The Influence of Prescribed Fire, Habitat, and Weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Tick Collection
2.3. Weather Data
2.4. Vegetation Sampling
2.5. Statistical Analyses
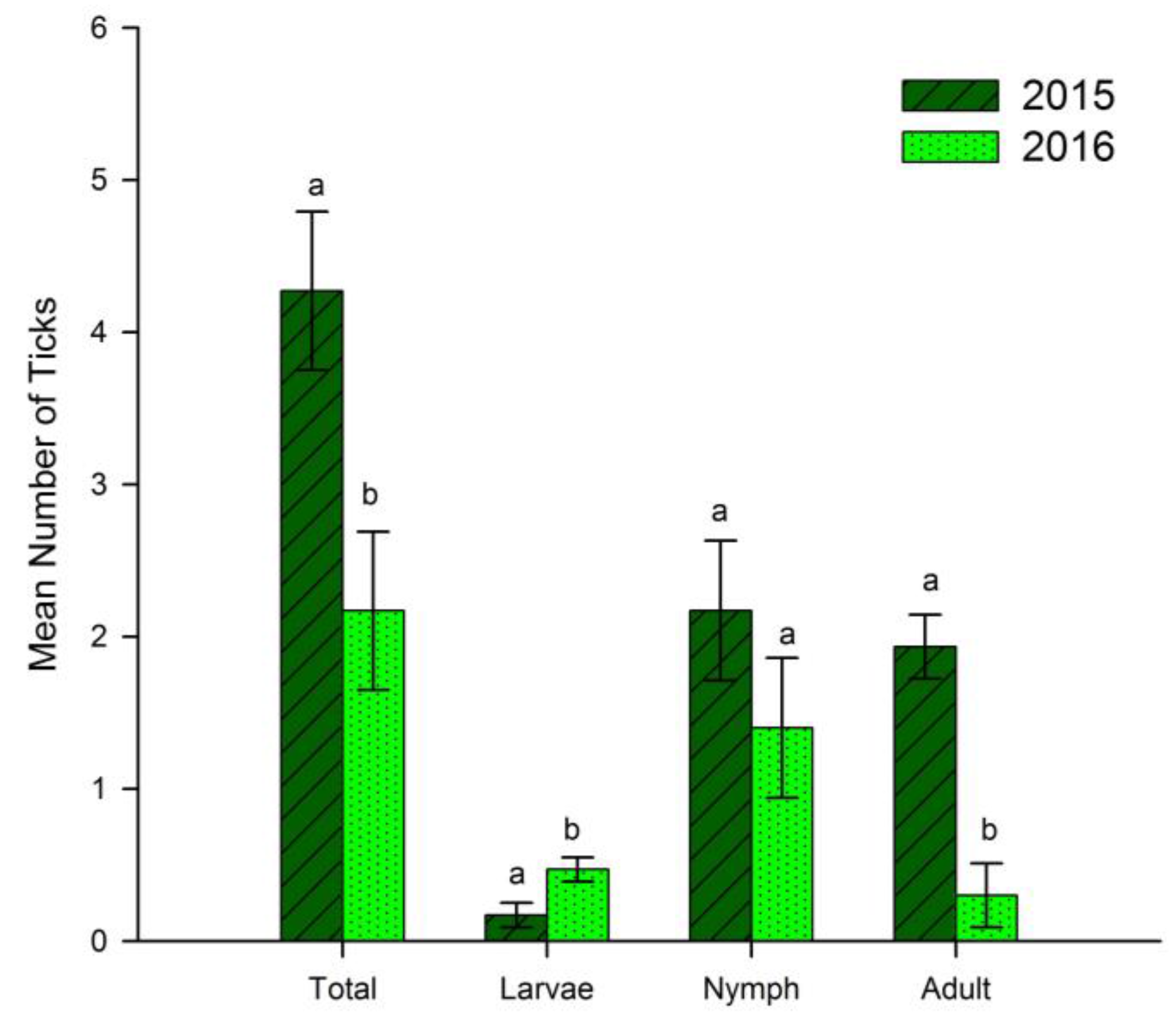
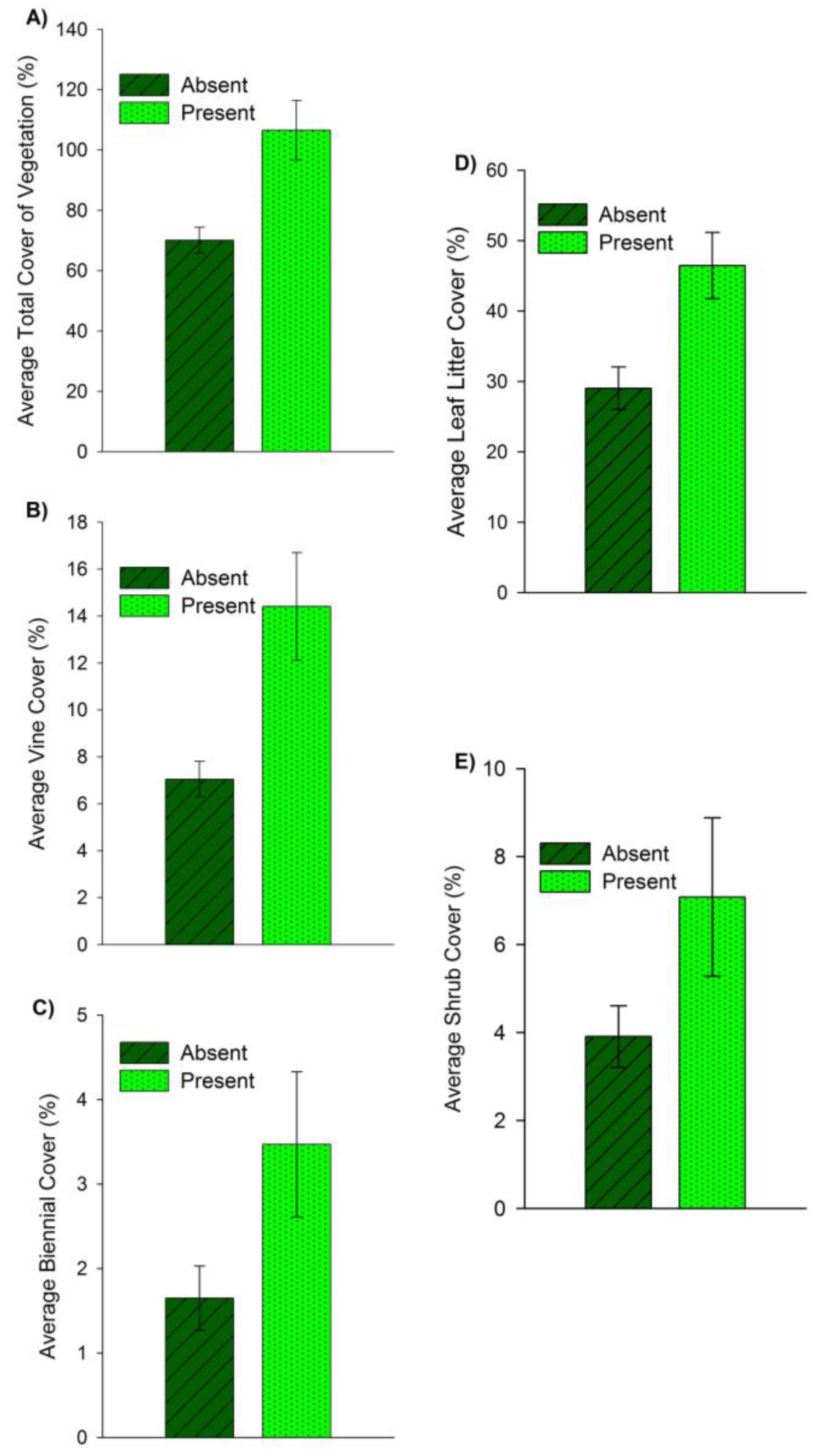
3. Results
3.1. Treatment and Landscape
3.2. Habitat
3.3. Weather
4. Discussion
4.1. Treatment and Landscape
4.2. Habitat
4.3. Weather
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129 (Suppl. S1), S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Stromdahl, E.Y.; Hickling, G.J. Beyond Lyme: Aetiology of tick-borne human diseases with emphasis on the south-eastern United States. Zoonoses Public Health 2012, 59, 48–64. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Tulloch, J.S.P.; Mcginley, L.; Sanchez-Vizcaino, F.; Medlock, J.M.; Radford, A.D. The passive surveillance of ticks using companion animal electronic health records. Epidemiol. Infect. 2017, 145, 2020–2029. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Dobson, A.D.; Levi, T.; Salkeld, D.J.; Swei, A.; Ginsberg, H.S.; Kjemtrup, A.; Padgett, K.A.; Jensen, P.M.; Fish, D. Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philos. Trans. R. Soc. B 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Luckhart, S.; Mullen, G.R.; Wright, J.C. Etiologic agent of Lyme disease, Borrelia burgdorferi, detected in ticks (Acari: Ixodidae) collected at a focus in Alabama. J. Med. Entomol. 1991, 28, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, M.E. The Influence of Prescribed Fire on Tick Abundance and Borrelia burgdorferi Prevalence in Amblyomma americanum. Master’s Thesis, Western Illinois University, Macomb, IL, USA, 2017. [Google Scholar]
- Schulze, T.L.; Bowen, G.S.; Bosler, E.M.; Lakat, M.F.; Parkin, W.E.; Altman, R.; Ormiston, B.G.; Shisler, J.K. Amblyomma americanum: A potential vector of Lyme disease in New Jersey. Science 1984, 224, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Burkhalter, K.L.; Harmon, J.R.; Ashley, D.C.; Panella, N.A.; Lambert, A.; Savage, H.M.; Nicholson, W.L.; Lash, R.R.; Godsey, M.S. First detection of Heartland virus (Bunyaviridae: Phlebovirus) from field collected arthropods. Am. J. Trop. Med. Hyg. 2013, 89, 445–452. [Google Scholar] [CrossRef]
