Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Organism
2.2. Experimental Colonies
2.3. Retrieval of Intact Pupae
2.4. Activity of Pupal Extracts
2.5. Activity of Fipronil-Treated Ant Cadaver Extracts
2.6. Response Variables
2.7. Statistical Analysis
2.7.1. Activity of Pupal Extracts
2.7.2. Activity of Fipronil-Treated Ant Cadaver Extracts
3. Results
3.1. Pupa Retrieval Time
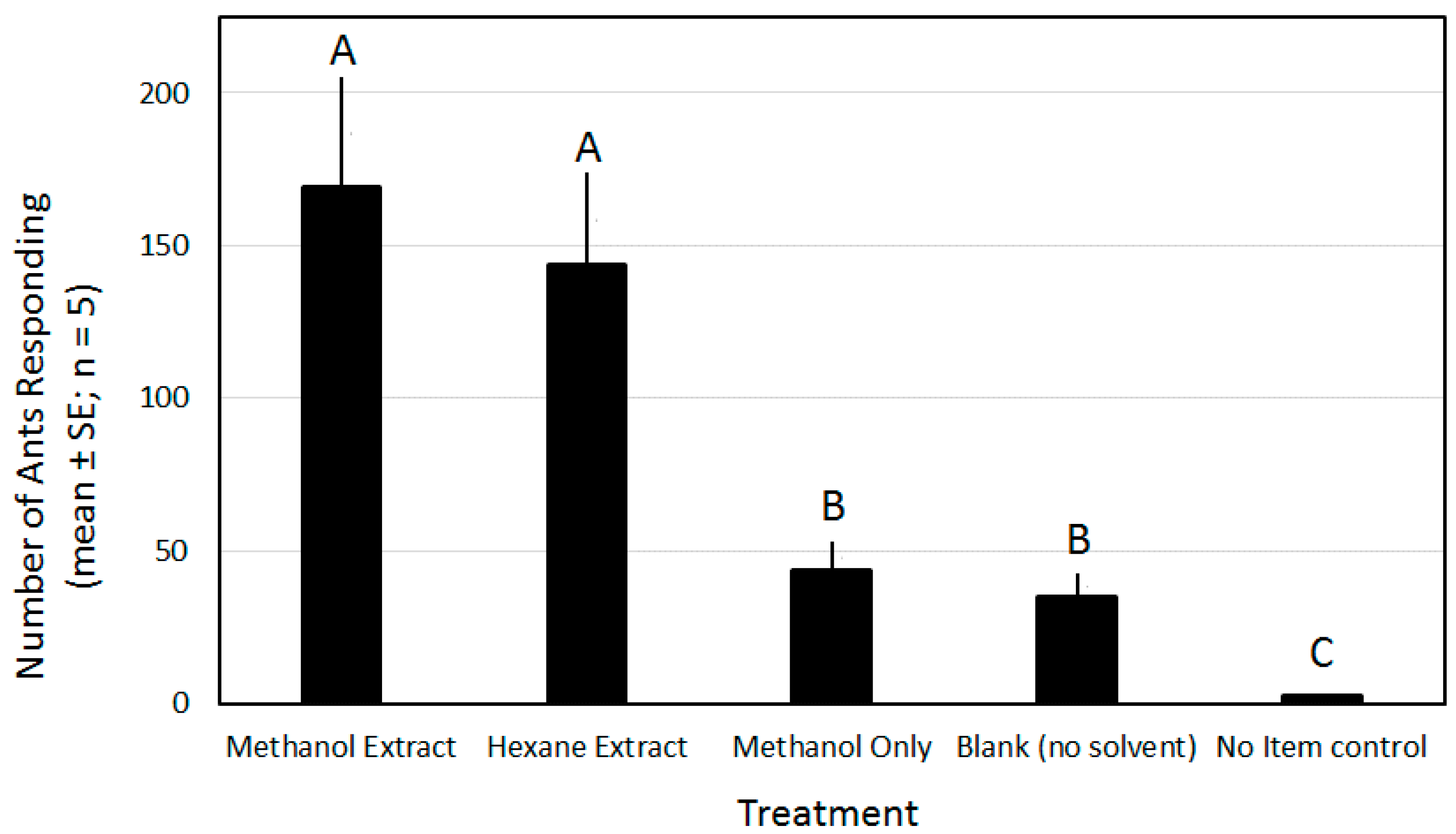
3.2. Activity of Pupal Extracts
3.3. Activity of Fipronil-Treated Ant Cadaver Extracts
4. Discussion
4.1. Pupa Retrieval Time
4.2. Activity of Pupal Extracts
4.3. Activity of Fipronil-Treated Ant Cadaver Extracts
Author Contributions
Funding
Conflicts of Interest
References
- Vega, S.J.; Rust, M.K. The Argentine ant—A significant invasive species in agricultural, urban and natural environments. Sociobiology 2001, 37, 3–25. [Google Scholar]
- Suarez, A.V.; Holway, D.A.; Tsutsui, N.D. Genetics and behavior of a colonizing species: The invasive Argentine ant. Am. Nat. 2008, 172, S72–S84. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, G.M.; Silverman, J. Queen acceptance and the complexity of nestmate discrimination in the Argentine ant. Behav. Ecol. Sociobiol. 2008, 62, 537–548. [Google Scholar] [CrossRef]
- Wiltz, B.A.; Suiter, D.R.; Gardner, W.A. Activity of bifenthrin, chlorfenapyr, fipronil, and thiamethoxam against Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2009, 102, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Reierson, D.A.; Klotz, J.H. Pest management of Argentine ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2003, 38, 159–169. [Google Scholar] [CrossRef]
- Choe, D.H.; Rust, M.K. Horizontal transfer of insecticides in laboratory colonies of the Argentine ant (Hymenoptera: Formicidae). J. Econ. Entomol. 2008, 101, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Hooper-Bui, L.M.; Rust, M.K. Oral toxicity of abamectin, boric acid, fipronil, and hydramethylnon to laboratory colonies of Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2000, 93, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Hooper-Bui, L.M.; Kwok, E.S.C.; Buchholz, B.A.; Rust, M.K.; Eastmond, D.A.; Vogel, J.S. Insecticide transfer efficiency and lethal load in Argentine ants. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2015, 361, 665–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, M.; Reierson, D.A.; Klotz, J.H. Delayed toxicity as a critical factor in the efficacy of aqueous baits for controlling Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2004, 97, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.; Reierson, D.A.; Klotz, J.H. Factors affecting the performance of bait toxicants for Argentine ants (Hymenoptera: Formicidae). In Proceedings of the 4th International Conference on Urban Pests, Charleston, SC, USA, 7–10 July 2002; Jones, S.C., Zhai, J., Robinson, W.H., Eds.; Pocahontas Press, Inc.: Blacksburg, VA, USA, 2002; pp. 115–120. [Google Scholar]
- Vega, S.Y.; Rust, M.K. Determining the foraging range and origin of resurgence after treatment of Argentine ant (Hymenoptera: Formicidae) in urban areas. J. Econ. Entomol. 2003, 96, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.H.; Rust, M.K.; Costa, H.S.; Reierson, D.A.; Kido, K. Strategies for controlling Argentine ants (Hymenoptera: Formicidae) with sprays and baits. J. Agric. Urban Entomol. 2002, 19, 85–94. [Google Scholar]
