Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Study Area
2.3. Sampling Regime
2.4. Group Size and Group Interaction
2.5. Plant Size and Plant Abundance
2.6. Nearest-Neighbor Analysis for Plant Community
2.7. Geotaxis Experiments
2.8. Statistical Analysis
3. Results
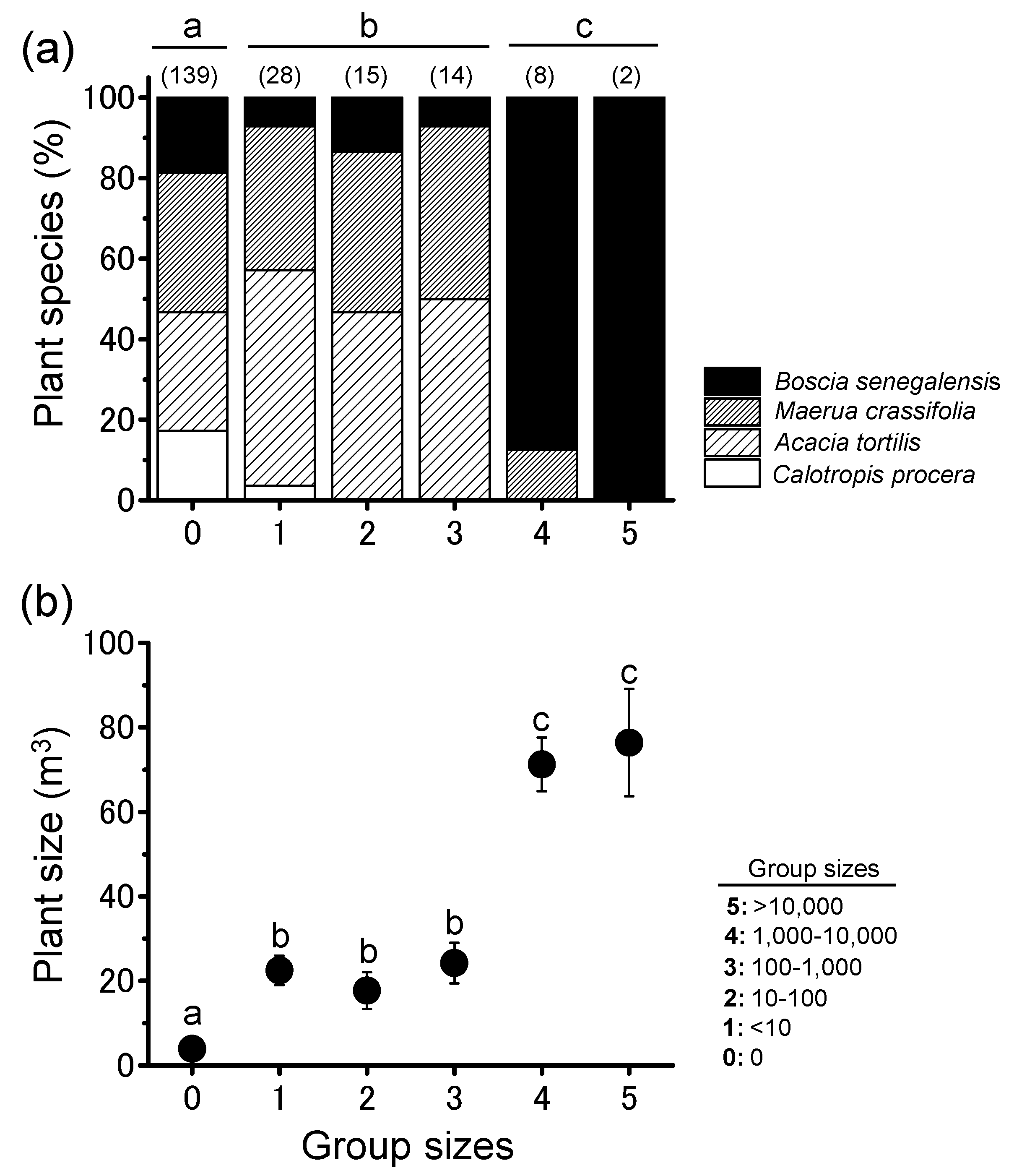
3.1. Night-Roosting Plant Preference
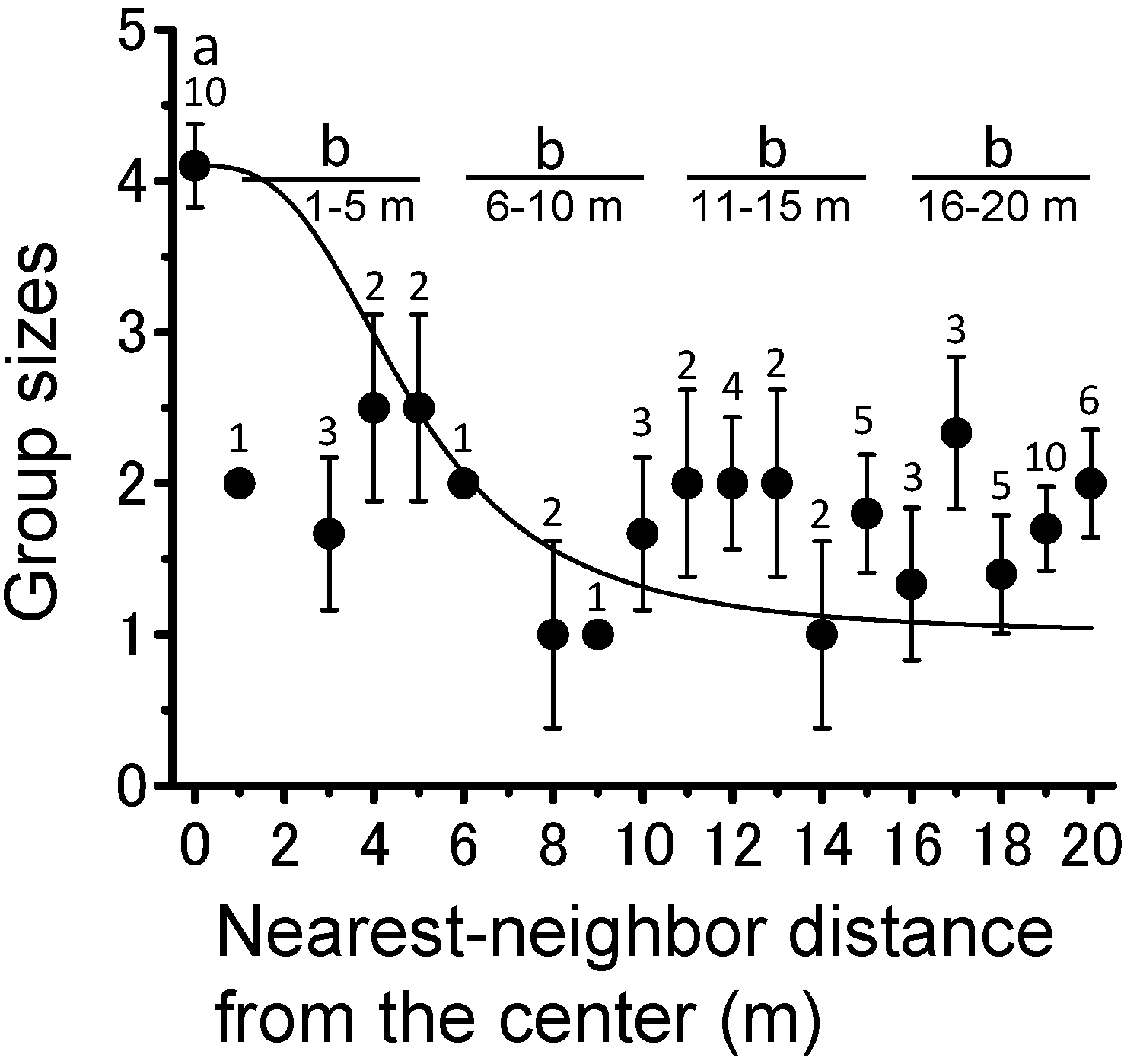
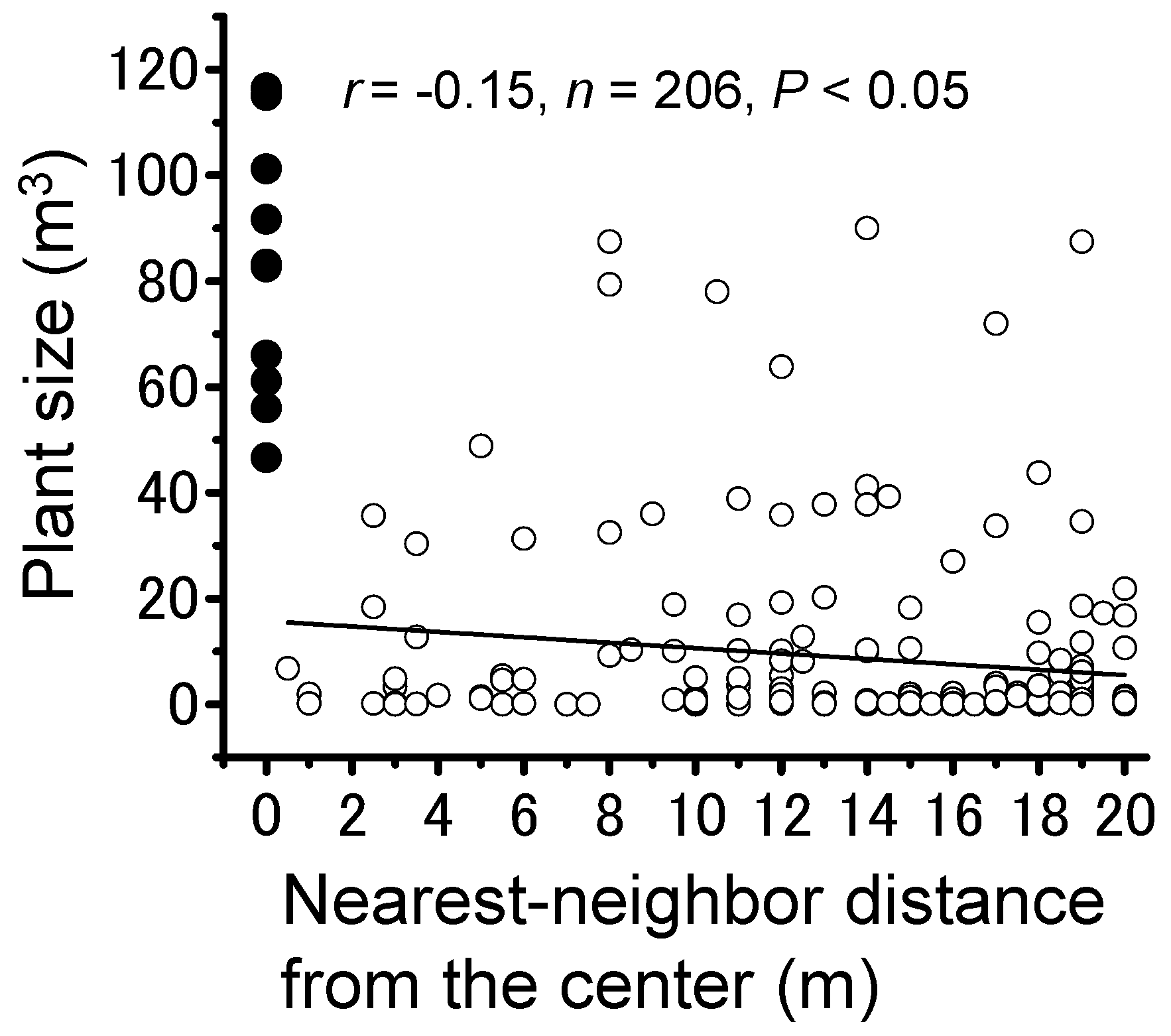
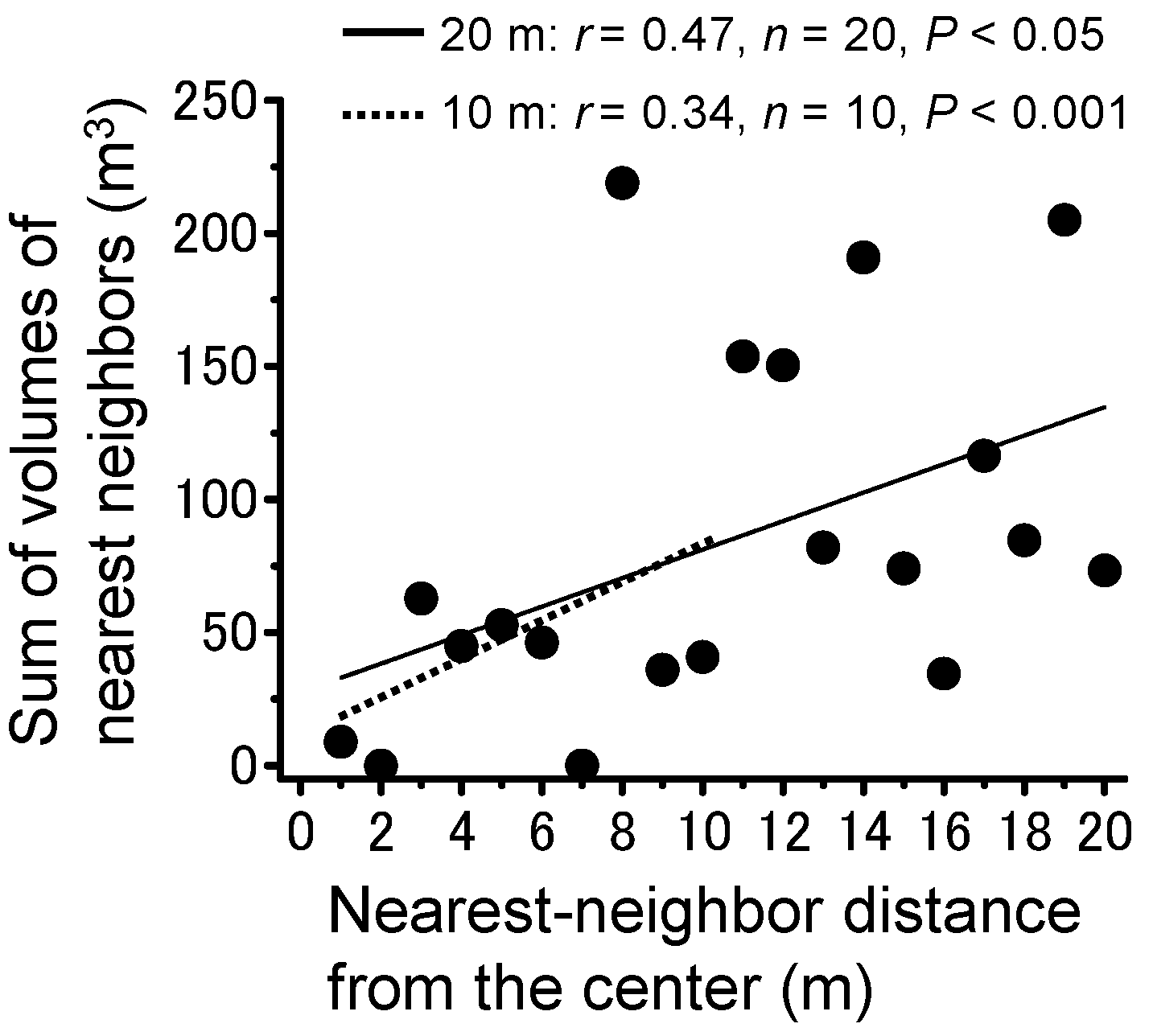
3.2. Locust Groups’ Interaction with the Plant Community
3.3. Plant Community
3.4. Geotaxis Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krause, J.; Ruxton, G.D. Living in Groups; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Beauchamp, G. Social Predation: How Group Living Benefits Predators and Prey; Academic Press: Boston, MA, USA, 2014; ISBN 978-0-12-407228-2. [Google Scholar]
- Hamilton, W.D. Geometry for the selfish herd. J. Theor. Biol. 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Parrish, J.K. Complexity, pattern, and evolutionary trade-offs in animal aggregation. Science 1999, 284, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Devigne, C.; Broly, P.; Deneubourg, J.L. Individual preferences and social interactions determine the aggregation of woodlice. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Jeanson, R.; Deneubourg, J. Conspecific attraction and shelter selection in gregarious insects. Am. Nat. 2007, 170, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R. Ethology and ecology of sleep in monkeys and apes. Adv. Study Behav. 1984, 14, 165–229. [Google Scholar] [CrossRef]