- Childs, J.E.; Paddock, C.D. The ascendancy of Amblyomma americanum as a vector of pathogens affecting humans in the United States. Annu. Rev. Entomol. 2003, 48, 307–337. [Google Scholar] [CrossRef] [PubMed]
- Masters, E.J.; Grigery, C.N.; Masters, R.W. STARI, or Masters Disease: Lone star tick–vectored Lyme-like illness. Infect. Dis. Clin. N. Am. 2008, 22, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.A.; Dahm, N.M.; Ruiz, M.O.; Brown, W.M. Temporal and spatial distribution of tick-borne disease cases among humans and canines in Illinois (2000-2009). Environ. Health Insights 2014, 8, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J.; Varela-Stokes, A.S. Role of the lone star tick, Amblyomma americanum (L.), in human and animal diseases. Vet. Parasitol. 2009, 160, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Monzón, J.D.; Atkinson, E.G.; Henn, B.M.; Benach, J.L. Population and evolutionary genomics of Amblyomma americanum, an expanding arthropod disease vector. Genome Biol. Evol. 2016, 8, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Stegall-Faulk, T.; Clark, D.C.; Wright, S.M. Detection of Borrelia lonestari in Amblyomma americanum (Acari: Ixodidae) from Tennessee. J. Med. Entomol. 2003, 40, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Springer, Y.P.; Eisen, L.; Beati, L.; James, A.M.; Eisen, R.J. Spatial distribution of counties in the continental United States with records of occurrence of Amblyomma americanum (Ixodida: Ixodidae). J. Med. Entomol. 2014, 51, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Trudel, L.; Artsob, H.; Barker, I.K.; Beauchamp, G.; Charron, D.F.; Drebot, M.A.; Galloway, T.D.; O’Handley, R.; Thompson, R.A.; et al. Ixodes scapularis ticks collected by passive surveillance in Canada: Analysis of geographic distribution and infection with Lyme borreliosis agent Borrelia burgdorferi. J. Med. Entomol. 2006, 43, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Stein, K.J.; Waterman, M.; Waldon, J.L. The effects of vegetation density and habitat disturbance on the spatial distribution of ixodid ticks (Acari: Ixodidae). Geospat. Health 2008, 2, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Allan, B.F.; Goessling, L.S.; Storch, G.A.; Thach, R.E. Blood meal analysis to identify reservoir hosts for Amblyomma americanum ticks. Emerg. Infect. Dis. 2010, 16, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Clymer, B.C.; Howell, D.E.; Hair, J.A. Animal hosts of economically important ticks (Acarina) in east-central Oklahoma. Ann. Entomol. Soc. Am. 1970, 63, 612–613. [Google Scholar] [CrossRef]
