- Haifig, I.; Costa-Leonardo, A.M.; Marchetti, F.F. Effects of nutrients on feeding activities of the pest termite Heterotermes tenuis (Isoptera: Rhinotermitidae). J. Appl. Entomol. 2008, 132, 497–501. [Google Scholar] [CrossRef]
- Haifig, I.; Marchetti, F.F.; Costa-Leonardo, A.M. Nutrients affecting food choice by the pest subterranean termite Coptotermes gestroi (Isoptera: Rhinotermitidae). Int. J. Pest Manag. 2010, 56, 371–375. [Google Scholar] [CrossRef]
- Saran, R.K.; Rust, M.K. Feeding, uptake, and utilization of carbohydrates by western subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2005, 98, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Swoboda, L.E.; Miller, D.M.; Fell, R.J.; Mullins, D.E. The effect of nutrient compounds (sugars and amino-acids) on bait consumption by Reticulitermes spp. (Isoptera: Rhinotermitidae). Sociobiology 2004, 44, 547–563. [Google Scholar]
- Waller, D.A.; Morlino, S.E.; Matkins, N. Factors affecting termite recruitment to baits in laboratory and field studies. In Proceedings of the 3rd International Conference on Urban Pests, Czech University of Agriculture, Prague, Czech Republic, 19–22 July 1999; Robinson, W.M., Rettich, F., Rambo, G.W., Eds.; Grafické Závody Hronov: Prague, Czech Republic, 1999; pp. 597–600. [Google Scholar]
- Castillo, V.P.; Sajap, A.S.; Sahri, M.H. Feeding response of subterranean termites Coptotermes curvignathus and Coptotermes gestroi (Blattodea: Rhinotermitidae) to baits supplemented with sugars, amino acids, and cassava. J. Econ. Entomol. 2013, 106, 1794–1801. [Google Scholar] [CrossRef] [PubMed]
- Haifig, I.; Jost, C.; Fourcassie, V.; Zana, Y.; Costa-Leonardo, A. Dynamics of foraging trails in the Neotropical termite Velocitermes heteropterus (Isoptera: Termitidae). Behav. Proc. 2015, 118, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wallace, B.A.; Judd, T.M. A test of seasonal responses to sugars in four populations of the termite Reticulitermes flavipes. J. Econ. Entomol. 2010, 103, 2126–2131. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. A multi-level analysis of feeding behaviour: The geometry of nutritional decisions. Philos. Trans. R. Soc. B 1993, 342, 381–402. [Google Scholar] [CrossRef]
- Simpson, S.J.; Raubenheimer, D. The geometric analysis of feeding and nutrition: A user’s guide. J. Insect Physiol. 1995, 41, 545–553. [Google Scholar] [CrossRef]
- Behmer, S.T.; Raubenheimer, D.; Simpson, S.J. Frequency-dependent food selection in locusts: a geometric analysis of the role of nutrient balancing. Anim. Behav. 2001, 61, 995. [Google Scholar] [CrossRef]
- Hewson-Hughes, A.K.; Hewson-Hughes, V.L.; Miller, A.T.; Hall, S.R.; Simpson, S.J.; Raubenheimer, D. Geometric analysis of macronutrient selection in the adult domestic cat, Felis catus. J. Exp. Biol. 2011, 214, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Hewson-Hughes, A.K.; Hewson-Hughes, V.L.; Coyler, A.; Miller, A.T.; McGrane, S.J.; Hall, S.R.; Butterwick, R.F.; Simpson, S.J.; Raubenheimer, D. Geometric analysis of macronutrient selection in breeds of the domestic dog, Canis lupus familiaris. Behav. Ecol. 2013, 24, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Hancock, D.P.; Gosby, A.K.; McMahon, A.C.; Solom, S.M.C.; Le Couteur, D.G.; Conigrave, A.D.; Raubenheimer, D.; Simpson, S.J. Effects of dietary protein to carbohydrate balance on energy intake, fat storage, and heat production in mice. Obesity 2013, 21, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.; Mayntz, D.; Toft, S.; Clissold, F.J.; Hunt, J.; Raubenheimer, D.; Simpson, S.J. Optimal foraging for specific nutrients in predatory beetles. Proc. Biol. Sci. 2012, 279, 2212–2218. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Behmer, S.T.; Simpson, S.J.; Raubenheimer, D. A geometric analysis of nutrient regulation in the generalist caterpillar Spodoptera littoralis (Boisduval). J. Insect Physiol. 2002, 48, 655–665. [Google Scholar] [CrossRef]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef] [PubMed]
