Cross-Resistance: A Consequence of Bi-partite Host-Parasite Coevolution


Abstract
:1. Introduction
2. Materials and Methods
2.1. Host
2.2. Parasites
2.3. Survival Assays
2.4. Host Treatment Prior to Investigating Gene Expression
2.5. RNA Extraction and cDNA Synthesis
2.6. Candidate Genes and Primer Validation
2.7. Gene Expression Using Quantitative PCR
2.8. Statistical Analysis
2.9. Survival Analysis
2.10. Gene Expression Analysis
3. Results
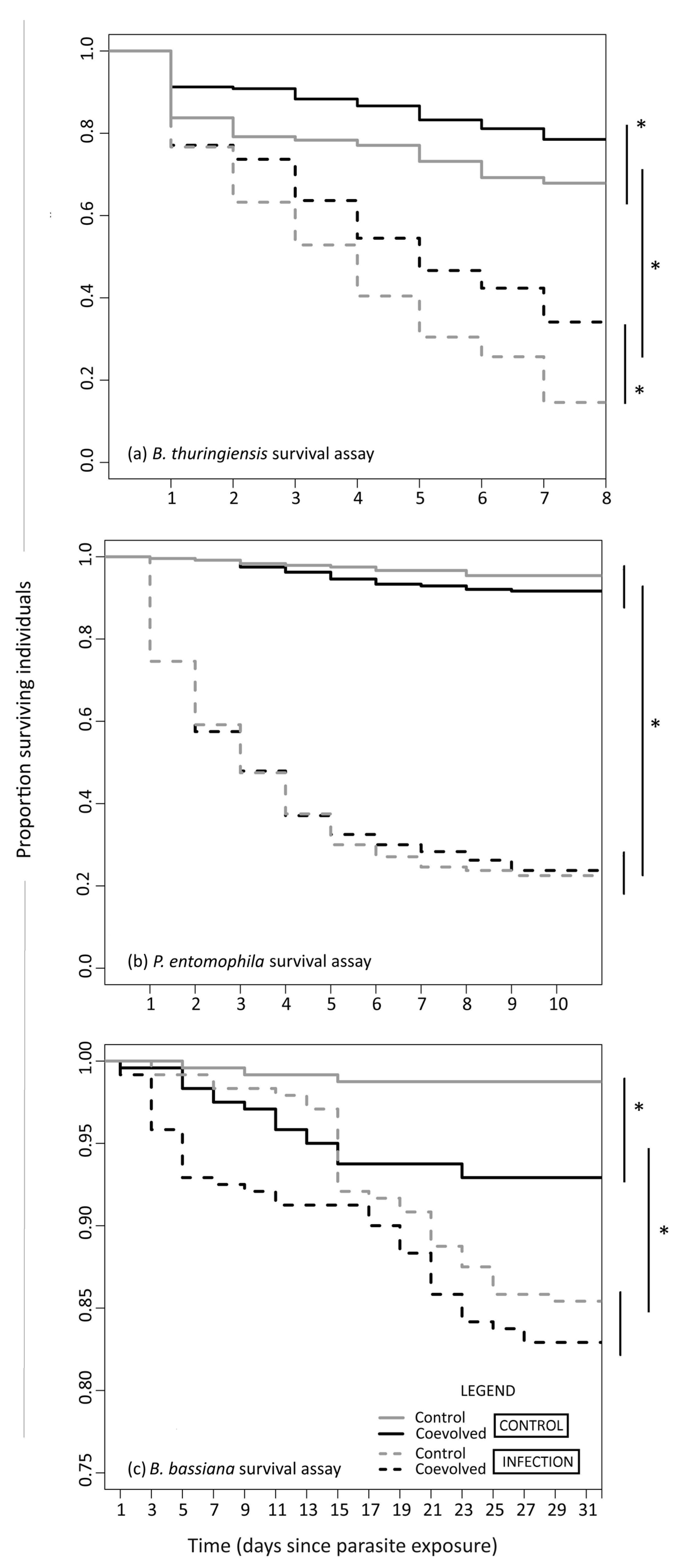
3.1. B. bassiana Coevolved Beetles Are Positively Cross-Resistant to B. thuringiensis
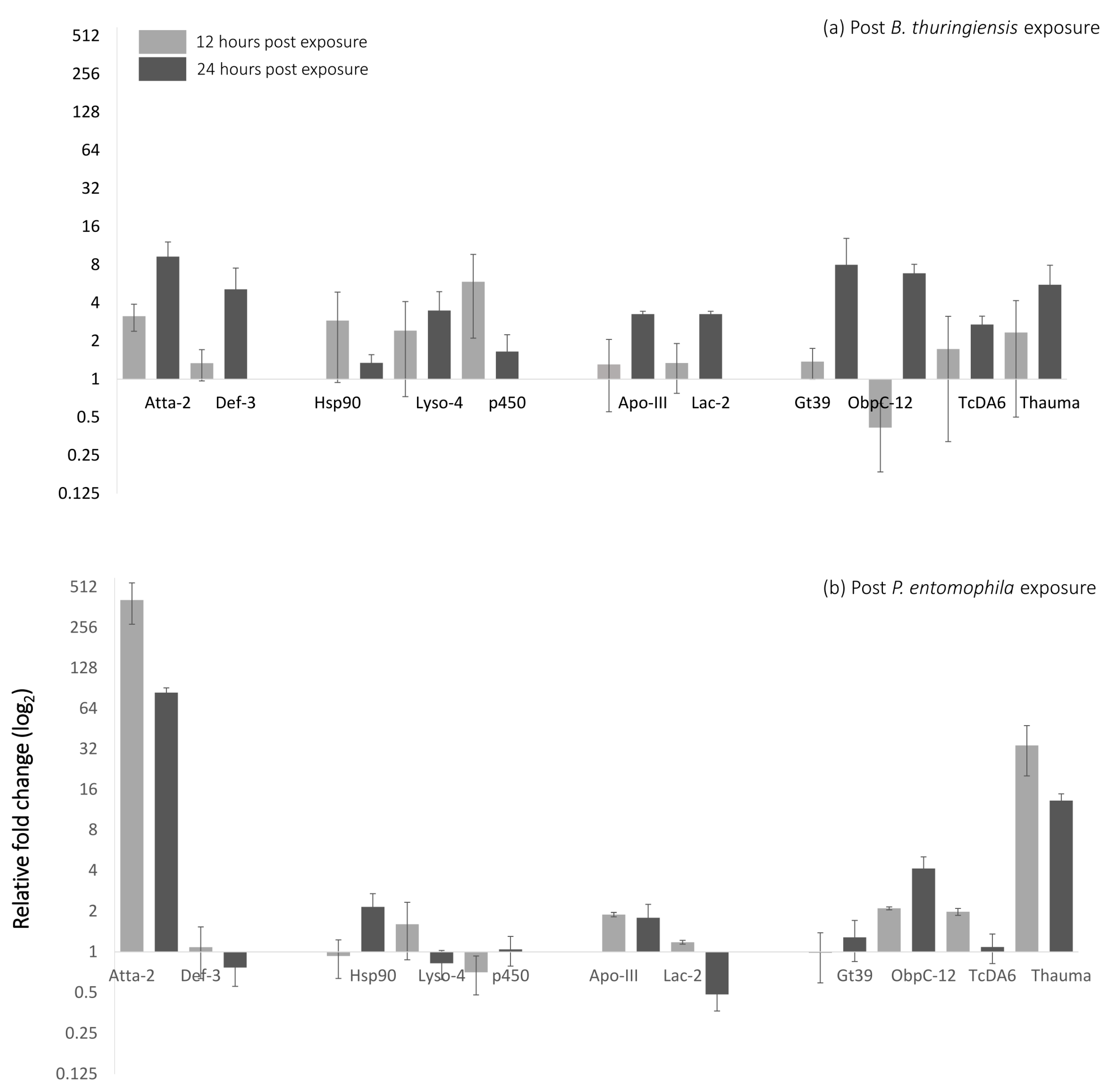
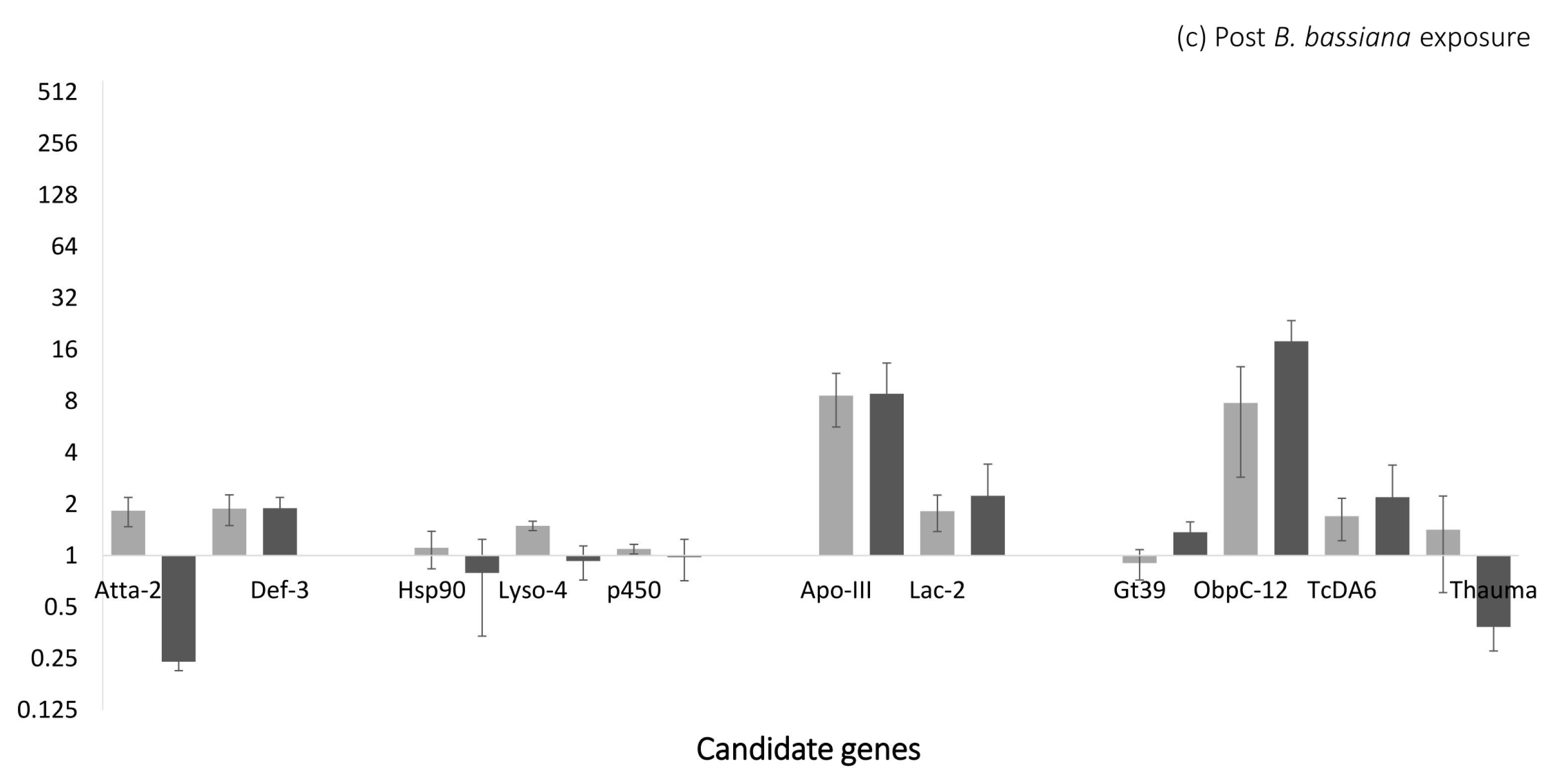
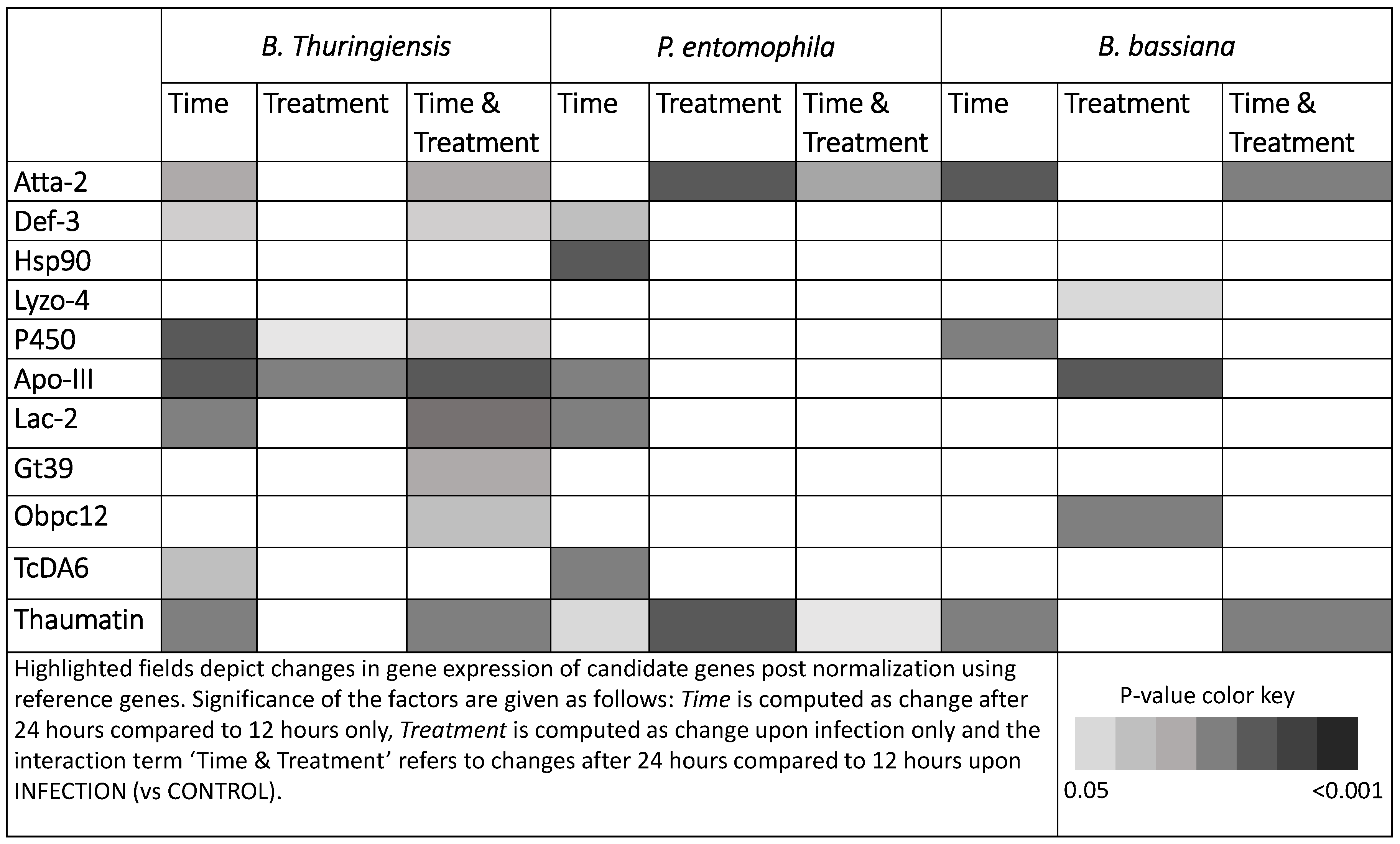
3.2. Gene Expression Analysis Reveals Expression of Markers for Oral Toxicity upon B. bassiana Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bargielowski, I.; Koella, J.C. A Possible Mechanism for the Suppression of Plasmodium berghei Development in the Mosquito Anopheles gambiae by the Microsporidian Vavraia culicis. PLoS ONE 2009, 4, e4676. [Google Scholar] [CrossRef] [PubMed]
- Betts, A.; Rafaluk, C.; King, K.C. Host and Parasite Evolution in a Tangled Bank. Trends Parasitol. 2016, 32, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Hafer, N.; Milinski, M. Inter- and intraspecific conflicts between parasites over host manipulation. Proc. R. Soc. B 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Bordes, F.; Morand, S. Coevolution between multiple helminth infestations and basal immune investment in mammals: Cumulative effects of polyparasitism? Parasitol. Res. 2009, 106, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Start, D.; Gilbert, B. Host–parasitoid evolution in a metacommunity. Proc. R. Soc. B 2016, 283, 20160477. [Google Scholar] [CrossRef] [PubMed]