- Kollars, T.M.; Oliver, J.H.; Durden, L.A.; Kollars, P.G. Host association and seasonal activity of Amblyomma americanum (Acari: Ixodidae) in Missouri. J. Parasitol. 2000, 86, 1156–1159. [Google Scholar] [CrossRef]
- Paddock, C.D.; Yabsley, M.J. Ecological havoc, the rise of white-tailed deer, and the emergence of Amblyomma americanum-associated zoonoses in the United States. Curr. Top. Microbiol. Immunol. 2007, 315, 289–324. [Google Scholar] [PubMed]
- Gill, R.M.A.; Beardall, V. The impact of deer on woodlands: The effects of browsing and seed dispersal on vegetation structure and composition. Forestry 2001, 74, 209–218. [Google Scholar] [CrossRef]
- Abbas, F.; Morellet, N.; Hewison, A.J.M.; Merlet, J.; Cargnelutti, B.; Lourtet, B.; Angibault, J.-M.; Daufresne, T.; Aulagnier, S.; Verheyden, H. Landscape fragmentation generates spatial variation of diet composition and quality in a generalist herbivore. Oecologia 2011, 167, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Nixon, C.M.; Hansen, L.P.; Brewer, P.A.; Chelsvig, J.E. Ecology of white-tailed deer in an intensively farmed region of Illinois. Wildl. Monogr. 1991, 3–77. [Google Scholar]
- Davidson, W.R.; Siefken, D.A.; Creekmore, L.H. Seasonal and annual abundance of Amblyomma americanum (Acari: Ixodidae) in central Georgia. J. Med. Entomol. 1994, 31, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. Bioscience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Procházka, J.; Brom, J.; Št’astný, J.; Pecharová, E. The impact of vegetation cover on temperature and humidity properties in the reclaimed area of a brown coal dump. Int. J. Min. Reclam. Environ. 2011, 25, 350–366. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; de la Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Gleim, E.R.; Conner, L.M.; Berghaus, R.D.; Levin, M.L.; Zemtsova, G.E.; Yabsley, M.J. The phenology of ticks and the effects of long-term prescribed burning on tick population dynamics in Southwestern Georgia and Northwestern Florida. PLoS ONE 2014, 9, e112174. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.S.; Patrick, C.D.; Semtner, P.J.; Hair, J.A. The ecology and behavior of the lone star tick (Acarina: Ixodidae). VI. Response of unfed adults to certain environmental parameters. J. Med. Entomol. 1975, 12, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Dautel, H.; Estrada-Peña, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Effects of microscale habitat physiognomy on the focal distribution of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) nymphs. Environ. Entomol. 2002, 31, 1085–1090. [Google Scholar] [CrossRef]
- Peterson, D.W.; Reich, P.B. Prescribed fire in oak savanna: Fire frequency effects on stand structure and dynamics. Ecol. Appl. 2001, 11, 914–927. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Nix, B.; Jacobs, K.A.; Bowles, M.L. Two decades of low-severity prescribed fire increases soil nutrient availability in a Midwestern, USA oak (Quercus() forest. Geoderma 2012, 183–184, 80–91. [Google Scholar] [CrossRef]
- Barnes, T.A.; Van Lear, D.H. Prescribed fire effects on advanced regeneration in mixed hardwood stands. South. J. Appl. For. 1998, 22, 138–142. [Google Scholar]
- Scasta, J.D. Fire and parasites: An under-recognized form of anthropogenic land use change and mechanism of disease exposure. Ecohealth 2015, 12, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Willis, D.; Carter, R.; Murdock, C.; Blair, B. Relationship between habitat type, fire frequency, and Amblyomma americanum populations in east-central Alabama. J. Vector Ecol. 2012, 37, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, K.A.; Nix, B.; Scharenbroch, B.C. The effects of prescribed burning on soil and litter invertebrate diversity and abundance in an Illinois oak woodland. Nat. Areas J. 2015, 35, 318–327. [Google Scholar] [CrossRef]
- Jenkins, S.E. Spatial Demography of an Ozark Savanna. Ph.D. Thesis, University of Missouri, Columbia, MO, USA, 1997. [Google Scholar]
- Illinois Natural Areas Inventory. Available online: https://www.dnr.illinois.gov/conservation/NaturalHeritage/Documents/Database/INAICountyList.pdf (accessed on 8 December 2017).
