Two Decades of Invasive Western Corn Rootworm Population Monitoring in Croatia






{kind=link}
Abstract
:1. Introduction
Invasive Western corn rootworm (WCR)
2. Monitoring Trap Methods
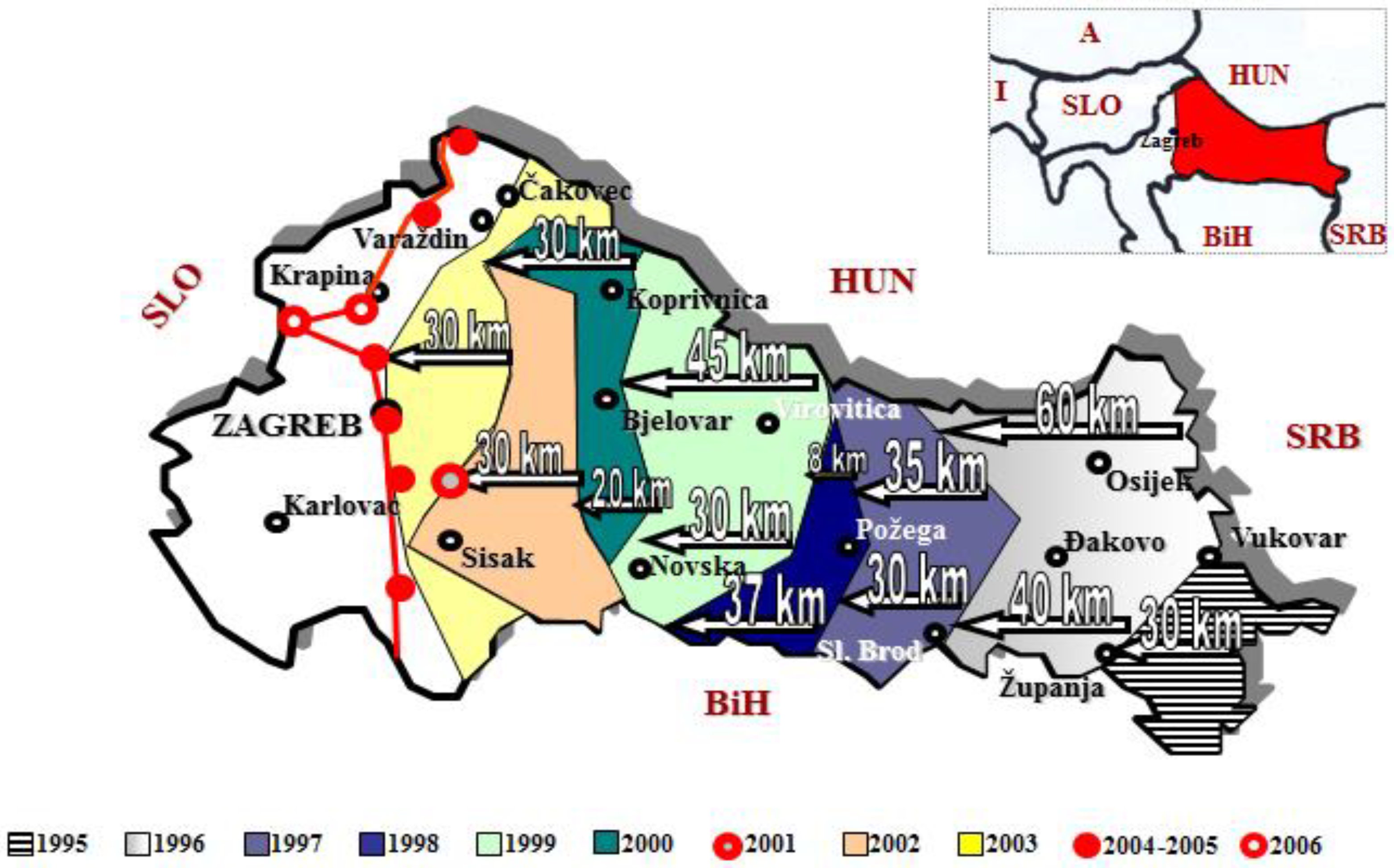
3. Spatial and Density Monitoring
4. Genetic Monitoring
5. Geometric Morphometric Monitoring
6. Future Work
List of Projects Related to WCR in Croatia
- 2017–2021: Monitoring of insect pest resistance: novel approach for detection, and effective resistance management strategies (MONPERES), Croatian science foundation (coordinator: R. Bažok)
- 2009: The landscape genetics of the invasive western corn rootworm in Croatia (Ministry of science, education and sport—Unity through knowledge fund—UKF)
- 2007–2013: The spatial distribution of economically important pests with the use of GIS (Ministry of science, education and sport, Croatia)
- 2005–2007: Developing IPM in maize through WCR risk management—FAO
- 2005–2006: Development of IPM for WCR in collaboration with Secondary agricultural schools-FAO
- 2003–2007: Integrated pest management for western corn rootworm in Central and Eastern Europe (FAO, GTF)
- 2002–2006: Biological control the base of ecologically acceptable plant protection (Ministry of science and technology, Croatia)
- 2002–2004: The possibility of the control of the Western corn rootworm with minimal input (Ministry of agriculture and forestry Croatia)
- 1998–2006: Monitoring of the western corn rootworm (Ministry of agriculture and forestry Croatia)
- 1998–2001: Diabrotica virgifera virgifera (Ministry of science and technology Croatia—young researcher project)
- 1997–2000: Management of Western corn rootworm in central Europe FAO/TCP
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Igrc-Barčić, J.; Bažok, R. The influence of the different food sources on the life parameters of western corn rootworm (Diabrotica virgifera virgifera LeConte, Coleoptera: Chrysomelidae). Razpr. IV Razreda SAZU 2004, 45, 75–86. [Google Scholar]