- Hamilton, W.J. Baboon sleeping site preferences and relationships to primate grouping patterns. Am. J. Primatol. 1982, 3, 41–53. [Google Scholar] [CrossRef]
- Uvarov, B.P. Grasshoppers and Locusts; Cambridge University Press: Cambridge, UK, 1966; Volume 1. [Google Scholar]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism:an update. Adv. In Insect Phys. 2009, 36, 1–272. [Google Scholar] [CrossRef]
- Cullen, D.A.; Cease, A.J.; Latchininsky, A.V.; Ayali, A.; Berry, K.; Buhl, J.; De Keyser, R.; Foquet, B.; Hadrich, J.C.; Matheson, T.; et al. From molecules to management: Mechanisms and consequences of locust phase polyphenism. In Advances in Insect Physiology; Academic Press: Cambridge, UK, 2017; Volume 53, pp. 167–285. [Google Scholar]
- Ellis, P.E.; Ashall, C. Field studies on diurnal behaviour, movement and aggregation in the desert locust (Schistocerca gregaria Forskål). Anti-Locust Bull. 1957, 25, 1–94. [Google Scholar]
- Ariel, G.; Ayali, A. Locust collective motion and its modeling. PLoS Comput. Biol. 2015, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.S. The behaviour of the Desert Locust (Schistocerca gregaria (Forsl.))(Orthopt.) in an outbreak centre. Trans. R. Entomol. Soc. Londonent. Soc. Lond. 1939, 89, 385–542. [Google Scholar] [CrossRef]
