Quiescence in Aedes aegypti: Interpopulation Differences Contribute to Population Dynamics and Vectorial Capacity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Strains
2.2. Parental Generation (PG)
2.3. Quiescent Eggs
2.4. Effect of Quiescence on Different Biological Parameters of A. aegypti
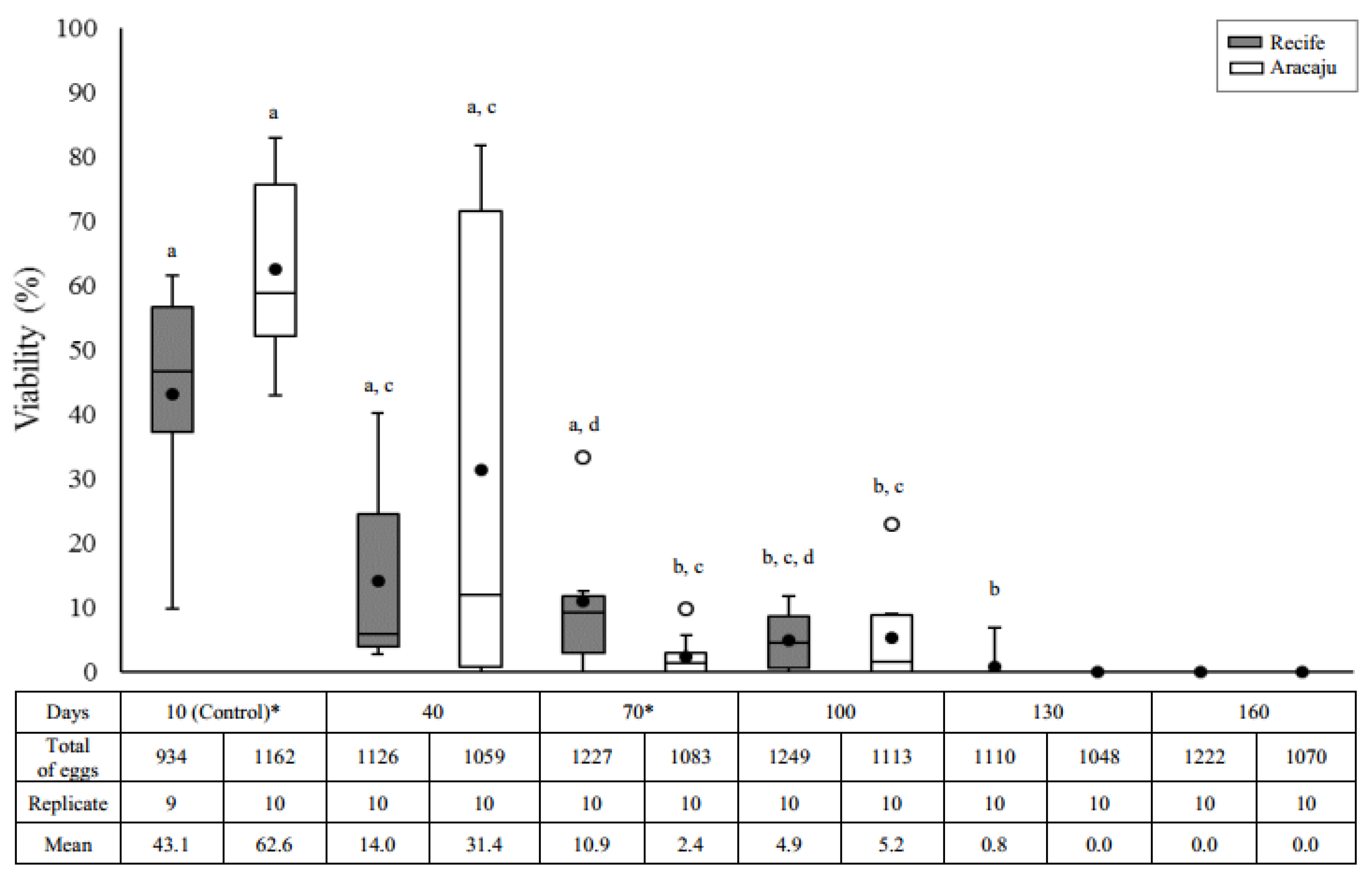
2.4.1. Egg Viability
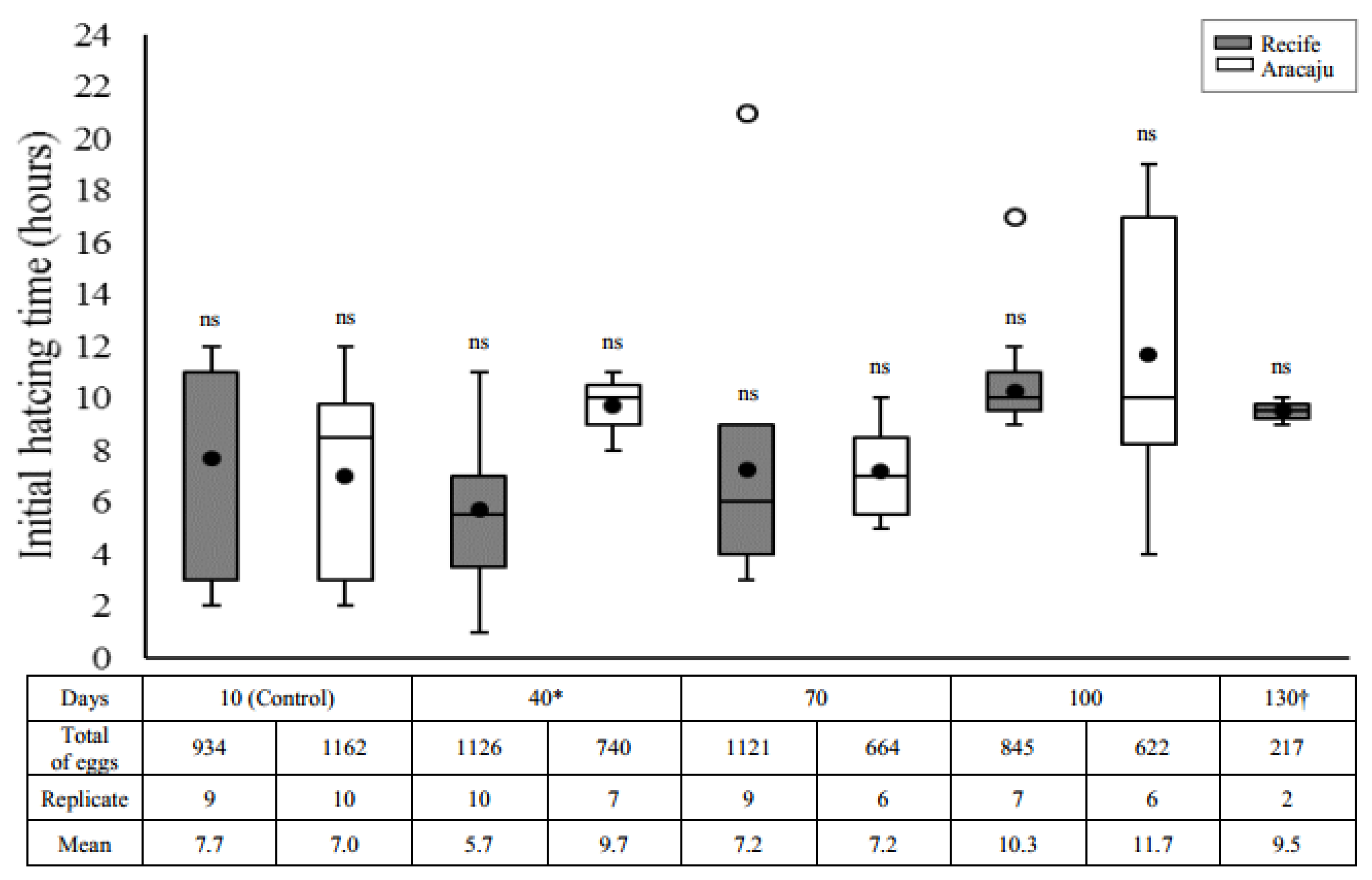
2.4.2. Initial Hatching Time
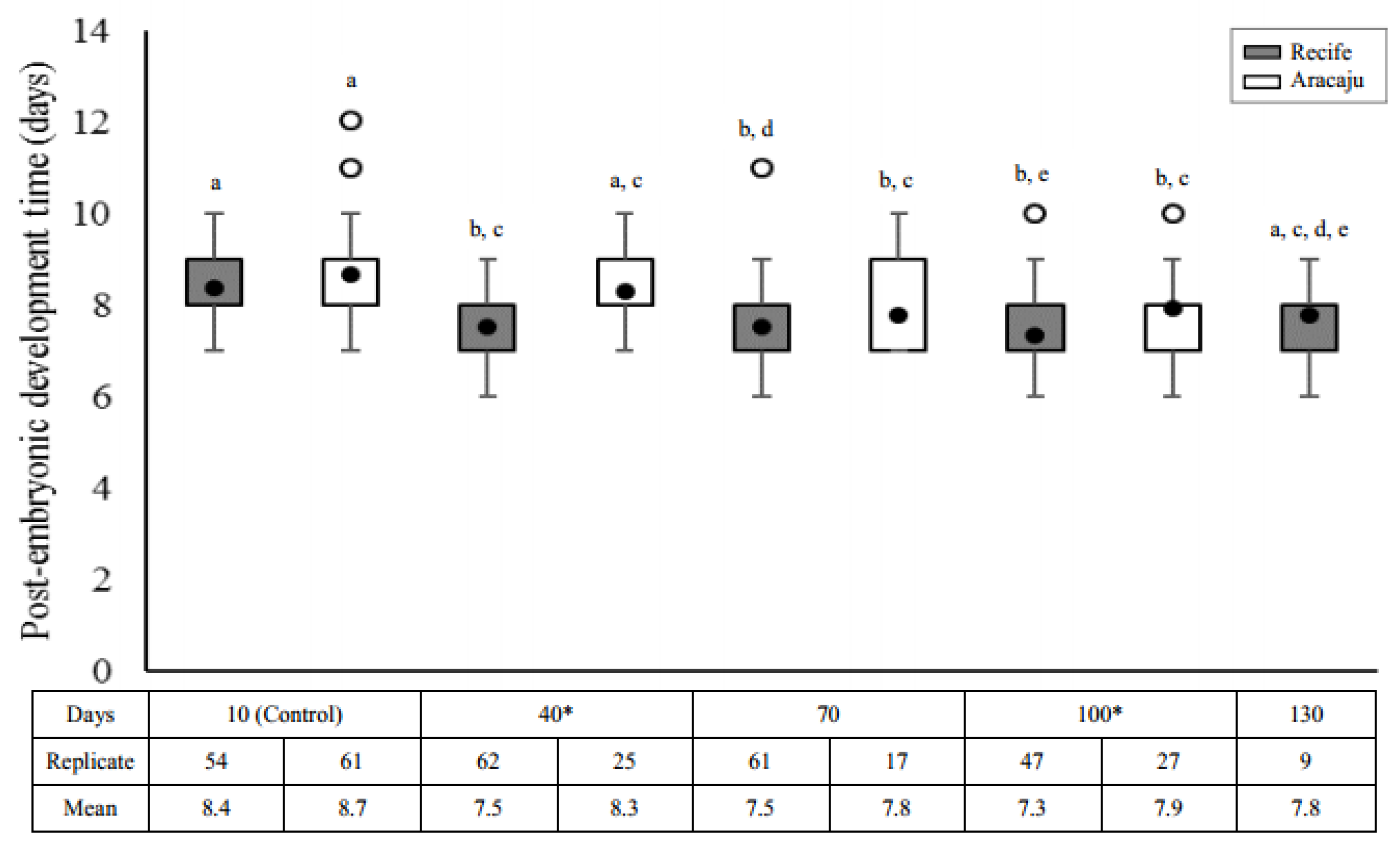
2.4.3. Post-Embryonic Development Time
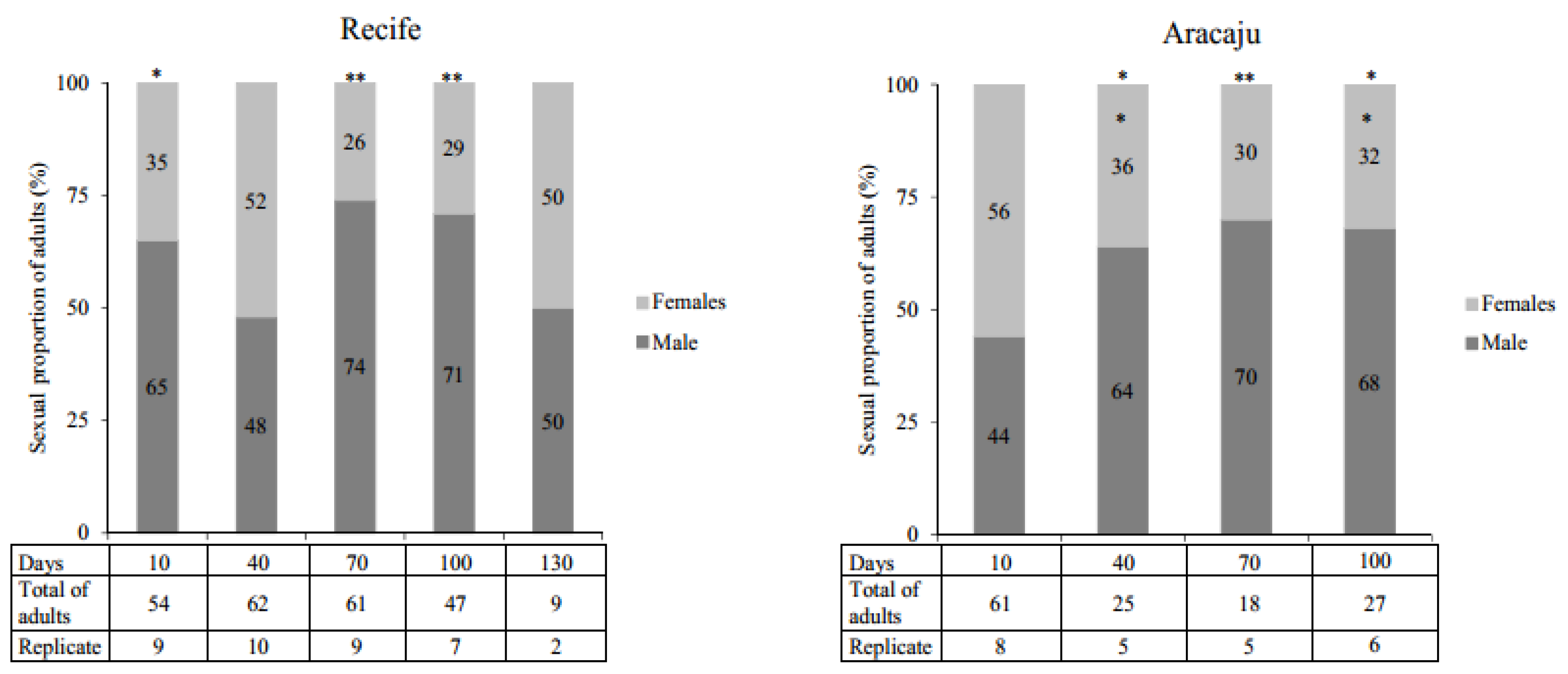
2.4.4. Emergence Rate and Sexual Proportion
2.4.5. Fecundity and Fertility
2.4.6. Adult Size
2.5. Statistical Analyses
3. Results
3.1. Egg Viability
3.2. Initial Hatching Time
3.3. Post-Embryonic Development Time
3.4. Emergence Rate and Sexual Proportion
3.5. Fecundity and Fertility
3.6. Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors.Aedes aegypti and Ae. Albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R.; Powers, A.M. Chikungunya: Epidemiology. F1000Research 2016, 5, F1000 Faculty Rev-82. [Google Scholar] [CrossRef] [PubMed]
- Alaniz, A.J.; Bacigalupo, A.; Cattan, P.E. Spatial quantification of the world population potentially exposed to Zika virus. Int. J. Epidemiol. 2017, 46, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.J.; Wilson, A.J.; Hay, S.I.; Graham, A.J. The global distribution of yellow fever and dengue. Adv. Parasitol. 2006, 62, 181–220. [Google Scholar] [PubMed]
- Messina, J.P.; Brady, O.J.; Scott, T.W.; Zou, C.; Pigott, D.M.; Duda, K.A.; Bhatt, S.; Katzelnick, L.; Howes, R.E.; Battle, K.E.; et al. Global spread of dengue virus types: Mapping the 70 year history. Trends Microbiol. 2014, 22, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Rezza, G. Dengue and chikungunya: Long-distance spread and outbreaks in naïve areas. Pathog. Glob. Health 2014, 108, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Nsoesie, E.O.; Kraemer, M.U.; Golding, N.; Pigott, D.M.; Brady, O.J.; Moyes, C.L.; Johansson, M.A.; Gething, P.W.; Velayudhan, R.; Khan, K.; et al. Global distribution and environmental suitability for chikungunya virus, 1952 to 2015. Euro Surveill. 