- Lemic, D.; Mikac, K.M.; Kozina, A.; Benitez, H.A.; McLean, C.M.; Bažok, R. Monitoring techniques of the western corn rootworm are the precursor to effective IPM strategies. Pest Manag. Sci. 2016, 72, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Dobrinčić, R.; Igrc-Barčić, J.; Edwards, R.C. Determining of the injuriousness of the larvae of western corn rootworm (Diabrotica virgifera virgifera LeConte) in Croatian conditions. Agric. Conspec. Sci. 2002, 67, 1–9. [Google Scholar]
- Baca, F. New member of the harmful entomofauna of Yugoslavia, Diabrotica virgifera virgifera LeConte (Coleoptera, Chrysomelidae). Zast. Bilja 1994, 45, 125–131. [Google Scholar]
- Bača, F.; Berger, H.K. Bedroht ein neuer Schadling unsere Maisernten? Pflanzenschutz 1994, 1, 9–10. [Google Scholar]
- Szalai, M.; Komáromi, J.P.; Bažok, R.; Igrc-Barčić, J.; Kiss, J.; Toepfer, S. The growth rate of Diabrotica virgifera virgifera populations in Europe. J. Pest Sci. 2010, 84, 133–142. [Google Scholar] [CrossRef]
- Ciosi, M.; Miller, N.J.; Kim, K.S.; Giordano, R.; Estoup, A.; Guillemaud, T. Invasion of Europe by the western corn rootworm, Diabrotica virgifera virgifera: Multiple transatlantic introductions with various reductions of genetic diversity. Mol. Ecol. 2008, 17, 3614–3627. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.; Edwards, C.R.; Berger, H.K.; Cate, P.; Cean, M.; Cheek, S. Monitoring of western corn rootworm (Diabrotica virgifera virgifera LeConte) in Europe 1992–2003. In Western Corn Rootworm: Ecology and Management, Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005; Vidal, S., Kuhlmann, U., Edwards, C.R., Eds.; CAB International: Wallingford, UK, 2005. [Google Scholar]
- Ciosi, M.; Miller, N.J.; Toepfer, S.; Estoup, A.; Guillemaud, T. StratiŢed dispersal and increasing geneticvariation during the invasion of Central Europe by the western corn rootworm, Diabrotica virgifera virgifera. Evol. Appl. 2010, 4, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Estoup, A.; Toepfer, S.; Bourguet, D.; Lapchin, L.; Derridj, S. Multiple transatlantic introductionsof the western corn rootworm. Science 2005, 310, 992. [Google Scholar] [CrossRef] [PubMed]
- European and Mediterranean Plant Protection Organisation (EPPO). Present Situation of Diabrotica virgifera virgifera in Europe. 2012. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/shortnotes_qps/diabrotica_virgifera (accessed on 4 September 2018).
