Larval Food Limitation in a Speyeria Butterfly (Nymphalidae): How Many Butterflies Can Be Supported?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
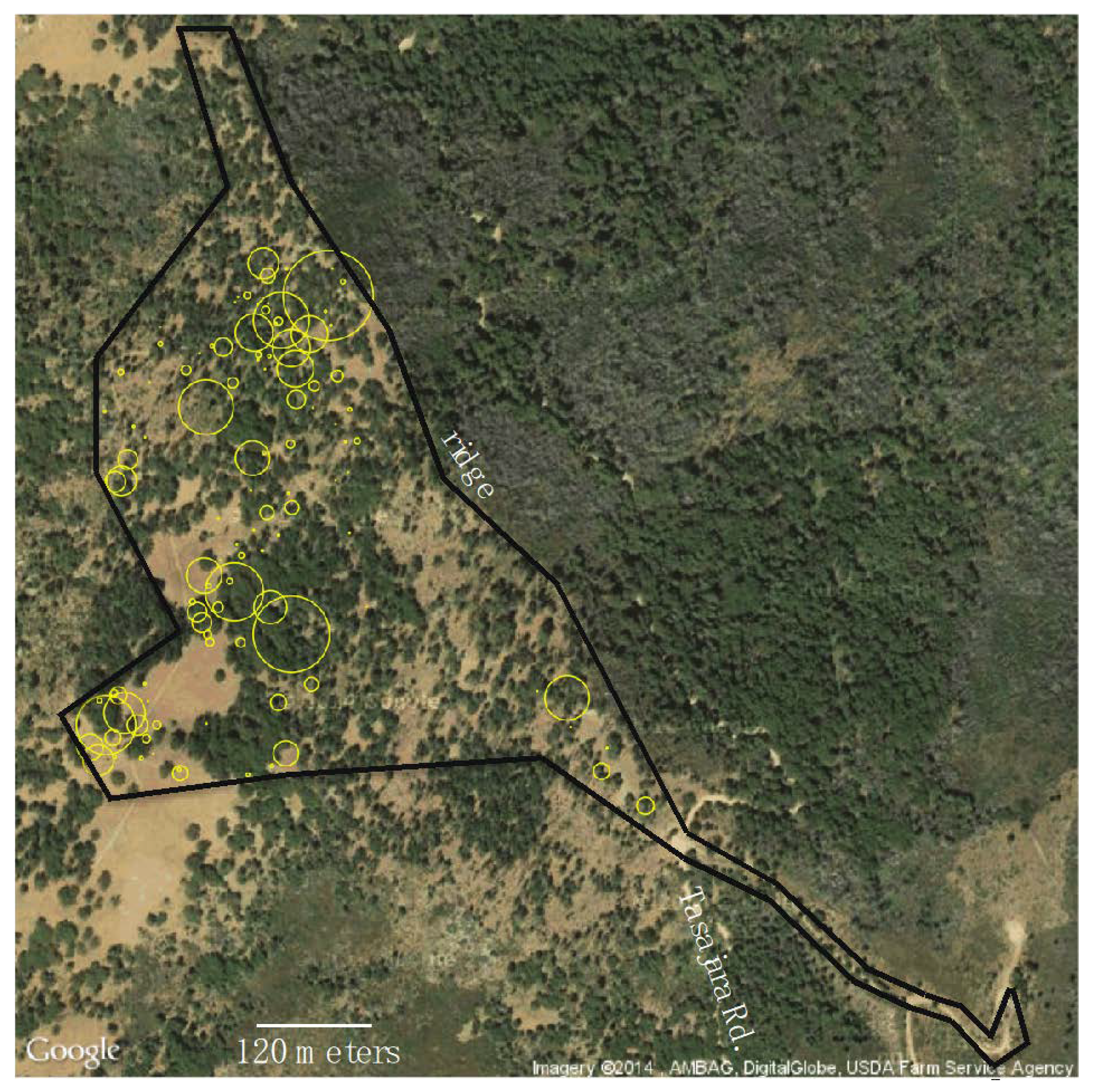
2.2. Abundance of Adult Butterflies and Food Plant
2.3. Leaf Area Consumption
2.4. Model Fitting and Simulation Analysis
3. Results
3.1. Viola Purpurea Quercetorum Abundance, Density and Amount of Leaf Present on Chews Ridge
3.2. Food Plant Consumption in Lab
3.3. Simulation and Comparison of Estimated Number Pupae to Adult Population Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
- dtnorm2<- function(x, mean, sd, a, b) {
- dnorm(x, mean, sd)/(pnorm(b, mean, sd)-pnorm(a, mean, sd))
- }
- ptnorm <- function(x, mean, sd, a, b) {
- (pnorm(x,mean,sd) - pnorm(a,mean,sd)) /
- (pnorm(b,mean,sd) - pnorm(a,mean,sd))
- }
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distribution | Likelihood | AIC |
|---|---|---|
| log-normal | −778.7 | 1561.5 |
| gamma | −780.2 | 1564.4 |
| weibull | −784.2 | 1572.3 |
| trunc-normal | −793.5 | 1590.9 |
| normal | −841.0 | 1686.0 |
| Distribution | Likelihood | AIC |
|---|---|---|
| gamma | −2107.8 | 4219.5 |
| weibull | −2108.5 | 4221.0 |
| trunc-normal | −2119.5 | 4242.9 |
| log-normal | −2122.9 | 4249.7 |
| normal | −2138.3 | 4280.6 |
| Distribution | Likelihood | AIC |
|---|---|---|
| weibull | 39.3 | −74.6 |
| normal | 38.7 | −73.4 |
| trunc-normal | 38.7 | −73.4 |
| gamma | 38.3 | −72.6 |
| log-normal | 38.0 | −72.0 |
References
- Wilkinson, D.M.; Sherratt, T.N. Why is the world green? The interactions of top–down and bottom–up processes in terrestrial vegetation ecology. Plant Ecol. Divers. 2016, 9, 127–140. [Google Scholar] [CrossRef]
- Hairston, N.G.; Smith, F.E.; Slobodkin, L.B. Community structure, population control and competition. Am. Nat. 1960, 94, 421–425. [Google Scholar] [CrossRef]
- Power, M.E. Top-down and bottom-up forces in food webs: Do plants have primacy? Ecology 1992, 73, 733–746. [Google Scholar] [CrossRef]
- Hunter, M.D.; Price, P.W. Chutes and ladders: Heterogeneity and the relative roles of bottom-up and top-down forces in natural communities. Ecology 1992, 73, 724–732. [Google Scholar]
- Vidal, M.C.; Murphy, S.M. Bottom-up vs. top-down effects on terrestrial insect herbivores: A meta-analysis. Ecol. Lett. 2018, 21, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Marini, L.; Zalucki, M.P. Density-dependence in the declining population of the monarch butterfly. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.B.; Dlugosch, K.M. Nectar and hostplant scarcity limit populations of an endangered Oregon butterfly. Oecologia 1999, 119, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Debinski, D.M. Relationship of host plant density to size and abundance of the regal fritillary Speyeria idalia Drury (Nymphalidae). J. Lepid Soc. 1998, 52, 262–276. [Google Scholar]
- Curtis, R.J.; Brereton, T.M.; Dennis, R.L.H.; Carbone, C.; Isaac, N.J.B. Butterfly abundance is determined by food availability and is mediated by species traits. J. Appl. Ecol. 2015, 52, 1676–1684. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A. Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 339–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.A. Butterfly communities under threat. Science 2016, 353, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.C.; Segerer, A.; Ulrich, W.; Torchyk, O.; Weisser, W.W.; Schmitt, T. Butterfly community shifts over two centuries. Conserv. Biol. 2016, 30, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.S.; Freitas, A.V.L. Atlantic Forest Butterflies: Indicators for Landscape Conservation. Biotropica 2000, 32, 934–956. [Google Scholar] [CrossRef]