2016, 21, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Shragai, T.; Tesla, B.; Murdock, C.; Laura, C.H. Zika and chikungunya: Mosquito-borne viruses in a changing world. Ann. N. Y. Acad. Sci. 2017, 1399, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Clark, G.G.; Lorenz, L.H.; Amerasinghe, P.H.; Reiter, P.; Edman, J.D. Detection of multiple blood feeding in Aedes aegypti (Diptera: Culicidae) during a single gonotrophic cycle using a histologic technique. J. Med. Entomol. 1993, 30, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, H.H.; da Silva, I.G. The effect of the period of egg quiescence on the life cycle of Aedes aegypti (Linnaeus, 1762) (Diptera, Culicidae) under laboratory conditions. Rev. Soc. Bras. Med. Trop. 1999, 32, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Colton, Y.M.; Chadee, D.D.; Severson, D.W. Natural skip oviposition of the mosquito Aedes aegypti indicated by codominant genetic markers. Med. Vet. Entomol. 2003, 17, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P. Oviposition, dispersal, and survival in Aedes aegypti: Implications for the efficacy of control strategies. Vector-Borne Zoonotic Dis. 2007, 7, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Oliva, L.O.; Correia, J.C.; Albuquerque, C.M.R. How mosquito age and the type and color of oviposition sites modify skip-oviposition behavior in Aedes aegypti (Diptera: Culicidae)? J. Insect Behav. 2014, 27, 81–91. [Google Scholar] [CrossRef]
- Chitolina, R.F.; Anjos, F.A.; Lima, T.S.; Castro, E.A.; Costa-Ribeiro, M.C. V Raw sewage as breeding site to Aedes (Stegomyia) aegypti (Diptera, Culicidae). Acta Trop. 2016, 164, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Rezende, G.L.; Martins, A.J.; Gentile, C.; Farnesi, L.C.; Pelajo-Machado, M.; Peixoto, A.A.; Valle, D. Embryonic desiccation resistance in Aedes aegypti: Presumptive role of the chitinized serosal cuticle. BMC Dev. Biol. 2008, 8, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Vargas, H.C.M.; Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Serosal cuticle formation and distinct degrees of desiccation resistance in embryos of the mosquito vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus. J. Insect Physiol. 