- De Roode, J.C.; Culleton, R.; Cheesman, S.J.; Carter, R.; Read, A.F. Host heterogeneity is a determinant of competitive exclusion or coexistence in genetically diverse malaria infections. Proc. Biol. Sci. 2004, 271, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, M.J.; Dick, J.T.; Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Front. Ecol. Environ. 2012, 10, 186–194. [Google Scholar] [CrossRef]
- Von Beeren, C.; Maruyama, M.; Hashim, R.; Witte, V. Differential host defense against multiple parasites in ants. Evol. Ecol. 2010, 25, 259–276. [Google Scholar] [CrossRef]
- Fellowes, M.D.E.; Kraaijeveld, A.R.; Godfray, H.C.J. Cross-Resistance Following Artificial Selection for Increased Defense against Parasitoids in Drosophila melanogaster. Evolution 1999, 53, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.E.; Faria, V.G.; Teixeira, L.; Magalhães, S.; Sucena, É. Host Adaptation Is Contingent upon the Infection Route Taken by Pathogens. PLoS Pathog. 2013, 9, e1003601. [Google Scholar] [CrossRef] [PubMed]
- Dubovskiy, I.M.; Whitten, M.M.A.; Yaroslavtseva, O.N.; Greig, C.; Kryukov, V.Y.; Grizanova, E.V.; Mukherjee, K.; Vilcinskas, A.; Glupov, V.V.; Butt, T.M. Can Insects Develop Resistance to Insect Pathogenic Fungi? PLoS ONE 2013, 8, e60248. [Google Scholar] [CrossRef] [PubMed]
- Faria, V.G.; Martins, N.E.; Paulo, T.; Teixeira, L.; Sucena, É.; Magalhães, S. Evolution of Drosophila resistance against different pathogens and infection routes entails no detectable maintenance costs. Evolution 2015, 69, 2799–2809. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.E.; Faria, V.G.; Nolte, V.; Schlötterer, C.; Teixeira, L.; Sucena, É.; Magalhães, S. Host adaptation to viruses relies on few genes with different cross-resistance properties. Proc. Natl. Acad. Sci. USA 2014, 111, 5938–5943. [Google Scholar] [CrossRef] [PubMed]
