
Thermal Stability Changes in Telomeric G-Quadruplex Structures Due to N6-Methyladenine Modification


Abstract
:1. Introduction
2. Results
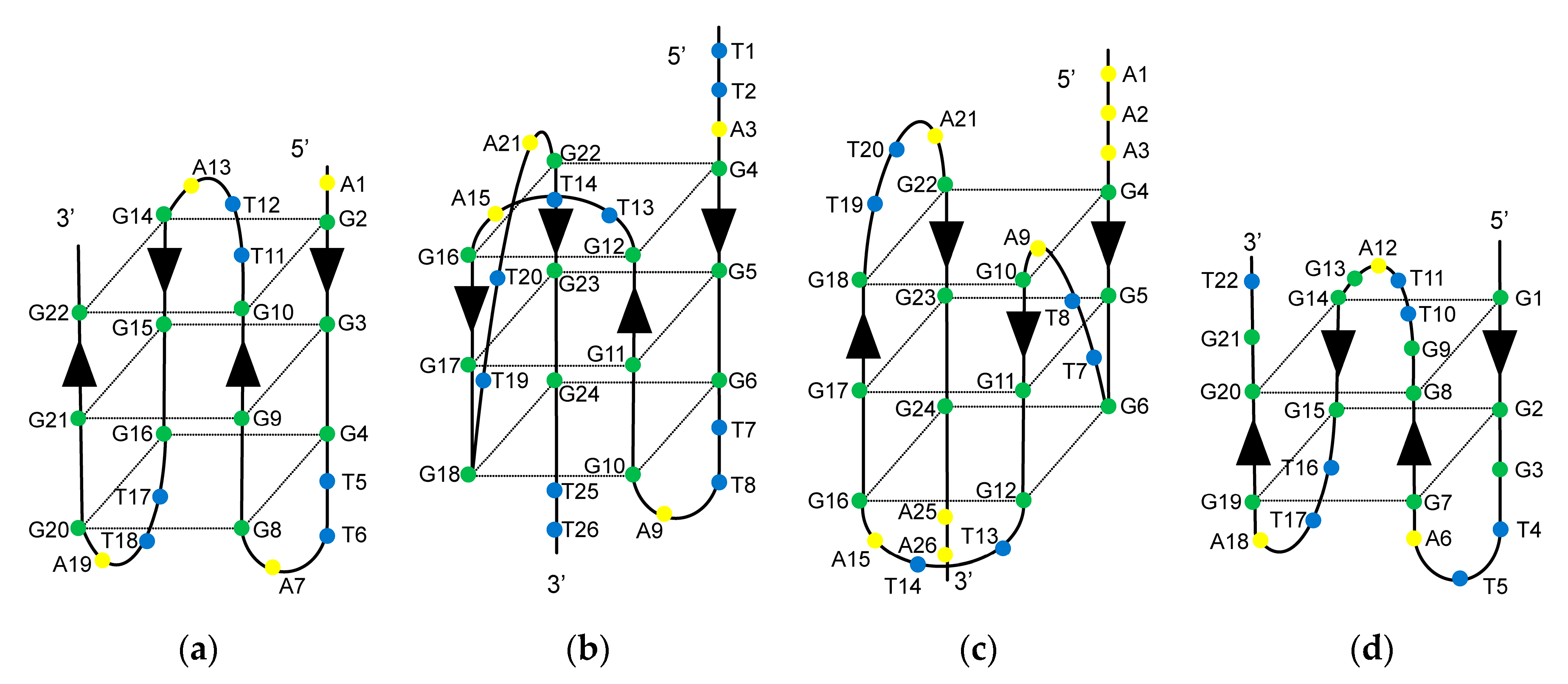
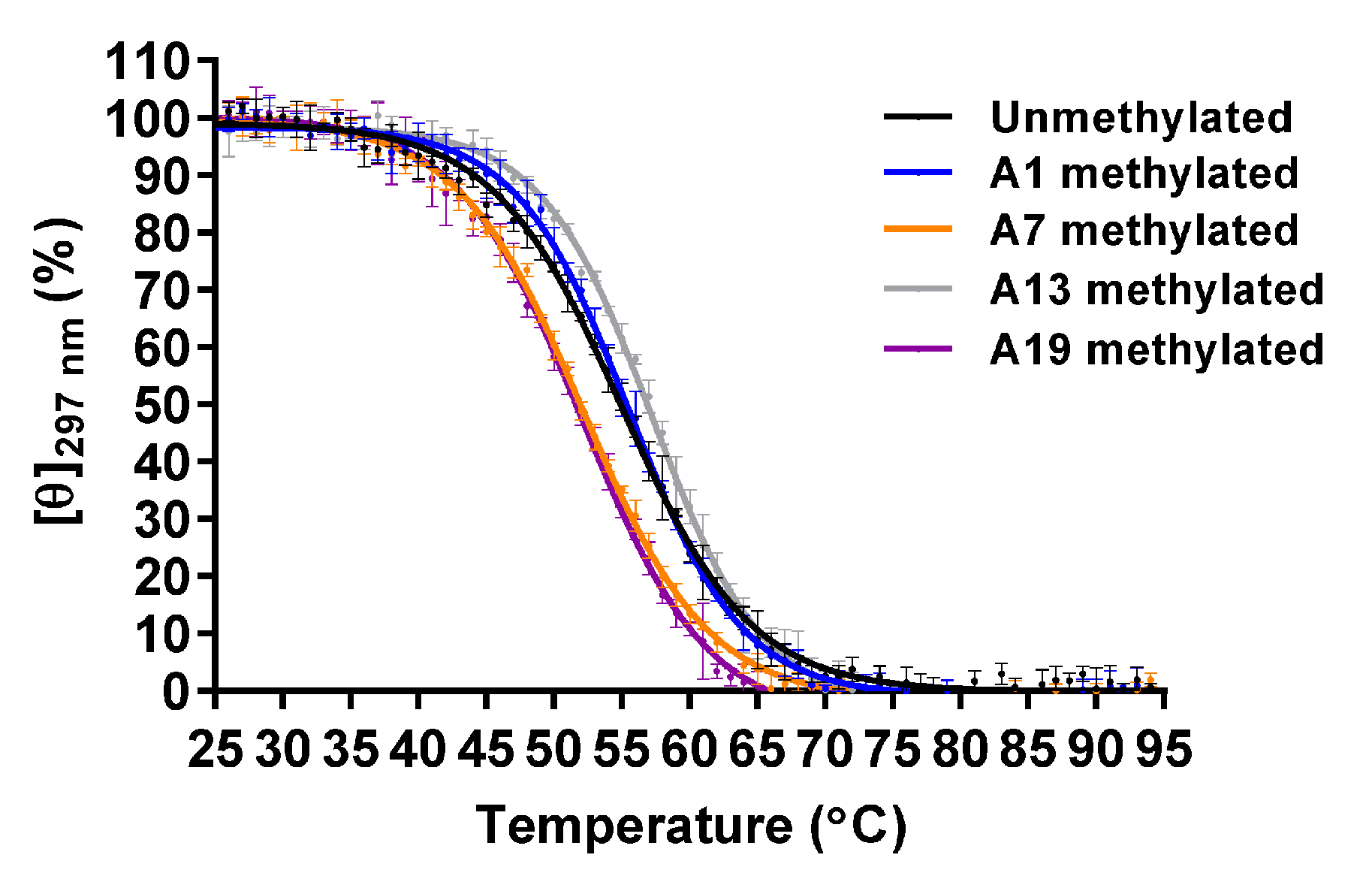
2.1. Effect of m6dA Modifications on the Thermal Stability of the Antiparallel Basket-Type G4 Structure That Formed in Na+
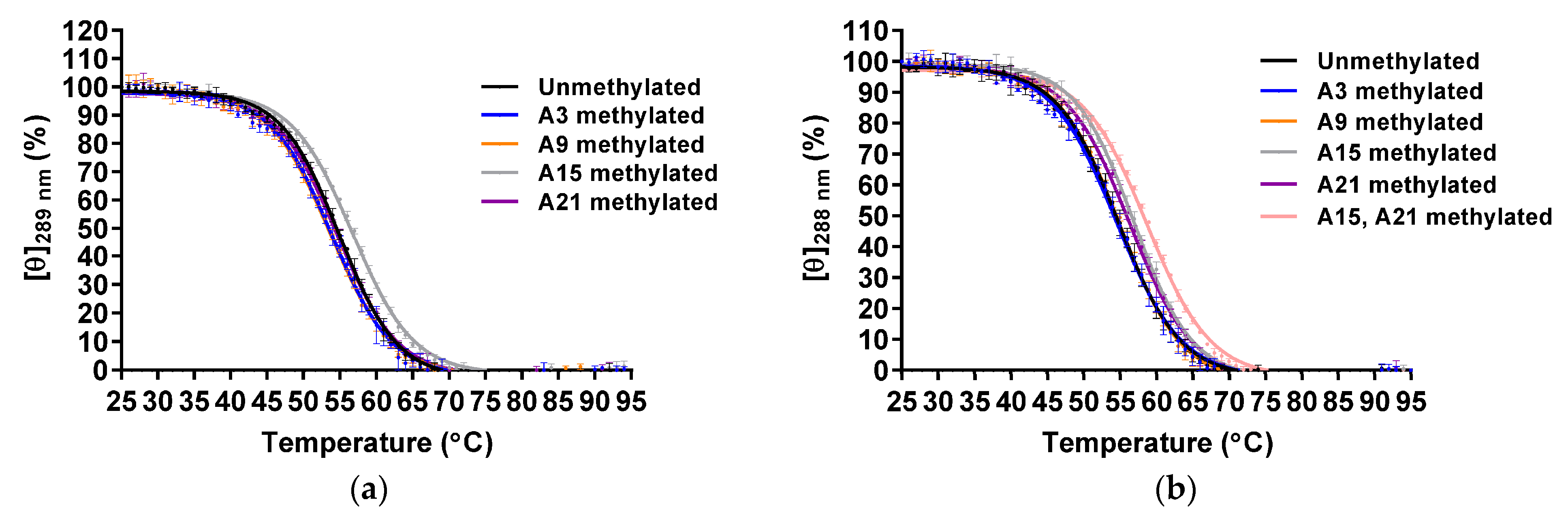
2.2. Effect of m6dA Modifications on the Thermal Stability of the Hybrid-Type G4 Structures Formed in K+
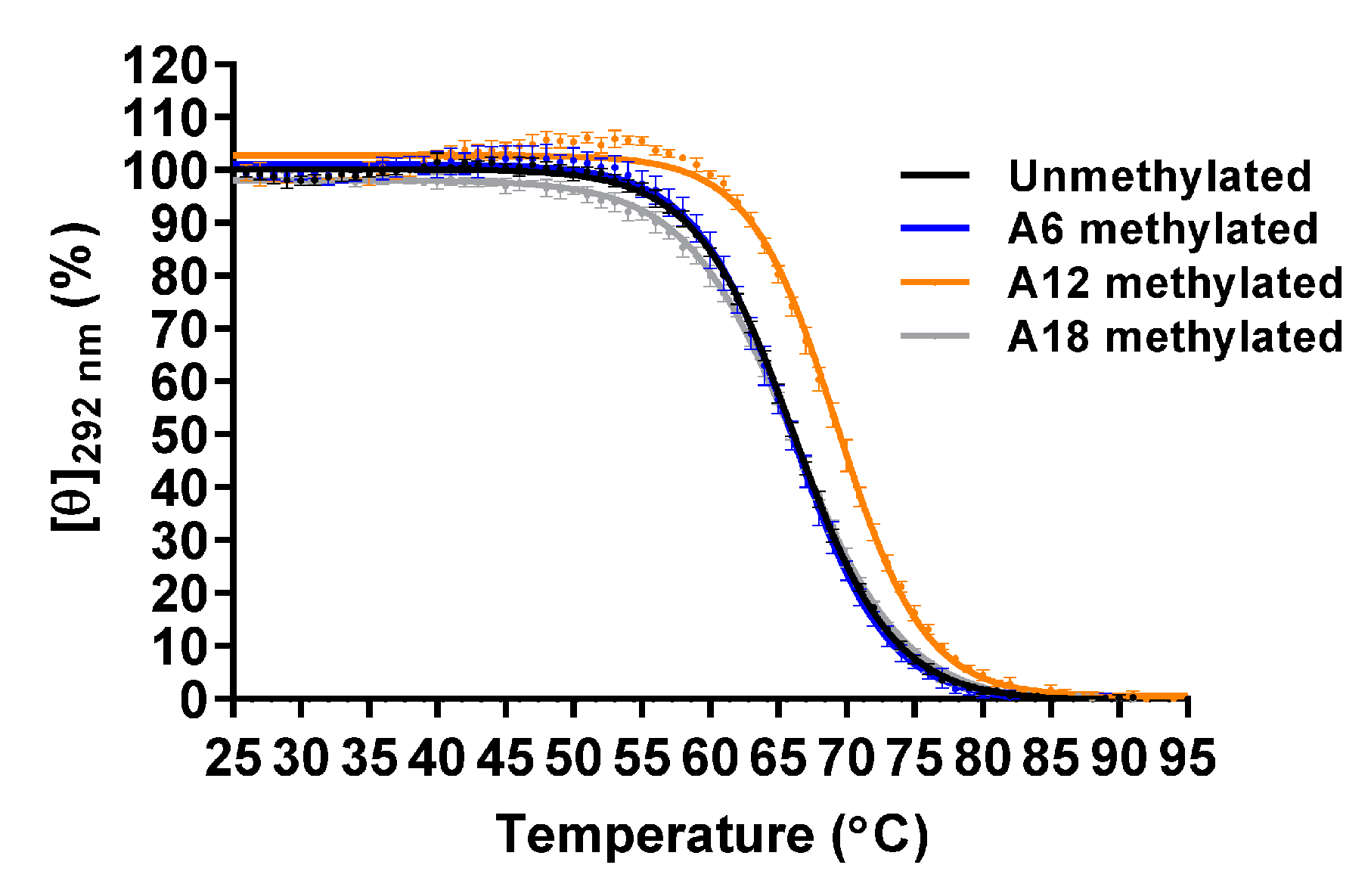
2.3. Effect of m6dA Modifications on the Thermal Stability of the Antiparallel Basket-Type G4 Structure Formed in K+
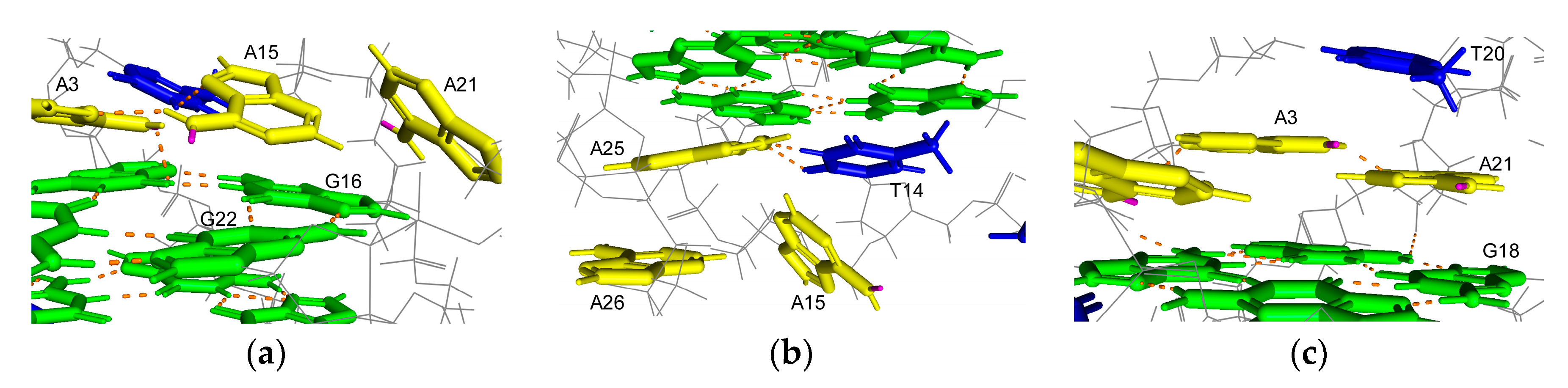
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. CD Analysis
4.3. PAGE Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human Telomere Biology: A Contributory and Interactive Factor in Aging, Disease Risks, and Protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef]
- Neidle, S.; Parkinson, G.N. The Structure of Telomeric DNA. Curr. Opin. Struct. Biol. 2003, 13, 275–283. [Google Scholar] [CrossRef]
- Yu, H.Q.; Miyoshi, D.; Sugimoto, N. Characterization of Structure and Stability of Long Telomeric DNA G-Quadruplexes. J. Am. Chem. Soc. 2006, 128, 15461–15468. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.; Mergny, J.L.; Salgado, G.F.; Queiroz, J.A.; Cruz, C. G-Quadruplex, Friend or Foe: The Role of the G-Quartet in Anticancer Strategies. Trends Mol. Med. 2020, 26, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, D.; Mirihana Arachchilage, G.; Basu, S. Metal Cations in G-Quadruplex Folding and Stability. Front. Chem. 2016, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, S.; Miyoshi, D.; Sugimoto, N. Effects of Molecular Crowding on the Structures, Interactions, and Functions of Nucleic Acids. Chem. Rev. 2014, 114, 2733–2758. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Hou, J.Q.; Xiang, H.D.; Yan, Y.Y.; Gu, Y.C.; Tan, J.H.; Li, D.; Gu, L.Q.; Ou, T.M.; Huang, Z.S. Stabilization of G-Quadruplex DNA by C-5-Methyl-Cytosine in Bcl-2 Promoter: Implications for Epigenetic Regulation. Biochem. Biophys. Res. Commun. 2013, 433, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, W.; Yoshioka, H.; Bay, D.H.; Iida, K.; Ikebukuro, K.; Nagasawa, K.; Karube, I. Detection of DNA Methylation of G-Quadruplex and I-Motif-Forming Sequences by Measuring the Initial Elongation Efficiency of Polymerase Chain Reaction. Anal. Chem. 2016, 88, 7101–7107. [Google Scholar] [CrossRef]
- Yoshida, W.; Terasaka, M.; Laddachote, S.; Karube, I. Stabilization of G-Quadruplex Structure on Vascular Endothelial Growth Factor Gene Promoter Depends on CpG Methylation Site and Cation Type. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1933–1937. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.J.; Stuffrein-Roberts, S.; Cree, S.L.; Gibb, A.; Miller, A.L.; Doudney, K.; Aitchison, A.; Eccles, M.R.; Joyce, P.R.; Filichev, V.V.; et al. G-Quadruplex Structures and CpG Methylation Cause Drop-out of the Maternal Allele in Polymerase Chain Reaction Amplification of the Imprinted Mest Gene Promoter. PLoS ONE 2014, 9, e113955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukakoshi, K.; Saito, S.; Yoshida, W.; Goto, S.; Ikebukuro, K. CpG Methylation Changes G-Quadruplex Structures Derived from Gene Promoters and Interaction with VEGF and SP1. Molecules 2018, 23, 944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, E.L.; Blanco, M.A.; Gu, L.; Sendinc, E.; Liu, J.; Aristizabal-Corrales, D.; Hsu, C.H.; Aravind, L.; He, C.; Shi, Y. DNA Methylation on N6-Adenine in C. Elegans. Cell 2015, 161, 868–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Luo, G.Z.; Chen, K.; Deng, X.; Yu, M.; Han, D.; Hao, Z.; Liu, J.; Lu, X.; Dore, L.C.; et al. N6-Methyldeoxyadenosine Marks Active Transcription Start Sites in Chlamydomonas. Cell 2015, 161, 879–892. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Huang, H.; Liu, D.; Cheng, Y.; Liu, X.; Zhang, W.; Yin, R.; Zhang, D.; Zhang, P.; Liu, J.; et al. N6-Methyladenine DNA Modification in Drosophila. Cell 2015, 161, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Koziol, M.J.; Bradshaw, C.R.; Allen, G.E.; Costa, A.S.H.; Frezza, C.; Gurdon, J.B. Identification