2014, 62, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.F.A.; De Albuquerque, C.M.R.; Oliva, L.O.; De Melo-Santos, M.A.V.; Ayres, C.F.J. Diapause and quiescence: Dormancy mechanisms that contribute to the geographical expansion of mosquitoes and their evolutionary success. Parasit. Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Embryonic development of Aedes aegypti (Diptera: Culicidae): Influence of different constant temperatures. Mem. Inst. Oswaldo Cruz 2009, 104, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Farnesi, L.C.; Menna-Barreto, R.F.S.; Martins, A.J.; Valle, D.; Rezende, G.L. Physical features and chitin content of eggs from the mosquito vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus: Connection with distinct levels of resistance to desiccation. J. Insect Physiol. 2015, 83, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Christophers, S.R. Aedes aegypti (L.) the Yellow FEVER Mosquito, 1st ed.; Cambridge University Press: London, UK, 1960. [Google Scholar]
- Kliewer, J.W. Weight and hatchability of Aedes aegypti eggs (Diptera: Culicidae). Ann. Entomol. Soc. Am. 1961, 54, 912–917. [Google Scholar] [CrossRef]
- Faull, K.J.; Williams, C.R. Intraspecific variation in desiccation survival time of Aedes aegypti (L.) mosquito eggs of Australian origin. J. Vector Ecol. 2015, 40, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, D.; Carbajo, A.E. Aedes aegypti, Aedes albopictus, and dengue in Argentina: Current knowledge and future directions. Mem. Inst. Oswaldo Cruz 2008, 103, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Capinha, C.; Rocha, J.; Sousa, C.A. Macroclimate determines the global range limit of Aedes aegypti. Ecohealth 2014, 11, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.; Choudhury, M.A.; Tong, S. Neglected Tropical Diseases—Oceania—Dengue: Emergence, Determinants and Climate Change; Loukas, A., Ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Chan, M.; Johansson, M.A. The incubation periods of dengue viruses. PLoS ONE 2012, 7, e50972. [Google Scholar] [CrossRef] [PubMed]