- Decaestecker, E.; Gaba, S.; Raeymaekers, J.A.M.; Stoks, R.; Van Kerckhoven, L.; Ebert, D.; De Meester, L. Host–parasite ‘Red Queen’ dynamics archived in pond sediment. Nature 2007, 450, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Schulte, R.D.; Makus, C.; Hasert, B.; Michiels, N.K.; Schulenburg, H. Host–parasite local adaptation after experimental coevolution of Caenorhabditis elegans and its microparasite Bacillus thuringiensis. Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 2832–2839. [Google Scholar] [CrossRef] [PubMed]
- Schulte, R.D.; Makus, C.; Hasert, B.; Michiels, N.K.; Schulenburg, H. Multiple reciprocal adaptations and rapid genetic change upon experimental coevolution of an animal host and its microbial parasite. Proc. Natl. Acad. Sci. USA 2010, 107, 7359–7364. [Google Scholar] [CrossRef] [PubMed]
- Bérénos, C.; Schmid-Hempel, P.; Wegner, K.M. Complex adaptive responses during antagonistic coevolution between Tribolium castaneum and its natural parasite Nosema whitei revealed by multiple fitness components. BMC Evol. Biol. 2012, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bérénos, C.; Schmid-Hempel, P.; Mathias Wegner, K. Evolution of host resistance and trade-offs between virulence and transmission potential in an obligately killing parasite. J. Evol. Biol. 2009, 22, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Bérénos, C.; Wegner, K.M.; Schmid-Hempel, P. Antagonistic coevolution with parasites maintains host genetic diversity: An experimental test. Proc. R. Soc. Lond. B Biol. Sci. 2010. [Google Scholar] [CrossRef] [Green Version]
- Dupas, S.; Carton, Y.; Poiriè, M. Genetic dimension of the coevolution of virulence–resistance in Drosophila––Parasitoid wasp relationships. Heredity 2003, 90, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Kerstes, N.A.; Bérénos, C.; Schmid-Hempel, P.; Wegner, K.M. Antagonistic experimental coevolution with a parasite increases host recombination frequency. BMC Evol. Biol. 2012, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraaijeveld, A.R.; Godfray, H.C.J. Trade-off between parasitoid resistance and larval competitive ability in Drosophila melanogaster. Nature 1997, 389, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Kraaijeveld, A.R.; Layen, S.J.; Futerman, P.H.; Godfray, H.C.J. Lack of Phenotypic and Evolutionary Cross-Resistance against Parasitoids and Pathogens in Drosophila melanogaster. PLoS ONE 2012, 7, e53002. [Google Scholar] [CrossRef] [PubMed]