- König, H.; Li, L.; Fröhlich, J. The cellulolytic system of the termite gut. Appl. Microbiol. Biotechnol. 2013, 97, 7943–7962. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Bentley, B.L. Nitrogen fixation in termites: Fate of newly fixed nitrogen. J. Insect Physiol. 1984, 30, 653–655. [Google Scholar] [CrossRef]
- Breznak, J.A.; Brill, W.J.; Mertins, J.W.; Coppel, H.C. Nitrogen fixation in termites. Nature 1973, 244, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Curtis, A.D.; Waller, D.A. Seasonal patterns of nitrogen fixation in termites. Funct. Ecol. 1998, 12, 803–807. [Google Scholar] [CrossRef]
- Ohkuma, M.; Noda, S.; Kudo, T. Phylogenetic diversity of nitrogen fixation genes in the symbiotic microbial community in the gut of diverse termites. Appl. Environ. Microbiol. 1999, 65, 4926–4934. [Google Scholar] [PubMed]
- Prestwich, G.D.; Bentley, B.L.; Carpenter, E.J. Nitrogen sources for neotropical nasute termites: Fixation and selective foraging. Oecologia 1980, 46, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Janzow, M.P.; Judd, T.M. The termite Reticulitermes flavipes (Rhinotermitidae: Isoptera) can acquire micronutrients from soil. Environ. Entomol. 2015, 44, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.C. Insect Diets Science and Technology; CRC Press: New York, NY, USA, 2004. [Google Scholar]
- Nation, J.L. Insect Physiology and Biochemistry; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Botch, P.S.; Judd, T.M. The effects of soil cations on the foraging behavior of the termite Reticulitermes flavipes. J. Econ. Entomol. 2011, 104, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Brauman, A.; Koenig, J.F.; Dutreix, J.; Garcia, J.L. Characterization of two sulfate-reducing bacteria from the gut of the soil-feeding termite, Cubitermes speciosus. Antonie Van Leeuwenhoek 1990, 58, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Droge, S.; Limper, U.; Emtiazi, F.; Schonig, I.; Pavlus, N.; Dryzga, O.; Fischer, U.; Konig, H. In vitro and in vivo sulfate reduction in the gut contents of the termite Mastotermes darwiniensis and the rose-chafer Pachnoda marginata. J. Gen. Appl. Microbiol. 2005, 51, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Kuhnigk, T.; Branke, J.; Krekeler, D.; Cypionka, H.; König, H. A feasible role of sulfate-reducing bacteria in the termite gut. Syst. Appl. Microbiol. 1996, 19, 139–149. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Nichol, H.; Law, J.H.; Winzerling, J.J. Iron metabolism in insects. Annu. Rev. Entomol. 2002, 47, 535–559. [Google Scholar] [CrossRef] [PubMed]
- Judd, T.M.; Corbin, C.C. Effect of cellulose concentration on the feeding preferences of the termite Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2009, 53, 775–784. [Google Scholar]
- Zachariah, N.; Das, A.; Murthy, T.G.; Borges, R.M. Building mud castles: A perspective from brick-laying termites. Sci. Rep. 2017, 7, 4692. [Google Scholar] [CrossRef] [PubMed]
- Daniel, W.W. Applied Nonparametric Statistics, 2nd ed.; PWS-Kent: Boston, MA, USA, 1990. [Google Scholar]
- Emmans, G.C. Diet selection by animals: Theory and experimental design. Proc. Nutr. Soc. 1991, 50, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Jungreis, A.M. Distribution of magnesium in tissues of silkmoth Hyalophora cecropia. Am. J. Physiol. 1973, 224, 27–30. [Google Scholar]
- Kiceniuk, J.; Phillips, J.E. Magnesium regulation in mosquito larvae (Aedes compestris) living in waters of high MgSO4 content. J. Exp. Biol. 1974, 61, 749–760. [Google Scholar] [PubMed]