- Gan, J.; Bondarenko, S.; Oki, L.; Haver, D.; Li, J.X. Occurrence of fipronil and its biologically active derivatives in urban residential runoff. Environ. Sci. Technol. 2012, 46, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Y.; Soeprono, A.; Rust, M.K.; Gan, J. Ant control efficacy of pyrethroids and fipronil on outdoor concrete surfaces. Pest Manag. Sci. 2014, 70, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, J.P.; Rouse, D.R.; Avants, J.K.; Garrison, A.W.; Delorenzo, M.E.; Chung, K.W.; Key, P.B.; Wilson, W.A.; Black, M.C. Toxicity of fipronil and its enantiomers to marine and freshwater non-targets. J. Environ. Sci. Health Part B 2007, 42, 471–480. [Google Scholar] [CrossRef]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, L.; Rust, M.K.; Klotz, J.H.; Haver, D.; Kabashima, J.N.; Bondarenko, S.; Gan, J. Impact of ant control technologies on insecticide runoff and efficacy. Pest Manag. Sci. 2010, 66, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Wiltz, B.A.; Suiter, D.R.; Gardner, W.A. Deterrency and toxicity of essential oils to Argentine and Red Imported Fire ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2007, 42, 239–249. [Google Scholar] [CrossRef]
- Guerra, M.D.; Suiter, D.R.; Scocco, C.M. Topical toxicity of nine essential oils to Camponotus pennsylvanicus (Hymenoptera: Formicidae). Sociobiology 2011, 58, 419–426. [Google Scholar]
- Scocco, C.M.; Suiter, D.R.; Gardner, W.A. Repellency of five essential oils to Linepithema humile (Hymenoptera: Formicidae). J. Entomol. Sci. 2012, 47, 150–159. [Google Scholar] [CrossRef]
- Greenberg, L.; Klotz, J.H. Argentine ant (Hymenoptera: Formicidae) trail pheromone enhances consumption of liquid sucrose solution. J. Econ. Entomol. 2000, 93, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Tsai, K.; Lopez, C.M.; Campbell, K. Pheromone-assisted techniques to improve the efficacy of insecticide sprays against Linepithema humile (Hymenoptera: Formicidae). J. Econ. Entomol. 2014, 107, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Peck, R.W.; Manning, L.M.; Stringer, L.D.; Cappadonna, J.; El-Sayed, A.M. Pheromone disruption of Argentine ant trail integrity. J. Chem. Ecol. 2008, 34, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Peck, R.W.; Stringer, L.D.; Snook, K.; Banko, P.C. Trail pheromone disruption of Argentine ant trail formation and foraging. J. Chem. Ecol. 2010, 36, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Stringer, L.D.; Corn, J.E. Argentine ant trail pheromone disruption is mediated by trail concentration. J. Chem. Ecol. 2011, 37, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.H.; Rust, M.K.; Greenberg, L.; Robertson, M.A. Developing low risk management strategies for Argentine ants (Hymenoptera: Formicidae). Sociobiology 2010, 55, 779–785. [Google Scholar]
- Buczkowski, G. Prey-baiting as a conservation tool: Selective control of invasive ants with minimal non-target effects. Insect Conserv. Divers. 2017, 10, 302–309. [Google Scholar] [CrossRef]
- Hughes, L.; Westoby, M. Capitula on stick insect eggs and elaiosomes on seeds: Convergent adaptations for burial by ants. Funct. Ecol. 1992, 6, 642–648. [Google Scholar] [CrossRef]