- Zuk, M.; Stoehr, A.M. Immune Defense and Host Life History. Am. Nat. 2002, 160, S9–S22. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Lin, D.M.; Buckling, A.; Thompson, J.N. The costs of evolving resistance in heterogeneous parasite environments. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C.J. Aphid Protected from Pathogen by Endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.J.; Spragg, C.J.; Altincicek, B.; Gerardo, N.M. Symbiont-Mediated Protection against Fungal Pathogens in Pea Aphids: A Role for Pathogen Specificity? Appl. Environ. Microbiol. 2013, 79, 2455–2458. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Butler, S.; Sanchez, G.; Mateos, M. Male killing Spiroplasma protects Drosophila melanogaster against two parasitoid wasps. Heredity 2014, 112, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Bentz, M.L.; Humphrey, E.A.; Harshman, L.G.; Wayne, M.L. Sigma Virus (DMelSV) Incidence in Lines of Drosophila melanogaster Selected for Survival following Infection with Bacillus cereus. Psyche J. Entomol. 2017, 2017, e3593509. [Google Scholar] [CrossRef]
- Behrens, S.; Peuß, R.; Milutinović, B.; Eggert, H.; Esser, D.; Rosenstiel, P.; Schulenburg, H.; Bornberg-Bauer, E.; Kurtz, J. Infection routes matter in population-specific responses of the red flour beetle to the entomopathogen Bacillus thuringiensis. BMC Genomics 2014, 15, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockhurst, M.A.; Koskella, B. Experimental coevolution of species interactions. Trends Ecol. Evol. 2013, 28, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Rafaluk, C.; Yang, W.; Mitschke, A.; Rosenstiel, P.; Schulenburg, H.; Joop, G. Highly potent host external immunity acts as a strong selective force enhancing rapid parasite virulence evolution. Environ. Microbiol. 2017, 19, 2090–2100. [Google Scholar] [CrossRef] [PubMed]