- Igrc-Barčić, J.; Maceljski, M. Monitoring of Diabrotica virgifera virgifera LeConte in Croatia in 1995. IWGO Newsl. 1996, 16, 11–13. [Google Scholar]
- Dobrinčić, R.; Igrc-Barčić, J. Istraživanja privlačnosti različitih mamaca za kukuruznu zlaticu (Diabrotica virgifera virgifera LeConte, Col. Chrysomelidae). In Sažeci Znanst. Skupa III. Kolokvij—Entomofauna Hrvatske i Susj. Zemalja; Hrvatsko Društvo Biljne Zaštite: Zagreb, Croatia, 1997. [Google Scholar]
- Dobrinčić, R.; Igrc-Barčić, J. Rezultati monitoringa kukuruzne zlatice (Diabrotica virgifera virgifera LeConte) u 1998. godini u Hrvatskoj. In Glasnik Zaštite Bilja; Sažeci 43; Seminara Biljne Zaštite: Opatija, Croatia; Hrvatsko Društvo Biljne Zaštite: Zagreb, Croatia, 1999. [Google Scholar]
- Dobrinčić, R. An Investigation of the Biology and Ecology of Diabrotica virgifera virgifera LeConte, a New Member of the Entomofauna of Croatia. Ph.D. Thesis, Faculty of Agriculture, University of Zagreb, Zagreb, Croatia, 2001. [Google Scholar]
- Dobrinčić, R.; Igrc-Barčić, J.; Edwards, R.C. The Investigation of the Relationship between WCR Population Level and Corn Yield-Croatian Experiencies. In FAO WCR Network Papers, Proceedings of the XXI IWGO Conference and VIII Diabrotica Subgroup Meeting, Venezia, Italy, 29–30 October 2001; Veneto Agricoltura: Legnaro, Italy, 2001. [Google Scholar]
- Igrc-Barčić, J.; Dobrinčić, R. 1998 Results of Monitoring Diabrotica virgifera virgifera LeConte. Acta Phytopathol. Èntomol. Hung. 2002, 37, 137–144. [Google Scholar] [CrossRef]
- Dobrinčić, R.; Igrc-Barčić, J.; Tuska, T.; Galo, A.; Paučova, O.; Karić, N.; Ivanova, I.; Allara, M. Participatory Approach as a Management Tool for Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). In Ecology and Management of Western Corn Rootworm; CABI Publishing: Wallingford, UK; Gottingen, Germany, 2003. [Google Scholar]
- Bažok, R.; Igrc-Barčić, J.; Edwards, C.R. Effects of proteinase inhibitors on western corn rootworm life parameters. J. Appl. Èntomol. 2005, 129, 185–190. [Google Scholar] [CrossRef]
- Bažok, R. Western corn rootworm. Glas. Biljn. Zaštite 2007, 7, 316. [Google Scholar]
- Lemic, D.; Bažok, R. Procjena rizika od kukuruzne zlatice Diabrotica virgifera virgifera LeConte na području Moslavine. Agronomski Glasnik Glasilo Hrvatskog Agronomskog Društva 2010, 71, 337–346. [Google Scholar]
- Bažok, R.; Sivčev, I.; Kos, T.; Igrc-Barčić, J.; Kiss, J.; Jankovič, S. Pherocon AM trapping and the “Whole plant count” method—A comparison of two sampling techniques to estimate the WCR adult densities in Central Europe. Cereal Res. Commun. 2011, 39, 298–305. [Google Scholar] [CrossRef]
- Prinzinger, G. Monitoring of western corn rootworm (Diabrotica virgifera virgifera LeConte) in Hungary in 1995. IWGO News Lett. 1996, 16, 7–11. [Google Scholar]
- Sivcev, I.; Stankovic, S.; Kostic, M.; Lakic, N.; Popovic, Z. Population density of Diabrotica virgifera virgifera LeConte beetles in Serbian first year and continuous maize fields. J. Appl. Èntomol. 2009, 133, 430–437. [Google Scholar] [CrossRef]
- Boriani, M.; Agosti, M.; Kiss, J.; Edwards, C.R. Sustainable management of the western corn rootworm, Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae), in infested areas: Experiences in Italy, Hungary and the USA. Bull. OEPP/EPPO Bull. 2006, 36, 531–537. [Google Scholar] [CrossRef]