- Swengel, S.R.; Schlicht, D.; Olsen, F.; Swengel, A.B. Declines of prairie butterflies in the midwestern USA. J. Insect Conserv. 2011, 15, 327–339. [Google Scholar] [CrossRef]
- Sands, D.P.A.; New, T.R. Birdwing butterflies and their conservation needs. In Conservation of the Richmond Birdwing Butterfly in Australia; Sands, D.P.A., New, T.R., Eds.; Spring: Dordrecht, The Netherlands, 2013. [Google Scholar]
- New, T.R. Insect Conservation: Past, Present and Prospects; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Agrawal, A.A.; Inamine, H. Mechanisms behind the monarch’s decline. Science 2018, 360, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Hammond, P.C.; McCorkle, D.V. The decline and extinction of Speyeria populations resulting from human environmental disturbances (Nymphalidae: Argynninae). J. Res. Lepid. 1983, 22, 217–224. [Google Scholar]
- Wells, C.N.; Tonkyn, D.W. Range collapse in the Diana fritillary, Speyeria diana (Nymphalidae). Insect Conserv. Divers. 2014, 7, 365–380. [Google Scholar] [CrossRef]
- Wells, C.; Tonkyn, D. Changes in the geographic distribution of the Diana fritillary (Speyeria diana: Nymphalidae) under forecasted predictions of climate change. Insects 2018, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- McCorkle, D.V.; Hammond, P.C. Biology of Speyeria zerene hippolyta (Nymphalidae) in a marine-modified environment. J. Lepid. Soc. 1988, 42, 184–195. [Google Scholar]
- Davenport, K. Lepidoptera of North. America 15. Butterflies of Southern California in 2018: Updating Emmel and Emmel’s 1973 Butterflies of southern California; Contributions of the C.P. Gillette Museum of Arthropod Diversity Colorado State University: Fort Collins, CO, USA, 2018; Volume 15, p. 175. [Google Scholar]
- Zaman, K.; Tenney, C.; Brunell, M.; Chen, M.; Hill, R.I. Life history and ecology of Speyeria adiaste clemencei (Comstocki, 1925) (Lepidoptera: Nymphalidae). J. Res. Lepid. 2014, 47, 29–43. [Google Scholar]
- Zaman, K.; Tenney, C.; Rush, C.E.; Hill, R.I. Population ecology of a California endemic: Speyeria adiaste clemencei. J. Insect Conserv. 2015, 19, 753–763. [Google Scholar] [CrossRef]
- Longcore, T.; Lam, C.S.; Kobernus, P.; Polk, E.; Wilson, J.P. Extracting useful data from imperfect monitoring schemes: Endangered butterflies at San Bruno Mountain, San Mateo County, California (1982–2000) and implications for habitat management. J. Insect Conserv. 2010, 14, 335–346. [Google Scholar] [CrossRef]
- Longcore, T.; Osborne, K.H. Butterflies are not Grizzly Bears: Lepidoptera conservation in practice. In Butterfly Conservation in North America; Daniels, J.C., Ed.; Springer: New York, NY, USA, 2015; pp. 161–192. [Google Scholar]
- McCullough, K.; Albanese, G.; Haukos, D.A. Novel observation of larval fire survival, feeding behavior, and host plant use in the Regal Fritillary, Speyeria idalia (Drury)(Nymphalidae). J. Lepid. Soc. 2017, 71, 146–152. [Google Scholar] [CrossRef]