- Rohani, A.; Wong, Y.C.; Zamre, I.; Lee, H.L.; Zurainee, M.N. The effect of extrinsic incubation temperature on development of dengue serotype 2 and 4 viruses in Aedes aegypti (L.). Southeast Asian J. Trop. Med. Public Health 2009, 40, 942–950. [Google Scholar] [PubMed]
- Mourya, D.T.; Gokhale, M.D.; Basu, A.; Barde, P.V.; Sapkal, G.N.; Padbidri, V.S.; Gore, M.M. Horizontal and vertical transmission of dengue virus type 2 in highly and lowly susceptible strains of Aedes aegypti mosquitoes. Acta Virol. 2001, 45, 67–71. [Google Scholar] [PubMed]
- Satoto, T.B.T.; Listyantanto, A.; Agustjahjani, S.D.; Josef, H.K.; Widartono, B.S. Vertical transmission of dengue virus in the Yogyakarta airport area. Environ. Health Prev. Med. 2018, 23, 22. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.H.; Noriega, F.G. Aedes aegypti pharate 1(st) instar quiescence affects larval fitness and metal tolerance. J. Insect Physiol. 2012, 58, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.H.; Noriega, F.G. Aedes aegypti pharate 1(st) instar quiescence: A case for anticipatory reproductive plasticity. J. Insect Physiol. 2013, 59, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Ayres, C.E.J.; Melo-Santos, M.A.V.; Prota, J.R.M.; Solé-Cava, A.M.; Regis, L.; Furtado, A.E. Genetic structure of natural populations of Aedes aegypti at the micro- and macrogeographic levels in Brazil. J. Am. Mosq. Control Assoc. 2004, 20, 350–356. [Google Scholar] [PubMed]
- Gonçalves, C.M.; Melo, F.F.; Bezerra, J.M.T.; Chaves, B.A.; Silva, B.M.; Silva, L.D.; Pessanha, J.E.M.; Arias, J.R.; Secundino, N.F.C.; Norris, D.E.; et al. Distinct variation in vector competence among nine field populations of Aedes aegypti from a Brazilian dengue-endemic risk city. Parasit. Vectors 2014, 7, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 4, 1633–1644. [Google Scholar] [CrossRef]