- Roffey, J.; Popov, G. Environmental and behavioural processes in a Desert Locust outbreak. Nature 1968, 219, 446–450. [Google Scholar] [CrossRef]
- Collett, M.; Despland, E.; Simpson, S.J.; Krakauer, D.C. Spatial scales of desert locust gregarization. Proc. Natl. Acad. Sci. USA 1998, 95, 13052–13055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Despland, E.; Collett, M.; Simpson, S.J. Small-scale processes in desert locust swarm formation: How vegetation patterns influence gregarization. Oikos 2000, 88, 652–662. [Google Scholar] [CrossRef]
- Maeno, O.K.; Piou, C.; Ould Ely, S.; Ould Mohamed, S.; El Hacen Jaavar, M.; Ould Babah, M.A.; Nakamura, S. Field observations of the sheltering behavior of the solitarious phase of the Desert locust, Schistocerca gregaria, with particular reference to antipredator strategies. Jpn. Agric. Res. Q. 2012, 46, 339–345. [Google Scholar] [CrossRef]
- Maeno, O.K.; Ould Ely, S.; Nakamura, S.; Abdellaoui, K.; Cissé, S.; Jaavar, M.E.H.; Ould Mohamed, S.; Atheimine, M.; Ould Babah, M.A. Daily microhabitat shifting of solitarious-phase Desert locust adults: Implications for meaningful population monitoring. Springerplus 2016, 5, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeno, O.K.; Ould Ely, S.; Ould Mohamed, S.; Jaavar, M.E.H.; Nakamura, S.; Ould Babah Ebbe, M.A. Behavioral plasticity in anti-predator defense in the desert locust. J. Arid Environ. 2018, in press. [Google Scholar] [CrossRef]