- Oyarzun, S.E.; Crawshaw, G.J.; Valdes, E.V. Nutrition of the tamandua: I. Nutrient composition of termites (Nasutitermes spp.) and stomach contents from wild tamanduas (Tamandua tetradactyla). Zoo Biol. 1996, 15, 509–524. [Google Scholar] [CrossRef]
- Blumenthal, E.M. Regulation of chloride permeability by endogenously produced tyramine in the Drosophila Malpighian tubule. Am. J. Physiol. 2003, 284, C718–C728. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.R.; Cioffi, M.; Dow, J.A.; Wolfersberger, M.G. Potassium ion transport ATPase in insect epithelia. J. Exp. Biol. 1983, 106, 91–117. [Google Scholar] [PubMed]
- Popham, H.J.; Sun, R.; Shelby, K.S.; Robertson, J.D. Changes in trace metals in hemolymph of baculovirus-infected noctuid larvae. Biol. Trace Elem. Res. 2012, 146, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Inoue, J.I.; Saita, K.; Kudo, T.; Ui, S.; Ohkuma, M. Hydrogen production by termite gut protists: Characterization of iron hydrogenases of parabasalian symbionts of the termite Coptotermes formosanus. Eukaryot. Cell 2007, 6, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Arms, K.; Feeny, P.; Lederhouse, R.C. Sodium: Stimulus for puddling behavior by tiger swallowtail butterflies, Papilio glaucus. Science 1974, 185, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Barrows, E.M. Aggregation behavior and response to sodium chloride in females of a solitary bee, Augochlora pura (Hymenoptera: Halictidae). Fla. Entomol. 1974, 57, 189–193. [Google Scholar] [CrossRef]
- Pivnick, K.A.; McNeil, J.M. Puddling in butterflies: Sodium affects reproductive success in Thymelicus lineola. Physiol. Entomol. 1987, 12, 461–472. [Google Scholar] [CrossRef]
- Spring, J.H.; Hyatt, A.D.; Marshall, A.T. Uptake and release of sodium and potassium by the fat body of the American cockroach in vitro. J. Insect Physiol. 1986, 32, 439–444. [Google Scholar] [CrossRef]
- Bonoan, R.E.; Tai, T.M.; Tagle Rodriguez, M.; Feller, L.; Daddario, S.R.; Czaja, R.A.; O’Connor, L.D.; Burruess, G.; Starks, P.T. Seasonality of salt foraging in honey bees (Apis mellifera). Ecol. Entomol. 2017, 42, 195–201. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Type | KCl mg (mM) | MgSO4 mg (mM) | FePO4 mg (mM) |
|---|---|---|---|
| kFE | 0.025 (3.35 × 10−3) | 0.050 (4.15 × 10−3) | 0.100 (6.63 × 10−3) |
| kMG | 0.025 (3.35 × 10−3) | 0.100 (8.31 × 10−3) | 0.050 (3.32 × 10−3) |
| mgFE | 0.050 (6.71 × 10−3) | 0.025 (2.08 × 10−3) | 0.100 (6.63 × 10−3) |
| Mgfe | 0.050 (6.71 × 10−3) | 0.100 (8.31 × 10−3) | 0.025 (1.66 × 10−3) |
| Kmg | 0.100 (13.4 × 10−2) | 0.025 (2.08 × 10−3) | 0.050 (3.32 × 10−3) |
| Kfe | 0.100 (13.4 × 10−2) | 0.050 (4.15 × 10−3) | 0.025 (1.66 × 10−3) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Judd, T.M.; Landes, J.R.; Ohara, H.; Riley, A.W. A Geometric Analysis of the Regulation of Inorganic Nutrient Intake by the Subterranean Termite Reticulitermes flavipes Kollar. Insects 2017, 8, 97. https://doi.org/10.3390/insects8030097
Judd TM, Landes JR, Ohara H, Riley AW. A Geometric Analysis of the Regulation of Inorganic Nutrient Intake by the Subterranean Termite Reticulitermes flavipes Kollar. Insects. 2017; 8(3):97. https://doi.org/10.3390/insects8030097
Chicago/Turabian StyleJudd, Timothy M., James R. Landes, Haruna Ohara, and Alex W. Riley. 2017. "A Geometric Analysis of the Regulation of Inorganic Nutrient Intake by the Subterranean Termite Reticulitermes flavipes Kollar" Insects 8, no. 3: 97. https://doi.org/10.3390/insects8030097
APA StyleJudd, T. M., Landes, J. R., Ohara, H., & Riley, A. W. (2017). A Geometric Analysis of the Regulation of Inorganic Nutrient Intake by the Subterranean Termite Reticulitermes flavipes Kollar. Insects, 8(3), 97. https://doi.org/10.3390/insects8030097