- Rafaluk-Mohr, C.; Wagner, S.; Joop, G. Cryptic changes in immune response and fitness in Tribolium castaneum as a consequence of coevolution with Beauveria bassiana. J. Invertebr. Pathol. 2018, 152, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vodovar, N.; Vallenet, D.; Cruveiller, S.; Rouy, Z.; Barbe, V.; Acosta, C.; Cattolico, L.; Jubin, C.; Lajus, A.; Segurens, B.; et al. Complete genome sequence of the entomopathogenic and metabolically versatile soil bacterium Pseudomonas entomophila. Nat. Biotechnol. 2006, 24, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Milutinović, B.; Stolpe, C.; Peuβ, R.; Armitage, S.A.O.; Kurtz, J. The Red Flour Beetle as a Model for Bacterial Oral Infections. PLoS ONE 2013, 8, e64638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, O.; Joop, G.; Eggert, H.; Hilbert, J.; Daniel, J.; Schmid-Hempel, P.; Kurtz, J. Paternally derived immune priming for offspring in the red flour beetle, Tribolium castaneum. J. Anim. Ecol. 2010, 79, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Roth, O.; Sadd, B.M.; Schmid-Hempel, P.; Kurtz, J. Strain-specific priming of resistance in the red flour beetle, Tribolium castaneum. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Altincicek, B.; Knorr, E.; Vilcinskas, A. Beetle immunity: Identification of immune-inducible genes from the model insect Tribolium castaneum. Dev. Comp. Immunol. 2008, 32, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Muthukrishnan, S.; Beeman, R.W.; Kanost, M.R.; Kramer, K.J. Laccase 2 is the phenoloxidase gene required for beetle cuticle tanning. Proc. Natl. Acad. Sci. USA 2005, 102, 11337–11342. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Dixit, R.; Begum, K.; Park, Y.; Specht, C.A.; Merzendorfer, H.