- Rogers, S.M.; Cullen, D.A.; Anstey, M.L.; Burrows, M.; Despland, E.; Dodgson, T.; Matheson, T.; Ott, S.R.; Stettin, K.; Sword, G.A.; et al. Rapid behavioural gregarization in the desert locust, Schistocerca gregaria entails synchronous changes in both activity and attraction to conspecifics. J. Insect Physiol. 2014, 65, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Ould Babah, M.A. Strategy for controlling the desert locust in Mauritania. In New Strategies in Locust Control; Krall, S., Palf, P., Diallo, B.D., Eds.; Birkhaeuser Verlag: Basel, Switzerland, 1997; pp. 487–491. [Google Scholar]
- Ould Babah, M.A. Biogéographie du Criquet pèlerin en Mauritanie; Academic Press: Paris, France, 2010; pp. 1–268. [Google Scholar]
- Maeno, O.K.; Piou, C.; Ould Ely, S.; Ould Babah, M.A.; Pélissié, B.; Ould Mohamed, S.; Jaavar, M.E.H.; Etheimine, M.; Nakamura, S. Plant size-dependent escaping behavior of gregarious nymphs of the Desert locust, Schistocerca gregaria. J. Insect Behav. 2013, 26, 623–633. [Google Scholar] [CrossRef]
- Fonteyn, P.J.; Mahall, B.E. An experimental analysis of structure in a desert plant community. J. Ecol. 1981, 69, 883–896. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; ISBN 3-900051-07-0. [Google Scholar]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines 1: Biology and Behavior; FAO: Rome, Italy, 2001. [Google Scholar]