- Whelan, R.J. The Ecology of Fire; Cambridge University Press: Cambridge, NY, USA, 1995; p. 116. [Google Scholar]
- Rulison, E.L.; Kuczaj, I.; Pang, G.; Hickling, G.J.; Tsao, J.I.; Ginsberg, H.S. Flagging versus dragging as sampling methods for nymphal Ixodes scapularis (Acari: Ixodidae). J. Vector Ecol. 2013, 38, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Burks, C.S.; Stewart, R.L.; Needham, G.R.; Lee, R.E. The role of direct chilling injury and inoculative freezing in cold tolerance of Amblyomma americanum, Dermacentor variabilis and Ixodes scapularis. Physiol. Entomol. 1996, 21, 44–50. [Google Scholar] [CrossRef]
- Keirans, J.E.; Litwak, T.R. Pictorial key to the adults of hard ticks, family Ixodidae (Ixodida: Ixodoidea), east of the Mississippi River. J. Med. Entomol. 1989, 26, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Keirans, J.E.; Durden, L.A. Illustrated key to nymphs of the tick genus Amblyomma (Acari: Ixodidae) found in the United States. J. Med. Entomol. 1998, 3, 489–495. [Google Scholar] [CrossRef]
- National Centers for Environmental Information [US]. Available online: https://www.ncdc.noaa.gov/ (accessed on 8 December 2017).
- Coulloudon, B.; Eshelman, K.; Gianola, J.; Habich, N.; Hughes, L.; Johnson, C.; Pellant, M.; Podborny, P.; Rasmussen, A.; Robles, B. Sampling Vegetation Attributes; Interagency Technical Ref. 1734–4; BLM Technical Reference: Denver, CO, USA, 1999; pp. 31–130. [Google Scholar]
- Glossary of Forestry Terms from the West Virginia Forestry Association. Available online: http://www.wvfa.org/pdf/sfi/Glossaryofforestryterms.pdf (accessed on 8 December 2017).
- Bolker, B.; Skaug, H.; Magnusson, A.; Nielsen, A. Getting Started with the glmmADMB Package. 2012. Available online: glmmadmb.r-forge.r-project.org/glmmADMB.pdf (accessed on 8 December 2017).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- SigmaPlot, version 10.0; Systat Software Inc.: San Jose, CA, USA, 2007.
- Allan, B.F. Influence of prescribed burns on the abundance of Amblyomma americanum (Acari: Ixodidae) in the Missouri Ozarks. J. Med. Entomol. 2009, 46, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Mather, T.N.; Duffy, D.C.; Campbell, S.R. An unexpected result from burning vegetation to reduce Lyme disease transmission risks. J. Med. Entomol. 1993, 30, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C., III; Ward, J.S.; Magnarelli, L.A. Impact of controlled burns on the abundance of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1998, 35, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.L. Reduced abundance of adult Ixodes dammini (Acari: Ixodidae) following destruction of vegetation. J. Econ. Entomol. 1986, 79, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Padgett, K.A.; Casher, L.E.; Stephens, S.L.; Lane, A.R.S. Effect of prescribed fire for tick control in California chaparral. J. Med. Entomol. 2009, 46, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, S.; Gollino, S.; Zuliani, M.; Nazzi, F. Investigating the relationship between environmental factors and tick abundance in a small, highly heterogeneous region. J. Vector Ecol. 2015, 40, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Mount, G.A.; Haile, D.G. Computer simulation of population dynamics of the American dog tick (Acari: Ixodidae). J. Med. Entomol. 