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Analysis of functions of the chitin deacetylase gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2009, 39, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.; Rausell, C.; Real, M.D. Tribolium castaneum Apolipophorin-III acts as an immune response protein against Bacillus thuringiensis Cry3Ba toxic activity. J. Invertebr. Pathol. 2013, 113, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lehmann, S.; Weißbecker, B.; Ojeda Naharros, I.; Schütz, S.; Joop, G.; Wimmer, E.A. Odoriferous Defensive stink gland transcriptome to identify novel genes necessary for quinone synthesis in the red flour beetle, Tribolium castaneum. PLoS Genet. 2013, 9, e1003596. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.C.; Hartzer, K.; Toutges, M.; Oppert, B. Evaluation of quantitative PCR reference genes for gene expression studies in Tribolium castaneum after fungal challenge. J. Microbiol. Methods 2010, 80, 219–221. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; ISBN 3-900051-07-0. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in S. R Package Version 2.37-4. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 1 May 2016).
- Matz, M.V.; Wright, R.M.; Scott, J.G. No Control Genes Required: Bayesian Analysis of qRT-PCR Data. PLoS ONE 2013, 8, e71448. [Google Scholar] [CrossRef] [PubMed]
- Matz, M.V. MCMC.qpcr: Bayesian Analysis of qRT-PCR Data. 2016. Available online: https://cran.r-project.org/web/packages/MCMC.qpcr/index.html (accessed on 14 November 2016).
- Altincicek, B.; Elashry, A.; Guz, N.; Grundler, F.M.W.; Vilcinskas, A.; Dehne, H.-W. Next Generation Sequencing Based Transcriptome Analysis of Septic-Injury Responsive Genes in the Beetle Tribolium castaneum. PLoS ONE 2013, 8, e52004. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Ryu, J.-H.; Han, S.-J.; Choi, K.-H.; Nam, K.-B.; Jang, I.-H.; Lemaitre, B.; Brey, P.T.; Lee, W.-J. Gram-negative Bacteria-binding Protein, a Pattern Recognition Receptor for Lipopolysaccharide and β-1,3-Glucan That Mediates the Signaling for the Induction of Innate Immune Genes in Drosophila melanogaster Cells. J. Biol. Chem. 2000, 275, 32721–32727. [Google Scholar] [CrossRef] [PubMed]