- Prefeitura da Cidade de Recife. Available online: http://www.turismonorecife.com.br/pt-br/informacoes-importantes/clima-e-temperatura (accessed on 27 June 2018).
- IBGE. Instituto Brasileiro de Geografia e Estatística. Available online: https://cidades.ibge.gov.br/brasil/se/aracaju/panorama (accessed on 1 June 2018).
- Prefeitura da Cidade de Aracaju. Available online: http://www.aracaju.se.gov.br/aracaju/aspectos_geograficos (accessed on 27 June 2018).
- Consoli, R.A.G.B.; de Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Fiocruz, Ed.; Fiocruz: Rio de Janeiro, Brazil, 1994; Volume 11, ISBN 9788575412909. [Google Scholar]
- Mendonça, J.A. Quiescência dos ovos na população de Aedes (Stegomyia) aegypti (Linnaeus, 1762) da Universidade Federal de Pernambuco: Importância para o programa Zerando a Dengue; Universidade Federal de Pernambuco: Recife, Brazil, 2016. [Google Scholar]
- Gama, R.A.; de Alves, K.C.; Martins, R.F.; Eiras, Á.E.; de Resende, M.C. Efeito da densidade larval no tamanho de adultos de Aedes aegypti criados em condições de laboratório. Rev. Soc. Bras. Med. Trop. 2005, 38, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E.; Knight, K.L. Taxonomists’ Glossary of Mosquito Anatomy; Plexus Publishing, Inc.: Marlton, NJ, USA, 1980. [Google Scholar]
- Ayres, M.; Ayres, M., Jr.; Ayres, D.L.; Santos, A. BioEstat 5.3. Aplicações Estatísticas nas Áreas das Ciências Biológicas e médicas; Instituto de Desenvolvimento Sustentável Mamirauá: Pará, Brazil, 2014. [Google Scholar]
- Sota, T.; Mogi, M. Survival time and resistance to desiccation of diapause and non-diapause eggs of temperate Aedes (Stegomyia) mosquitoes. Entomol. Exp. Appl. 1992, 63, 155–161. [Google Scholar] [CrossRef]
- Diniz, D.F.A.; de Melo-Santos, M.A.V.; de Santos, E.M.M.; Beserra, E.B.; Helvecio, E.; de Carvalho-Leandro, D.; dos Santos, B.S.; de Menezes Lima, V.L.; Ayres, C.F.J. Fitness cost in field and laboratory Aedes aegypti populations associated with resistance to the insecticide temephos. Parasit. Vectors 2015, 8, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Soares-Pinheiro, V.C.; Dasso-Pinheiro, W.; Trindade-Bezerra, J.M.; Tadei, W.P. Eggs viability of Aedes aegypti Linnaeus (Diptera, Culicidae) under different environmental and storage conditions in Manaus, Amazonas, Brazil. Braz. J. Biol. 2017, 77, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.-L.; Zhang, D.-J.; Damiens, D.D.; Lees, R.S.; Gilles, J.R.L. Standard operating procedures for standardized mass rearing of the dengue and chikungunya vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)-II-Egg storage and hatching. Parasit. Vectors 2015, 8, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Farnesi, L.C.; Vargas, H.C.M.; Valle, D.; Rezende, G.L. Darker eggs of mosquitoes resist more to dry conditions: Melanin enhances serosal cuticle contribution in egg resistance to desiccation in Aedes, Anopheles and Culex vectors. PLoS Negl. Trop. Dis. 2017, 11, e0006063. [Google Scholar] [CrossRef] [PubMed]
- Urbanski, J.M.; Benoit, J.B.; Michaud, M.R.; Denlinger, D.L.; Armbruster, P. The molecular physiology of increased egg desiccation resistance during diapause in the invasive mosquito, Aedes albopictus. Proc. Biol. Sci. 2010, 277, 2683–2692. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, A.J. ABC of oxygen: Oxygen at high altitude. BMJ 1998, 317, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, D. Low oxygen pressure as a driving factor for the altitudinal decline in taxon richness of stream macroinvertebrates. Oecologia 2008, 154, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Judson, C.L.; Gojrati, H.A.N. The effects of various oxygen tensions on embryogeny and larval responses of Aedes aegypti. Entomol. Exp. Appl. 1967, 10, 181–188. [Google Scholar] [CrossRef]
- Gjullin, C.M.; Hegarty, C.P.; Bollen, W.B. The necessity of a low oxygen concentration for the hatching of Aedes mosquito eggs. J. Cell. Comp. Physiol. 1941, 17, 193–202. [Google Scholar] [CrossRef]
- Judson, C.L. The physiology of hatching of Aedine mosquito eggs: Hatching stimulus. Ann. Entomol. Soc. Am. 1960, 53, 688–691. [Google Scholar] [CrossRef]