- Alcock, J.; Dodson, G. The diverse mating systems of hilltopping insects. Am. Entomol. 2008, 54, 80–87. [Google Scholar] [CrossRef]
- Hassanali, A.; Njagi, P.G.N.; Bashir, M.O. Chemical Ecology of Locusts and Related Acridids. Annu. Rev. Entomol. 2005, 50, 223–245. [Google Scholar] [CrossRef] [PubMed]
- Uvarov, B.P. Grasshoppers and Locusts; Centre for Overseas Pest Research: London, UK, 1977; Volume 2. [Google Scholar]
- Whitman, D.W. Thermoregulation and daily activity patterns in a black desert grasshopper, Taeniopoda eques. Anim. Behav. 1987, 35, 1814–1826. [Google Scholar] [CrossRef]
- Whitman, D.W.; Orsak, L. Biology of Taeniopoda eques (Orthoptera: Acrididae) in southeastern Arizona. Ann. Entomol. 1985, 78, 811–825. [Google Scholar] [CrossRef]
- Scott, D.M.; Waite, S.; Maddox, T.M.; Freer, R.A.; Dunstone, N. The validity and precision of spotlighting for surveying desert mammal communities. J. Arid Environ. 2005, 61, 589–601. [Google Scholar] [CrossRef]
- Whitman, D.W. Function and evolution of thermoregulation in the desert grasshopper Taeniopoda eques. J. Anim. Ecol. 1988, 57, 369–383. [Google Scholar] [CrossRef]
- Peckarsky, B.L. Predator-prey interactions between stoneflies and mayflies: Behavioral observations. Ecology 1980, 61, 932–943. [Google Scholar] [CrossRef]