- Igrc-Barčić, J.; Dobrinčić, R.; Maceljski, M. The Spread and Population Density of the Western Corn Rootworm. In Proceedings of the Western Corn Rootworm 99: 4th FAO WCR/TCP Meeting of the EPPO ad hoc Panel and 6th International IWGO Workshop on Diabrotica virgifera virgifera LeConte, Paris, France, 4–5 November 1999. [Google Scholar]
- Ivezić, M.; Majić, I.; Raspudić, E.; Brmež, M.; Prakatur, B. The importance of Western corn rootworm in continuous maize. Poljoprivreda 2006, 12, 35–40. [Google Scholar]
- Ivezić, M.; Raspudić, E.; Brmež, M.; Pančić, S.; Majić, I. Implementation of pheromone traps in detecting click beetles population level in east Croatia. Cereal Res. Commun. 2007, 35, 513–516. [Google Scholar] [CrossRef]
- Husnjak, M.; Raspudić, E.; Brmež, M.; Majić, I.; Sarajlić, A. Comparison of Pferomone Traps with Yellow Sticky Traps in Monitoring Western Corn Rootworm in Virovitica–Podravina County. In Proceedings of the 7th International Scientific/Professional Conference, Agriculture in Nature and Environment, Vukovar, Croatia, 28–30 May 2014. [Google Scholar]
- Dobrinčić, R.; Igrc-Barčić, J. Istraživanje dinamike i gustoce populacije kukuruzne zlatice u Hrvatskoj. In Znanstveni Skup Hrvatskih Agronoma; Sažeci 37; Poljoprivredni Fakultet: Osijek, Croatia, 2001. [Google Scholar]
- Bažok, R.; Igrc-Barčić, J. Pheromone Applications in Maize Pest Control, 1st ed.; Novascience Publishers: Haupauge, NY, USA, 2010; pp. 23–35. ISBN 9781617286384. [Google Scholar]
- Maceljski, M.; Igrc-Barčić, J. Significance of Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) for Croatia. Poljopr. Znan. Smotra 1994, 59, 413–423. [Google Scholar]
- Maceljski, M.; Igrc-Barčić, J. Potential and Possible Damages of Diabrotica virgifera virgifera in Croatia. In Proceedings of the 1st Workshop on Diabrotica Virgifera, Graz, Austria, 20–21 March 1995. [Google Scholar]
- Žlof, V. Monitoring of Diabrotica virgifera virgifera LeConte in Croatia in 1996. In Proceedings of the IWGO Newsletter IWGO/EPPO Meeting, Zagreb, Croatia, 16–17 October 1996. [Google Scholar]
- Igrc-Barčić, J.; Maceljski, M. Kukuruzna zlatica (Diabrotica virgifera virgifera LeConte-Col.: Chrysomelidae)-novi štetnik u hrvatskom podunavlju. Agronomski Glasnik Glasilo Hrvatskog Agronomskog Društva 1997, 59, 429–443. [Google Scholar]
- Igrc-Barčić, J.; Maceljski, M. Establishment Potential of Diabrotica virgifera virgifera LeConte in Croatia. In Proceedings of the 3rd FAO WCR/TCP Meeting, 4th Meeting of the EPPO ad hoc PANEL, 5th International LeConteIWGO Workshop on Diabrotica virgifera virgifera LeConte, Rogaska Slatina, Slovenia, 28–29 October 1998. [Google Scholar]
- Igrc-Barčić, J.; Bažok, R.; Maceljski, M. Research on the western corn rootworm (Diabrotica virgifera virgifera LeConte, Coleoptera: Chrysomelidae) in Croatia (1994–2003). Èntomol. Croat. 2003, 7, 63–83. [Google Scholar]
- Shaw, J.T.; Ruesink, W.G.; Briggs, S.P.; Luckmann, W.H. Monitoring populations of corn rootworm beetle (Coleoptera: Crysomelidae) with trap bait with cucurbitacins. J. Econ. Entomol. 1984, 77, 1495–1499. [Google Scholar] [CrossRef]
- Csalomon. Traps Developed for Catching Diabrotica virgifera virgifera. Available online: http://www.csalomontraps.com/ (accessed on 16 October 2018).
- Great Lakes IPM. Insect Monitoring Sistem for the Professional Grower. Available online: https://www.greatlakesipm.com/cornrootwormtraps.html (accessed on 16 October 2018).