- Djawdan, M.; Chippindale, A.K.; Rose, M.R.; Bradley, T.J. Metabolic reserves and evolved stress resistance in Drosophila melanogaster. Physiol. Zool. 1998, 71, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Moss, J.I.; Jang, E.B. Effects of age and metabolic stress on heat tolerance of mediterranean fruit fly (Diptera: Tephritidae) eggs. J. Econ. Entomol. 1991, 84, 537–541. [Google Scholar] [CrossRef]
- Craig, G.B., Jr.; Vandehey, R.C.; Hickey, W.A. Genetic variability in populations of Aedes aegypti. Bull. World Health Organ. 1961, 24, 527–539. [Google Scholar] [PubMed]
- Hickey, W.A.; Craig, G.B. Genetic distortion of sex ratio in a mosquito, Aedes aegypti. Genetics 1966, 53, 1177–1196. [Google Scholar] [PubMed]
- Krzywinska, E.; Kokoza, V.; Morris, M.; De La Casa-Esperon, E.; Raikhel, A.S.; Krzywinski, J. The sex locus is tightly linked to factors conferring sex-specific lethal effects in the mosquito Aedes aegypti. Heredity (Edinb) 2016, 117, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Ducheyne, E.; Tran Minh, N.N.; Haddad, N.; Bryssinckx, W.; Buliva, E.; Simard, F.; Malik, M.R.; Charlier, J.; De Waele, V.; Mahmoud, O.; et al. Current and future distribution of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in WHO Eastern Mediterranean Region. Int. J. Health Geogr. 2018, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period of Quiescence (Day) | Population | Mann–Whitney U-Test | p-Value | |
|---|---|---|---|---|
| Recife | Aracaju | |||
| 10 days | 57.5 ± 22.76 (10.0–100) a n = 94 | 52.8 ± 23.13 (0–91.7) a n = 117 | 41.5 | 0.7751 |
| 40 days | 86.8 ± 9.74 (61.5–100) b,c n = 75 | 34.2 ± 30.48 (0–90) a n = 66 | 6.0 | 0.0047 |
| 70 days | 74.5 ± 15.40 (40.0–100) a,c n = 88 | 61.7 ± 35.37 (0–100) a n = 26 | 18.5 | 0.3165 |
| 100 days | 84.5 ± 12.29 (66.7–100) a,c n = 58 | 73.6 ± 30.14 (22.2–100) a n = 42 | 17.0 | 0.5700 |
| 130 days | 100 ± 0.00 (100–100) b,c n = 9 | No emergence | NA | NA |
| 160 days | No emergence | No emergence | NA | NA |