- Rudolf, D.C.; Ely, C.A.; Schaefer, R.R.; Williamson, J.H.; Thill, R.E. The Diana Fritillary (Speyeria diana) and Great Spangled Fritillary (S. cybele): Dependence on fire in the Ouachita Mountains of Arkansas. J. Lepid. Soc. 2006, 60, 218–226. [Google Scholar]
- Sims, S.R. Speyeria (Lepidoptera: Nymphalidae) Conservation. Insects 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Bierzychudek, P.; Warner, K.A.; McHugh, A.; Thomas, L. Testing the host-finding ability of a monophagous caterpillar in the field. Ecol. Entomol. 2009, 34, 632–637. [Google Scholar] [CrossRef]
- Bierzychudek, P.; Warner, K. Modeling caterpillar movement to guide habitat enhancement for Speyeria zerene hippolyta, the Oregon silverspot butterfly. J. Insect Conserv. 2015, 19, 45–54. [Google Scholar] [CrossRef]
- Comstock, J.A. Butterflies of California; Scientific Publishers: Los Angeles, CA, USA, 1927; p. 334. [Google Scholar]
- Emmel, T.C.; Emmel, J.F. The Butterflies of Southern California; Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1973; Volume 26, pp. 1–148. [Google Scholar]
- Shapiro, A.M.; Manolis, T.D. Field Guide to Butterflies of the San Francisco Bay and Sacramento Valley Regions; University of California Press: Berkeley, CA, USA, 2007. [Google Scholar]
- Roderick, M.L.; Cochrane, M.J. On the conservative nature of the leaf mass–area relationship. Ann. Bot. 2002, 89, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at statistical model identification. IEEE Trans. Automat. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- R_Development_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- James, D.G.; Nunnallee, D. Life Histories of Cascadia Butterflies; Oregon State University Press: Corvallis, OR, USA, 2011; p. 447. [Google Scholar]
- Scott, J.A. The Butterflies of North. America: A Natural History and Field Guide; Stanford University Press: Stanford, CA, USA, 1986; p. 583. [Google Scholar]
- Wells, C.N.; Edwards, L.; Hawkins, R.; Smith, L.; Tonkyn, D. A rearing method for Argynnis (Speyeria) diana (Lepidoptera: Nymphalidae) that avoids larval diapause. Psyche 2011, 2011, 940280. [Google Scholar]
- James, D.G. Comparative studies on the immature stages and developmental biology of five species of Argynnis spp. (Subgenus Speyeria) (Nymphalidae) from Washington. J. Lepid. Soc. 2008, 62, 61–66. [Google Scholar]
- Roslin, T.; Syrjälä, H.; Roland, J.; Harrison, P.J.; Fownes, S.; Matter, S.F. Caterpillars on the run—Induced defences create spatial patterns in host plant damage. Ecography 2008, 31, 335–347. [Google Scholar] [CrossRef]
- Rohwer, C.L.; Erwin, J.E. Spider mites (Tetranychus urticae) perform poorly on and disperse from plants exposed to methyl jasmonate. Entomol. Exp. Appl. 2010, 137, 143–152. [Google Scholar] [CrossRef]
- Gruber, C.W.; Elliott, A.G.; Ireland, D.C.; Delprete, P.G.; Dessein, S.; Göransson, U.; Trabi, M.; Wang, C.K.; Kinghorn, A.B.; Robbrecht, E.; et al. Distribution and evolution of circular miniproteins in flowering plants. Plant. Cell 2008, 20, 2471–2483. [Google Scholar] [CrossRef] [PubMed]
- Göransson, U.; Luijendijk, T.; Johansson, S.; Bohlin, L.; Claeson, P. Seven novel macrocyclic polypeptides from Viola arvensis. J. Nat. Prod. 1999, 62, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Pränting, M.; Lööv, C.; Burman, R.; Göransson, U.; Andersson, D.I. The cyclotide cycloviolacin O2 from Viola odorata has potent bactericidal activity against Gram-negative bacteria. J. Antimicrob. Chemother. 2010, 65, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Schöpke, T.; Hasan Agha, M.I.; Kraft, R.; Otto, A.; Hiller, K. Hämolytisch aktive komponenten aus Viola tricolor L. und Viola arvensis Murray. Sci. Pharm. 1993, 61, 145–153. [Google Scholar]