- Gottar, M.; Gobert, V.; Michel, T.; Belvin, M.; Duyk, G.; Hoffmann, J.A.; Ferrandon, D.; Royet, J. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 2002, 416, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.; Rausell, C.; Real, M.D. Proteome Response of Tribolium castaneum Larvae to Bacillus thuringiensis Toxin Producing Strains. PLoS ONE 2013, 8, e55330. [Google Scholar] [CrossRef]
- Pekrul, S.; Grula, E.A. Mode of infection of the corn earworm (Heliothis zea) by Beauveria bassiana as revealed by scanning electron microscopy. J. Invertebr. Pathol. 1979, 34, 238–247. [Google Scholar] [CrossRef]
- Siebeneicher, S.R.; BradleighˆVinson, S.; Kenerley, C.M. Infection of the red imported fire ant by Beauveria bassiana through various routes of exposure. J. Invertebr. Pathol. 1992, 59, 280–285. [Google Scholar] [CrossRef]
- Reber, A.; Chapuisat, M. No Evidence for Immune Priming in Ants Exposed to a Fungal Pathogen. PLoS ONE 2012, 7, e35372. [Google Scholar] [CrossRef] [PubMed]
- Knorr, E.; Schmidtberg, H.; Arslan, D.; Bingsohn, L.; Vilcinskas, A. Translocation of bacteria from the gut to the eggs triggers maternal transgenerational immune priming in Tribolium castaneum. Biol. Lett. 2015, 11, 20150885. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.C.; Cogburn, R.R. Activity of the Entomopathogenic Fungus Beauveria bassiana (Deuteromycota: Hyphomycetes) Against Three Coleopteran Pests of Stored Grain. J. Econ. Entomol. 1999, 92, 691–694. [Google Scholar] [CrossRef]
- Yanagita, T. Studies on oral infection of larvae of the silkworm, Bombyx mori, with Beauveria bassiana. J. Sericultural Sci. Jpn. 1987, 56, 279–284. [Google Scholar] [CrossRef]