of Methylated Deoxyadenosines in Vertebrates Reveals Diversity in DNA Modifications. Nat. Struct. Mol. Biol. 2016, 23, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Yao, B.; Cheng, Y.; Wang, Z.; Li, Y.; Chen, L.; Huang, L.; Zhang, W.; Chen, D.; Wu, H.; Tang, B.; et al. DNA N6-Methyladenine Is Dynamically Regulated in the Mouse Brain Following Environmental Stress. Nat. Commun. 2017, 8, 1122. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.P.; Wang, T.; Seetin, M.G.; Lai, Y.; Zhu, S.; Lin, K.; Liu, Y.; Byrum, S.D.; Mackintosh, S.G.; Zhong, M.; et al. DNA Methylation on N6-Adenine in Mammalian Embryonic Stem Cells. Nature 2016, 532, 329–333. [Google Scholar]
- Xiao, C.L.; Zhu, S.; He, M.; Chen, D.; Zhang, Q.; Chen, Y.; Yu, G.; Liu, J.; Xie, S.Q.; Luo, F.; et al. N6-Methyladenine DNA Modification in the Human Genome. Mol. Cell 2018, 71, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Wu, T.P.; Gimple, R.C.; Li, Z.; Prager, B.C.; Wu, Q.; Yu, Y.; Wang, P.; Wang, Y.; Gorkin, D.U.; et al. N6-Methyladenine DNA Modification in Glioblastoma. Cell 2018, 175, 1228–1243. [Google Scholar] [CrossRef] [Green Version]