- Igrc-Barčić, J.; Bažok, R.; Edwards, C.R.; Kos, T. Western corn rootworm adult movement and possible egg laying in fields bordering maize. J. Appl. Èntomol. 2007, 131, 400–405. [Google Scholar] [CrossRef]
- Kos, T.; Bažok, R.; Varga, B.; Igrc-Barčić, J. Estimation of western corn rootworm (Diabrotica virgifera virgifera LeConte) egg abundance based on the previous year adult capture. J. Central Eur. Agric. 2013, 14. [Google Scholar] [CrossRef]
- Stauss, R. Compendium of Growth Stage Identification Keys for Mono and Dicotyledonous Plants, Extended BBCH Scale; Ciba-Geigy AG: Basel, Switzerland, 1994. [Google Scholar]
- Kiss, J.; Khosbayar, B.; Komaromi, J.; Igrc-Barčić, J.; Dobrinčić, R.; Sivčev, I.; Edwards, C.R.; Hatala-Zseller, I. Is the Western Corn Rootworm Adapting Itself to the European Crop Rotation System? In Proceedings of the XXI IWGO Conference, VIII Diabrotica Subgroup Meeting, Legnaro-Padua-Venice, Italy, 27 October–3 November 2001. [Google Scholar]
- Kos, T. Damage Forecast and Risk Assesment for Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). Ph.D. Thesis, Faculty of Agriculture, University of Zagreb, Zagreb, Croatia, 2011. [Google Scholar]
- Kozina, A. The Factors for the Temporal and Spatial Distribution of the Economically Important Maize pests. Ph.D. Thesis, Faculty of Agriculture, University of Zagreb, Zagreb, Croatia, 2012. [Google Scholar]
- Bažok, R. Overview on the distribution and the management of the WCR in Croatia. In Proceedings of the Workshop Organized in the FAO—Project framework Integrated Pest Management for Western Corn Rootworm in Central and Eastern Europe (GTFS/RER/017-ITA PROJECT), Zagreb, Croatia, 4–7 October 2006; pp. 52–55. [Google Scholar]
- Lemic, D.; Mikac, K.; Bažok, R.; Čačija, M. Genetic structure, gene flow and dispersal patterns of western corn rootworm Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) populations from Croatia. In Proceedings of the IX European Congress of Entomology, Budapest, Hungary, 22–27 August 2010. [Google Scholar]
- Lemic, D.; Mikac, K.; Bažok, R. Historical and contemporary genotypic changes associated with the invasion of Croatia by the WCR. In Proceedings of the 24th IWGO Conference and 3rd Internacional Conference of Diabrotica Genetics, Freiburg, Germany, 24–26 October 2011. [Google Scholar]
- Lemic, D.; Mikac, K.M.; Bažok, R. Historical and contemporary population genetics of the invasive western corn rootworm (Coleoptera: Chrysomelidae) in Croatia. Environ. Èntomol. 2013, 42, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Lemic, D.; Mikac, K.; Bažok, R.; Čačija, M. Gene flow and genetic structure among invasive western corn rootworm populations from Croatia. In Book of Abstracts, Proceedings of the 3rd Congress of Croatian Geneticist with International Participation, Krk, Croatia, 13–16 May 2012; Franekić, J., Garaj-Vrhovac, V., Eds.; Croatian Genetic Society: Zagreb, Croatia, 2012; p. 90. [Google Scholar]
- Lemic, D.; Mikac, K.; Benitez, H.; Bažok, R.; Buketa, M. Diabrotica virgifera virgifera LeConte wing shape variation reveals multiple populations across the European expansion front. In Proceedings of the Abstracts of Lectures and Posters: International Conference on the German Diabrotica Research Program, Bonn, Federal Ministry of Food, Agriculture and Consumer Protection, Berlin, Germany, 14–17 November 2012; p. 9. [Google Scholar]
- Ivkosic, S.A.; Gorman, J.; Lemic, D.; Mikac, K.M. Genetic monitoring of western corn rootworm (Coleoptera: Chrysomelidae) populations on a microgeographic scale. Environ. Èntomol. 2014, 43, 804–818. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Sappington, T.W. Genetic structuring of western corn rootworm (Coleoptera: Chrysomelidae) populations in the U.S. based on microsatellite loci analysis. Environ. Entomol. 2005, 34, 494–503. [Google Scholar] [CrossRef]