- Putman, B.J.; Clark, R.W. Behavioral thermal tolerances of free-ranging rattlesnakes (Crotalus oreganus) during the summer foraging season. J. Therm. Biol. 2017, 65, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Alcock, J. Sleeping aggregations of the bee Idiomelissodes duplocincta (Cockerell) (Hymenoptera: Anthophorini) and their possible function. J. Kans. Entomol. Soc. 1998, 71, 74–84. [Google Scholar]
- Sword, G.A.; Lorch, P.D.; Gwynne, D.T. Insect behaviour: Migratory bands give crickets protection. Nature 2005, 433, 703. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, S. Efficiency of area-concentrated searching behaviour in a continuous patchy environment. J. Theor. Biol. 1992, 159, 67–81. [Google Scholar] [CrossRef]
- Van Huis, A.; Cressman, K.; Magor, J.I. Preventing desert locust plagues: Optimizing management interventions. Entomol. Exp. Appl. 2017, 122, 191–214. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | No. of Plants (1256 m2) | No. of Quadrads | Plant | No. of Plants | ||
|---|---|---|---|---|---|---|
| Maximum Width (m) | Maximum Height (m) | Volume (m3) | ||||
| Calotropis procera | 3.4 ± 3.0 | 10 | 0.97 ± 0.25 a | 1.03 ± 0.23 ab | 2.7 ± 4.3 a | 25 |
| Acacia tortilis | 7.1 ± 3.0 | 10 | 1.17 ± 0.15 a | 0.91 ± 0.14 a | 9.1 ± 2.0 a | 71 |
| Maerua crassifolia | 6.1 ± 3.0 | 10 | 2.14 ± 0.15 b | 2.34 ± 0.14 c | 21.8 ± 3.1 b | 70 |
| Boscia senegalensis | 4.1 ± 3.0 | 10 | 2.04 ± 0.19 b | 1.67 ± 0.18 b | 30.4 ± 4.8 b | 40 |
| Source of Variance | Group Size on Roosting Plants | |||