- Engel, J.D.; Von Hippel, P.H. Effects of Methylation on the Stability of Nucleic Acid Conformations: Studies at the Monomer Level. Biochemistry 1974, 13, 4143–4158. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.D.; Von Hippel, P.H. Effects of Methylation on the Stability of Nucleic Acid Conformations. Studies at the Polymer Level. J. Biol. Chem. 1978, 253, 927–934. [Google Scholar] [CrossRef]
- Roost, C.; Lynch, S.R.; Batista, P.J.; Qu, K.; Chang, H.Y.; Kool, E.T. Structure and Thermodynamics of N6-Methyladenosine in RNA: A Spring-Loaded Base Modification. J. Am. Chem. Soc. 2015, 137, 2107–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laddachote, S.; Nagata, M.; Yoshida, W. Destabilisation of the c-Kit1 G-Quadruplex Structure by N6-Methyladenosine Modification. Biochem. Biophys. Res. Commun. 2020, 524, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Geng, Y.; Miao, H.; Shi, X.; You, Y.; Xu, N.; Zhou, B.; Zhu, G. G-Quadruplex Structures Formed by Human Telomeric DNA and C9orf72 Hexanucleotide Repeats. Biophys. Rev. 2019, 11, 389–393. [Google Scholar] [CrossRef]
- Wang, Y.; Patel, D.J. Solution Structure of the Human Telomeric Repeat D[AG3(T2AG3)3] G-Tetraplex. Structure 1993, 1, 263–282. [Google Scholar] [CrossRef]
- Phan, A.T.; Luu, K.N.; Patel, D.J. Different Loop Arrangements of Intramolecular Human Telomeric (3+1) G-Quadruplexes in K+ Solution. Nucleic Acids Res. 2006, 34, 5715–5719. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Carver, M.; Punchihewa, C.; Jones, R.A.; Yang, D. Structure of the Hybrid-2 Type Intramolecular Human Telomeric G-Quadruplex in K+ Solution: Insights into Structure Polymorphism of the Human Telomeric Sequence. Nucleic Acids Res. 2007, 35, 4927–4940. [Google Scholar] [CrossRef] [Green Version]