- Lemic, D.; Mikac, K.M.; Ivkosic, S.A.; Bažok, R. The temporal and spatial invasion genetics of the western corn rootworm (Coleoptera: Chrysomelidae) in southern Europe. PLoS ONE 2015, 10, e0138796. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Stolz, U.; Miller, N.J.; Waits, E.; Guillemaud, T.; Sumerford, D.V. A core set of microsatellitemmarkers for western corn rootworm (Coleoptera: Chrysomelidae) population genetics studies. Environ. Entomol. 2008, 37, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Lemic, D.; Benítez, H.A.; Bažok, R. Intercontinental effect on sexual shape dimorphism and allometric relationships in the beetle pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Zool. Anz. 2014, 253, 203–206. [Google Scholar] [CrossRef]
- Lemic, D.; Mikac, K.; Benitez, H.; Bažok, R. Innovative and modern monitoring techniques–essential tool for effective pest control management. In Proceedings of the 8th CASEE Conference, Warsaw University of Life Sciences–SGGW, Warsaw, Poland, 14–16 May 2017. [Google Scholar]
- Mikac, K.M.; Douglas, J.; Spencer, J.L. Wing shape and size of the western corn rootworm (Coleoptera: Chrysomelidae) is related to sex and resistance to soybean-maize crop rotation. J. Econ. Entomol. 2013, 106, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Mikac, K.M.; Lemic, D.; Bažok, R. A decade of populations genetics and geometric morphometrics research on the western corn rootworm in Southern Europe: What have we learned and where to from here? In Proceedings of the European Congress of Entomology, ECE, Napoli, Italy, 2–6 July 2018. [Google Scholar]
- Benítez, H.A.; Lemic, D.; Bažok, R.; Gallardo-Araya, C.M.; Mikac, K.M. Evolutionary directional asymmetry and shape variation in Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae): An example using hind wings. Boil. J. Linn. Soc. 2013, 111, 110–118. [Google Scholar] [CrossRef]
- Benítez, H.A.; Lemic, D.; Bažok, R.; Bravi, R.; Buketa, M.; Püschel, T. Morphological integration and modularity in Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) hind wings. Zool. Anz. 2014, 253, 461–468. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Bažok, R.; Benítez, H.A. Wing shape changes: A morphological view of the Diabrotica virgifera virgifera European invasion. Boil. Invasions 2016, 18, 3401–3407. [Google Scholar] [CrossRef]
- Gidaszewski, N.A.; Baylac, M.; Klingenberg, C.P. Evolution of sexual dimorphism of wing shape in the Drosophila melanogaster subgroup. BMC Evol. Boil. 2009, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Shingleton, A.W.; Mirth, C.K.; Bates, P.W. Developmental model of static allometry in holometabolous insects. Proc. R. Soc. B Boil. Sci. 2008, 275, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingenberg, C.P. Morphological Integration and Developmental Modularity. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 115–132. [Google Scholar] [CrossRef]
- Coates, B.S.; Sumerford, D.V.; Miller, N.J.; Kim, K.S.; Sappington, T.W.; Siegfried, B.D.; Lewis, L.C. Comparative performance of single nucleotide polymorphism and microsatellite markers for population genetic analysis. J. Hered. 2009, 100, 556–564. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrganić, M.; Bažok, R.; Mikac, K.M.; Benítez, H.A.; Lemic, D. Two Decades of Invasive Western Corn Rootworm Population Monitoring in Croatia. Insects 2018, 9, 160. https://doi.org/10.3390/insects9040160
Mrganić M, Bažok R, Mikac KM, Benítez HA, Lemic D. Two Decades of Invasive Western Corn Rootworm Population Monitoring in Croatia. Insects. 2018; 9(4):160. https://doi.org/10.3390/insects9040160
Chicago/Turabian StyleMrganić, Martina, Renata Bažok, Katarina M. Mikac, Hugo A. Benítez, and Darija Lemic. 2018. "Two Decades of Invasive Western Corn Rootworm Population Monitoring in Croatia" Insects 9, no. 4: 160. https://doi.org/10.3390/insects9040160
APA StyleMrganić, M., Bažok, R., Mikac, K. M., Benítez, H. A., & Lemic, D. (2018). Two Decades of Invasive Western Corn Rootworm Population Monitoring in Croatia. Insects, 9(4), 160. https://doi.org/10.3390/insects9040160