- Tang, J.; Wang, C.K.; Pan, X.; Yan, H.; Zeng, G.; Xu, W.; He, W.; Daly, N.L.; Craik, D.J.; Tan, N. Isolation and characterization of cytotoxic cyclotides from Viola tricolor. Peptides 2010, 31, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Hashempour, H.; Ghassempour, A.; Daly, N.L.; Spengler, B.; Rompp, A. Analysis of cyclotides in Viola ignobilis by nano liquid chromatography fourier transform mass spectrometry. Protein Pept. Lett. 2011, 18, 747–752. [Google Scholar] [CrossRef]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar] [CrossRef] [PubMed]
- Barbeta, B.L.; Marshall, A.T.; Gillon, A.D.; Craik, D.J.; Anderson, M.A. Plant cyclotides disrupt epithelial cells in the midgut of lepidopteran larvae. Proc. Natl. Acad. Sci. USA 2008, 105, 1221–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, C.V.; Rosengren, K.J.; Daly, N.L.; Plan, M.; Stevens, J.; Scanlon, M.J.; Waine, C.; Norman, D.G.; Anderson, M.A.; Craik, D.J. Isolation, solution structure, and insecticidal activity of kalata B2, a circular protein with a twist: do Möbius strips exist in nature? Biochemistry 2005, 44, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Boggs, C.L.; Ross, C.L. The effect of adult food limitation on life-history traits in Speyeria mormonia (Lepidoptera, Nymphalidae). Ecology 1993, 74, 433–441. [Google Scholar] [CrossRef]
- García-Barros, E. Body size, egg size, and their interspecific relationships with ecological and life history traits in butterflies (Lepidoptera: Papilionoidea, Hesperioidea). Biol. J. Linn. Soc. 2000, 70, 251–284. [Google Scholar] [CrossRef] [Green Version]
- Boggs, C.L. Reproductive strategies of female butterflies: Variation in and constraints on fecundity. Ecol. Entomol. 1986, 11, 7–15. [Google Scholar] [CrossRef]
- Wolfe, K. Natural non-Viola hostplant of a sierran Speyeria (Nymphalidae, Heliconiinae) and an associated parasitoid (Diptera, Tachinidae). J. Lepid. Soc. 2017, 71, 129–131. [Google Scholar] [CrossRef]
- Arnaud, P.H. A Host-Parasite Catalog of North American Tachinidae (Diptera); Miscellaneous Publication No. 1319; United States Department of Agriculture: Washington, DC, USA, 1978; pp. 1–860.




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, R.I.; Rush, C.E.; Mayberry, J. Larval Food Limitation in a Speyeria Butterfly (Nymphalidae): How Many Butterflies Can Be Supported? Insects 2018, 9, 179. https://doi.org/10.3390/insects9040179
Hill RI, Rush CE, Mayberry J. Larval Food Limitation in a Speyeria Butterfly (Nymphalidae): How Many Butterflies Can Be Supported? Insects. 2018; 9(4):179. https://doi.org/10.3390/insects9040179
Chicago/Turabian StyleHill, Ryan I., Cassidi E. Rush, and John Mayberry. 2018. "Larval Food Limitation in a Speyeria Butterfly (Nymphalidae): How Many Butterflies Can Be Supported?" Insects 9, no. 4: 179. https://doi.org/10.3390/insects9040179
APA StyleHill, R. I., Rush, C. E., & Mayberry, J. (2018). Larval Food Limitation in a Speyeria Butterfly (Nymphalidae): How Many Butterflies Can Be Supported? Insects, 9(4), 179. https://doi.org/10.3390/insects9040179