- Xiao, G.; Ying, S.-H.; Zheng, P.; Wang, Z.-L.; Zhang, S.; Xie, X.-Q.; Shang, Y.; St. Leger, R.J.; Zhao, G.-P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Kraaijeveld, A.R.; Godfray, H.C.J. Selection for resistance to a fungal pathogen in Drosophila melanogaster. Heredity 2008, 100, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect Phiathogens as Biological Control Agents: Do They Have a Future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z. Naturforschung C 2017, 72, 337–349. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin × Treatment | B. thuringiensis Survival Assay | ||
|---|---|---|---|
| ‘Coevolved’ CONTROL | ‘Control’ CONTROL | ‘Coevolved’ INFECTION | |
| ‘Control’ CONTROL | 0.05 | ||
| ‘Coevolved’ INFECTION | <0.001 | <0.001 | |
| ‘Control’ INFECTION | <0.001 | <0.001 | <0.001 |
| P. entomophila survival assay | |||
| ‘Control’ CONTROL | 0.19 | ||
| ‘Coevolved’ INFECTION | <0.001 | <0.001 | |
| ‘Control’ INFECTION | <0.001 | <0.001 | 0.80 |
| B. bassiana survival assay | |||
| ‘Control’ CONTROL | 0.004 | ||
| ‘Coevolved’ INFECTION | 0.003 | <0.001 | |
| ‘Control’ INFECTION | 0.022 | <0.001 | 0.397 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, T.; Joop, G.; Rafaluk-Mohr, C. Cross-Resistance: A Consequence of Bi-partite Host-Parasite Coevolution. Insects 2018, 9, 28. https://doi.org/10.3390/insects9010028
Biswas T, Joop G, Rafaluk-Mohr C. Cross-Resistance: A Consequence of Bi-partite Host-Parasite Coevolution. Insects. 2018; 9(1):28. https://doi.org/10.3390/insects9010028
Chicago/Turabian StyleBiswas, Tilottama, Gerrit Joop, and Charlotte Rafaluk-Mohr. 2018. "Cross-Resistance: A Consequence of Bi-partite Host-Parasite Coevolution" Insects 9, no. 1: 28. https://doi.org/10.3390/insects9010028
APA StyleBiswas, T., Joop, G., & Rafaluk-Mohr, C. (2018). Cross-Resistance: A Consequence of Bi-partite Host-Parasite Coevolution. Insects, 9(1), 28. https://doi.org/10.3390/insects9010028