- Windsor, D.M.; Trapnell, D.W.; Amat, G. The egg capitulum of a Neotropical walkingstick, Calynda bicuspis, induces aboveground egg dispersal by the ponerine ant, Ectatomma ruidum. J. Insect Behav. 1996, 9, 353–367. [Google Scholar] [CrossRef]
- Brew, C.R.; O’Dowd, D.J.; Rae, I.D. Seed dispersal by ants: Behaviour-releasing compounds in elaiosomes. Oecologia 1989, 80, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Millar, J.G.; Rust, M.K. Chemical signals associated with life inhibit necrophoresis in Argentine ants. Proc. Natl. Acad. Sci. USA 2009, 106, 8251–8255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS Institute Inc. SAS/STAT Software Version 14.1; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Walsh, J.P.; Tschinkel, W.R. Brood recognition by contact pheromone in the red imported fire ant, Solenopsis invicta. Anim. Behav. 1974, 22, 695–704. [Google Scholar] [CrossRef]
- Bonavitacougourdan, A.; Clement, J.L.; Lange, C. Larvae recognition in the Camponotus vagus Scop ant—Larvae phenotypes of cuticular hydrocarbon patterns. C. R. Acad. Sci. Ser. Sci. Vie-Life Sci. 1988, 306, 299–305. [Google Scholar]
- Bonavitacougourdan, A.; Clement, J.L.; Lange, C. The role of cuticular hydrocarbons in recognition of larvae by workers of the ant Camponotus vagus—Changes in the chemical signature in response to social-environment (Hymenoptera: Formicidae). Sociobiology 1989, 16, 49–74. [Google Scholar]
- Van Wilgenburg, E.; Sulc, R.; Shea, K.J.; Tsutsui, N.D. Deciphering the chemical basis of nestmate recognition. J. Chem. Ecol. 2010, 36, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Shimoji, H.; Fujiki, Y.; Yamaoka, R.; Tsuji, K. Egg discrimination by workers in Diacamma sp from Japan. Insectes Soc. 2012, 59, 201–206. [Google Scholar] [CrossRef]
- Helantera, H.; Martin, S.J.; Ratnieks, F.L.W. Recognition of nestmate eggs in the ant Formica fusca is based on queen derived cues. Curr. Zool. 2014, 60, 131–136. [Google Scholar] [CrossRef]
- Krasnec, M.O.; Breed, M.D. Colony-specific cuticular hydrocarbon profile in Formica argentea ants. J. Chem. Ecol. 2013, 39, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sainz-Borgo, C.; Cabrera, A.; Hernandez, J.V. Nestmate recognition in the ant Odontomachus bauri (Hymenoptera: Formicidae). Sociobiology 2011, 58, 701–718. [Google Scholar]
- Bigley, W.S.; Vinson, S.B. Characterization of a brood pheromone isolated from the sexual brood of the imported fire ant, Solenopsis invicta. Ann. Entomol. Soc. Am. 1975, 68, 301–304. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Nestmate and brood recognition. In The Fire Ants; Belknap: Cambridge, MA, USA, 2006; pp. 263–268. ISBN 0-674-02207-6. [Google Scholar]
- Wiltz, B.A.; Suiter, D.; Gardner, W.; Berisford, C.W. A novel delivery method for ant (Hymenoptera: Formicidae) toxicants. Midsouth Entomol. 2010, 3, 79–88. [Google Scholar]
- Glancey, B.M.; Stringer, C.E.; Craig, C.H.; Bishop, P.M.; Martin, B.B. Pheromone may induce brood tending in the fire ant, Solenopsis saevissima. Nature 1970, 226, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.L.; Beattie, A.J.; Bollenbacher, W.E. Evidence for diglycerides as attractants in an ant-seed interaction. J. Chem. Ecol. 1979, 5, 335–344. [Google Scholar] [CrossRef]