1989, 26, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Trout Fryxell, R.T.; Moore, J.E.; Collins, M.D.; Kwon, Y.; Jean-Philippe, S.R.; Schaeffer, S.M.; Odoi, A.; Kennedy, M.; Houston, A.E. Habitat and vegetation variables are not enough when predicting tick populations in the southeastern United States. PLoS ONE 2015, 10, e0144092. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.C.; Ward, J.S. Effects of Japanese barberry (Ranunculales: Berberidaceae) removal and resulting microclimatic changes on Ixodes scapularis (Acari: Ixodidae) abundances in Connecticut, USA. Environ. Entomol. 2010, 39, 1911–1921. [Google Scholar] [CrossRef] [PubMed]
- Adalsteinsson, S.A.; D’Amico, V.; Shriver, W.G.; Brisson, D.; Buler, J.J. Scale-dependent effects of nonnative plant invasion on host-seeking tick abundance. Ecosphere 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.C.; Linske, M.A.; Ward, J.S. Long-term effects of Berberis thunbergii (Ranunculales: Berberidaceae) management on Ixodes scapularis (Acari: Ixodidae) abundance and Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) prevalence in Connecticut, USA. Environ. Entomol. 2017, 46, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Biases associated with several sampling methods used to estimate abundance of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 1997, 34, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Semtner, P.J.; Barker, R.W.; Hair, J.A. The ecology and behavior of the Lone Star Tick (Acarina: Ixodidae) II. Activity and survival in different ecological habitats. J. Med. Entomol. 1971, 8, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Meekins, J.F.; McCarthy, B.C. Responses of the biennial forest herb Alliaria petiolata to variation in population density, nutrient addition and light availability. J. Ecol. 2000, 88, 447–463. [Google Scholar] [CrossRef]
- Sotala, D.J.; Kirkpatrick, C.M. Foods of white-tailed deer, Odocoileus virginianus, in Martin County, Indiana. Am. Midl. Nat. 1973, 89, 281–286. [Google Scholar] [CrossRef]
- Solberg, V.B.; Neidhardt, K.; Sardelis, M.R.; Hildebrandt, C.; Hoffmann, F.J.; Boobar, L.R. Quantitative evaluation of sampling methods for Ixodes dammini and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 1992, 29, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Patrick, C.D.; Hair, J.A. Oviposition behavior and larval longevity of the lone star tick, Amblyomma americanum (Acarina: Ixodidae), in different habitats. Ann. Entomol. Soc. Am. 1979, 72, 308–312. [Google Scholar] [CrossRef]
- Lancaster, J.L.; McMillan, H.L. The effects of relative humidity on the lone star tick. J. Econ. Entomol. 1955, 48, 338–339. [Google Scholar] [CrossRef]
- Adler, G.H.; Telford, S.R.; Wilson, M.L.; Spielman, A. Vegetation structure influences the burden of immature Ixodes dammini on its main host, Peromyscus leucopus. Parasitology 1992, 105, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Hair, J.A.; Sauer, J.R.; Durham, K.A. Water balance and humidity preference in three species of ticks. J. Med. Entomol. 1975, 12, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Storey, K. Impact of microclimate on immature tick-rodent host interactions (Acari: Ixodidae): Implications for parasite transmission. J. Med. Entomol. 1999, 36, 741–748. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Transect | Treatment | Aspect | Position |