|---|---|---|---|---|
| df | ms | f | p | |
| plant species | 3 | 0.94 | 1.12 | 0.291 |
| plant size | 1 | 16.67 | 19.83 | <0.001 |
| distance from the center | 1 | 0.63 | 0.75 | 0.389 |
| plant species × plant size | 1 | 0.29 | 0.34 | 0.559 |
| plant species × distance | 1 | 0.89 | 1.05 | 0.306 |
| plant size × distance | 1 | 0.22 | 0.27 | 0.607 |
| plant species × plant size × distance | 1 | 3.21 | 3.82 | 0.052 |
| error | 198 | 166.45 | ||
| Source of Variance | Group Size on Roosting Plants | |||
|---|---|---|---|---|
| df | ms | f | p | |
| width | 1 | 0.1327117 | 0.1695 | 0.681 |
| height | 1 | 8.961103 | 11.4446 | 0.0009 |
| width × height | 1 | 0.3010408 | 0.3845 | 0.5359 |
| error | 198 | 166.45 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeno, K.O.; Ould Babah Ebbe, M.A. Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects 2018, 9, 99. https://doi.org/10.3390/insects9030099
Maeno KO, Ould Babah Ebbe MA. Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects. 2018; 9(3):99. https://doi.org/10.3390/insects9030099
Chicago/Turabian StyleMaeno, Koutaro Ould, and Mohamed Abdallahi Ould Babah Ebbe. 2018. "Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania" Insects 9, no. 3: 99. https://doi.org/10.3390/insects9030099
APA StyleMaeno, K. O., & Ould Babah Ebbe, M. A. (2018). Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects, 9(3), 99. https://doi.org/10.3390/insects9030099