- Buczkowski, G. The Trojan horse approach for managing invasive ants: A study with Asian needle ants, Pachycondyla chinensis. Biol. Invasions 2016, 18, 507–515. [Google Scholar] [CrossRef]
- Buczkowski, G.; Mothapo, N.P.; Wossler, T.C. Let them eat termites—Prey-baiting provides effective control of Argentine ants, Linepithema humile, in a biodiversity hotspot. J. Appl. Entomol. 2018, 142, 504–512. [Google Scholar] [CrossRef]
- Pulliainen, U.; Bos, N.; d’Errorre, P.; Sundström, L. Caste-dependent brood retrieval by workers in the ant Formica exsecta. Anim. Behav. 2018, 140, 151–159. [Google Scholar] [CrossRef]
- Mahler, B.J.; Van Metre, P.C.; Wilson, J.T.; Musgrove, M.; Zaugg, S.D.; Burkhardt, M.R. Fipronil and its degradates in indoor and outdoor dust. Environ. Sci. Technol. 2009, 43, 5665–5670. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, D.; Kuwayama, N.; Takeo, A.; Ishida, T.; Mano, H.; Inoue, M.N.; Nagai, T.; Sanchez-Bayo, F.; Goka, K.; Sawahata, T. Different acute toxicity of fipronil baits on invasive Linepithema humile supercolonies and some non-target ground arthropods. Ecotoxicology 2015, 24, 1221–1228. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Hour 1 | Hour 2 | Hour 24 |
|---|---|---|---|
| Ant Cadavers | 26.1 ± 2.6 a | 27.8 ± 2.2 a | 30.0 ± 2.2 a |
| 0.001% fipronil + cadavers | 26.8 ± 2.6 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.01% fipronil + cadavers | 26.8 ± 2.6 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.10% fipronil + cadavers | 25.9 ± 2.5 a | 27.8 ± 2.2 a | 30.0 ± 2.2 a |
| 0.001% fipronil + cadaver extract + wicks | 27.7 ± 2.7 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.01% fipronil + cadaver extract + wicks | 29.6 ± 2.8 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.10% fipronil + cadaver extract + wicks | 25.9 ± 2.5 a | 26.5 ± 2.2 a | 27.0 ± 2.1 a |
| 0.001% fipronil + wicks | 0.7 ± 0.3 b | 0.8 ± 0.4 c | 7.0 ± 1.1 c |
| 0.01% fipronil + wicks | 1.5 ± 0.5 b | 2.0 ± 0.6 bc | 4.8 ± 0.9 c |
| 0.10% fipronil + wicks | 1.7 ± 0.5 b | 2.7 ± 0.7 b | 13.7 ± 1.5 b |
| Treatment | Percent Mortality (Mean ± SE) |
|---|---|
| Ant cadavers | 15.1 ± 4.3 g |
| 0.001% fipronil + cadavers | 19.3 ± 1.2 f |
| 0.01% fipronil + cadavers | 58.9 ± 7.3 b |
| 0.10% fipronil + cadavers | 84.9 ± 4 a |
| 0.001% fipronil + cadaver extract + wicks | 14.0 ± 2.1 g |
| 0.01% fipronil + cadaver extract + wicks | 13.5 ± 1.5 g |
| 0.10% fipronil + cadaver extract + wicks | 43.9 ± 7.9 d |
| 0.001% fipronil + wicks | 18.3 ± 6.2 f |
| 0.01% fipronil + wicks | 25.9 ± 5.1 e |
| 0.10% fipronil + wicks | 51.9 ± 5.9 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gochnour, B.M.; Suiter, D.R.; Davis, J.W.; Huang, Q. Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects 2018, 9, 171. https://doi.org/10.3390/insects9040171
Gochnour BM, Suiter DR, Davis JW, Huang Q. Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects. 2018; 9(4):171. https://doi.org/10.3390/insects9040171
Chicago/Turabian StyleGochnour, Benjamin M., Daniel R. Suiter, Jerry W. Davis, and Qingguo Huang. 2018. "Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant" Insects 9, no. 4: 171. https://doi.org/10.3390/insects9040171
APA StyleGochnour, B. M., Suiter, D. R., Davis, J. W., & Huang, Q. (2018). Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects, 9(4), 171. https://doi.org/10.3390/insects9040171