|---|---|---|---|
| Bluff T1 | B15 | South | High |
| Bluff T2 | B15 | South | Low |
| CC1 T1 | B15 | West | High |
| CC1 T2 | B15 | West | High |
| CC2 T1 | B15 | West | High |
| CC2 T2 | B15 | West | High |
| NWBT1 | B04 | Northeast | High |
| NWB T2 | B04 | Northeast | Low |
| NWBPT1 T1 | B04 | Northeast | High |
| NWBPT1 T2 | B04 | Northeast | Low |
| NWBPT2 T1 | B14 | Northeast | High |
| NWBPT2 T2 | B14 | Northeast | Low |
| NWBPT3 T1 | B15 | Northeast | High |
| NWBPT3 T2 | B15 | Northeast | Low |
| PGIW T1 | B14 | Southwest | High |
| PGIW T2 | B14 | Southwest | Low |
| PGPO T1 | B04 | Southwest | High |
| PGPO T2 | B04 | Southwest | Low |
| PGPT T1 | B14 | West | High |
| PGPT T2 | B14 | West | Low |
| PGWO T1 | B04 | Southwest | High |
| PGWO T2 | B04 | Southwest | Low |
| PNDG T1 | B15 | Southwest | High |
| PNDG T2 | B15 | Southwest | Low |
| PNDPT T1 | B04 | Southwest | High |
| PNDPT T2 | B04 | Southwest | Low |
| UPOS T1 | B14 | Southwest | High |
| UPOS T2 | B14 | Southwest | Low |
| WNCK T1 | B14 | Southwest | High |
| WNCK T2 | B14 | Southwest | High |
| Total | Adult | Nymph | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variable | DF | F | p-Value | Spearman ρ | F | p-Value | Spearman ρ | F | p-Value | Spearman ρ |
| Shannon-diversity | 1,56 | 3.9 | 0.05 | −0.30 a | 2.1 | 0.15 | −0.18 | 1.4 | 0.24 | −0.15 |
| Evenness | 1,56 | 2.1 | 0.16 | −0.15 | 0.44 | 0.51 | −0.08 | 2.2 | 0.15 | −0.10 |
| Total Cover | 1,56 | 2.1 | 0.15 | −0.18 | 4.3 | 0.04 | −0.20 | 0.49 | 0.49 | −0.08 |
| Perennial Cover | 1,56 | 5.3 | 0.03 | −0.26 | 3.7 | 0.06 | −0.22 | 3.3 | 0.08 | −0.19 |
| Biennial Cover | 1,56 | 0.64 | 0.43 | −0.31 | 2.6 | 0.11 | −0.38 | <0.01 | 0.98 | −0.12 |
| Annual Cover | 1,56 | 0.97 | 0.33 | −0.20 | 0.46 | 0.50 | −0.27 | 5.0 | 0.03 | 0.0 |
| Grass Cover | 1,56 | 0.27 | 0.61 | 0.12 | 0.17 | 0.68 | 0.03 | 0.99 | 0.32 | 0.21 |
| Sedge Cover | 1,56 | 0.01 | 0.93 | −0.01 | 1.34 | 0.25 | 0.02 | 0.34 | 0.56 | −0.02 |
| Shrub Cover | 1,56 | 0.14 | 0.71 | 0.12 | 0.55 | 0.46 | 0.06 | 1.30 | 0.26 | 0.13 |
| Vine Cover | 1,56 | 0.64 | 0.43 | −0.14 | 2.46 | 0.12 | −0.18 | 0.82 | 0.37 | −0.14 |
| Garlic Mustard Cover | 1,56 | 0.03 | 0.86 | −0.20 | 0.74 | 0.39 | −0.20 | 0.19 | 0.67 | −0.11 |
| Mean Vegetation Height | 1,56 | 0.31 | 0.58 | 0.15 | 0.86 | 0.36 | 0.14 | <0.01 | 0.99 | 0.10 |
| Canopy Cover | 1,56 | 1.36 | 0.25 | −0.19 | 0.89 | 0.35 | −0.11 | 1.29 | 0.26 | −0.16 |
| Leaf Litter Cover | 1,56 | 0.03 | 0.86 | −0.01 | 4.68 | 0.03 | −0.26 | 0.26 | 0.62 | 0.10 |
| Course Woody Debris Cover | 1,56 | 6.89 | 0.01 | −0.40 | 0.70 | 0.41 | −0.49 | 1.04 | 0.31 | −0.14 |
| Bare Ground % | 1,56 | 7.58 | 0.008 | −0.36 | 2.06 | 0.16 | −0.13 | 3.87 | 0.05 | −0.26 |
| Basal Area | 1,56 | 0.03 | 0.86 | 0.0 | 0.24 | 0.62 | 0.13 | <0.01 | 0.95 | −0.04 |
| Parameter | n | β | SE | 95% Confidence Limits | χ2 | p-Value | |
|---|---|---|---|---|---|---|---|
| LCL | UCL | ||||||