| Period of Quiescence | Population | Mann–Whitney Test | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Recife | Aracaju | |||||||
| Males | Females | Males | Females | Males | Females | Males | Females | |
| 10 days | 2.1 ± 0.13 (1.6–2.7) a,A n = 15 | 2.7 ± 0.09 (2.3–2.9) a,B n = 15 | 2.1 ± 0.06 (2.0–2.3) a,A n = 15 | 2.7 ± 0.09 (2.3–2.9) a,B n = 15 | 99.0 | 94.5 | 0.5755 | 0.4553 |
| 40 days | 2.2 ± 0.11 (2.0–2.5) a,A n = 15 | 2.6 ± 0.09 (2.4–2.9) a,B n = 15 | 2.2 ± 0.10 (1.9–2.5) a,A n = 15 | 2.6 ± 0.17 (2.0–2.9) a,B n = 10 | 100.5 | 69.5 | 0.6187 | 0.7603 |
| 70 days | 2.1 ± 0.10 (1.9–2.6) a,A n = 15 | 2.7 ± 0.05 (2.5–2.9) a,A n = 15 | 2.2 ± 0.33 (1.9–2.7) a,A n = 06 | 2.5 ± 0.12 (2.2–2.8) a,B n = 11 | 37.0 | 34.5 | 0.5334 | 0.0127 |
| 100 days | 2.2 ± 0.15 (1.9–2.8) a,A n = 15 | 2.8 ± 0.04 (2.7–2.9) a,B n = 15 | 2.1 ± 0.11 (1.8–2.7) a,A n = 15 | 2.6 ± 0.08 (2.3–2.8) a,B n = 15 | 51.0 | 53.0 | 0.9158 | 0.0136 |
| 130 days | 2.0 ± 0.36 (1.2–2.4) a,A n = 07 | 2.7 ± 0.52 (2.7–2.8) a,B n = 02 | NA | NA | NA | NA | NA | NA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva, L.O.; La Corte, R.; Santana, M.O.; Albuquerque, C.M.R.d. Quiescence in Aedes aegypti: Interpopulation Differences Contribute to Population Dynamics and Vectorial Capacity. Insects 2018, 9, 111. https://doi.org/10.3390/insects9030111
Oliva LO, La Corte R, Santana MO, Albuquerque CMRd. Quiescence in Aedes aegypti: Interpopulation Differences Contribute to Population Dynamics and Vectorial Capacity. Insects. 2018; 9(3):111. https://doi.org/10.3390/insects9030111
Chicago/Turabian StyleOliva, Luciana O., Roseli La Corte, Marcelo O. Santana, and Cleide M. R. de Albuquerque. 2018. "Quiescence in Aedes aegypti: Interpopulation Differences Contribute to Population Dynamics and Vectorial Capacity" Insects 9, no. 3: 111. https://doi.org/10.3390/insects9030111
APA StyleOliva, L. O., La Corte, R., Santana, M. O., & Albuquerque, C. M. R. d. (2018). Quiescence in Aedes aegypti: Interpopulation Differences Contribute to Population Dynamics and Vectorial Capacity. Insects, 9(3), 111. https://doi.org/10.3390/insects9030111