| Total Cover | 60 | 0.02 | 0.008 | 0.002 | 0.03 | 4.7 | 0.03 |
| Biennial | 60 | 0.19 | 0.10 | 0.006 | 0.38 | 4.1 | 0.04 |
| Vine | 60 | 0.17 | 0.06 | 0.05 | 0.28 | 8.2 | 0.004 |
| Shrub | 60 | 0.09 | 0.05 | −0.009 | 0.19 | 3.2 | 0.07 |
| Leaf Litter | 60 | 0.04 | 0.01 | 0.013 | 0.07 | 7.9 | 0.005 |
| Response | Parameter | Level (a) | n | β | SE | 95% Confidence Limits | |
|---|---|---|---|---|---|---|---|
| LCL | UCL | ||||||
| Total | Relative Humidity | 75.1 | 420 | 0.0263 | 0.01 | 0.01 | 0.05 |
| Sample Period | 7.5 | 420 | −0.2609 | 0.04 | −0.34 | −0.19 | |
| Precipitation | 1.7 | 420 | 0.3843 | 0.08 | 0.23 | 0.54 | |
| Adult | Relative Humidity | 75.1 | 420 | 0.0337 | 0.01 | 0.01 | 0.06 |
| Sample Period | 7.5 | 420 | −0.4302 | 0.07 | −0.58 | −0.30 | |
| Precipitation | 1.7 | 420 | 0.2764 | 0.10 | 0.08 | 0.47 | |
| Nymphs | Temperature | 19.4 | 420 | 0.0448 | 0.03 | −0.01 | 0.10 |
| Relative Humidity | 75.1 | 420 | 0.0288 | 0.01 | 0.004 | 0.06 | |
| Sample Period | 7.5 | 420 | −0.2083 | 0.05 | −0.31 | −0.12 | |
| Precipitation | 1.7 | 420 | 0.5165 | 0.10 | 0.32 | 0.72 | |
| Larvae | Precipitation | 1.7 | 420 | −0.5690 | 0.34 | −1.4 | −0.004 |
| Response | Parameter | Level (a) | n | β | SE | 95% Confidence Limits | |
|---|---|---|---|---|---|---|---|
| LCL | UCL | ||||||
| Total | Temperature | 22.2 | 450 | 0.0773 | 0.03 | 0.0 | 0.14 |
| Precipitation | 1.4 | 450 | 0.3962 | 0.17 | 0.05 | 0.74 | |
| Adult | Relative Humidity | 58.0 | 450 | −0.1049 | 0.05 | −0.21 | −0.02 |
| Nymphs | Temperature | 22.2 | 450 | 0.1033 | 0.04 | 0.03 | 0.18 |
| Sample Period | 8 | 450 | −0.1894 | 0.05 | −0.29 | −0.10 | |
| Wind | 6.9 | 450 | 0.1211 | 0.06 | 0.00 | 0.25 | |
| Cloud Cover | Clear | 180 | −2.26 | 0.25 | −2.80 | −1.79 | |
| Partly | 120 | 0.063 | 0.37 | −0.68 | 0.80 | ||
| Full | 150 | −1.403 | 0.64 | −2.87 | −0.28 | ||
| Larvae | Sample Period | 8 | 450 | 0.24 | 0.06 | 0.13 | 0.36 |
| Wind | 6.9 | 450 | −0.1653 | 0.06 | −0.29 | −0.05 | |
| Precipitation | 1.4 | 450 | 0.6142 | 0.26 | −0.28 | 0.63 | |
| Cloud Cover | Clear | 180 | −2.732 | 0.31 | −3.41 | −2.17 | |
| Partly | 120 | −0.8653 | 0.59 | −0.22 | 1.48 | ||
| Full | 150 | 0.6241 | 0.43 | −2.17 | 0.23 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilliam, M.E.; Rechkemmer, W.T.; McCravy, K.W.; Jenkins, S.E. The Influence of Prescribed Fire, Habitat, and Weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA. Insects 2018, 9, 36. https://doi.org/10.3390/insects9020036
Gilliam ME, Rechkemmer WT, McCravy KW, Jenkins SE. The Influence of Prescribed Fire, Habitat, and Weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA. Insects. 2018; 9(2):36. https://doi.org/10.3390/insects9020036
Chicago/Turabian StyleGilliam, Mary E., Will T. Rechkemmer, Kenneth W. McCravy, and Seán E. Jenkins. 2018. "The Influence of Prescribed Fire, Habitat, and Weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA" Insects 9, no. 2: 36. https://doi.org/10.3390/insects9020036
APA StyleGilliam, M. E., Rechkemmer, W. T., McCravy, K. W., & Jenkins, S. E. (2018). The Influence of Prescribed Fire, Habitat, and Weather on Amblyomma americanum (Ixodida: Ixodidae) in West-Central Illinois, USA. Insects, 9(2), 36. https://doi.org/10.3390/insects9020036