- Luu, K.N.; Phan, A.T.; Kuryavyi, V.; Lacroix, L.; Patel, D.J. Structure of the Human Telomere in K+ Solution: An Intramolecular (3 + 1) G-Quadruplex Scaffold. J. Am. Chem. Soc. 2006, 128, 9963–9970. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human Telomeric Sequence Forms a Hybrid-Type Intramolecular G-Quadruplex Structure with Mixed Parallel/Antiparallel Strands in Potassium Solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Punchihewa, C.; Ambrus, A.; Chen, D.; Jones, R.A.; Yang, D. Structure of the Intramolecular Human Telomeric G-Quadruplex in Potassium Solution: A Novel Adenine Triple Formation. Nucleic Acids Res. 2007, 35, 2440–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Noguchi, Y.; Sugiyama, H. The New Models of the Human Telomere d[AGGG(TTAGGG)3] in K+ Solution. Bioorg. Med. Chem. 2006, 14, 5584–5591. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.W.; Amrane, S.; Bouaziz, S.; Xu, W.; Mu, Y.; Patel, D.J.; Luu, K.N.; Phan, A.T. Structure of the Human Telomere in K+ Solution: A Stable Basket-Type G-Quadruplex with Only Two G-Tetrad Layers. J. Am. Chem. Soc. 2009, 131, 4301–4309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Dai, J.; Veliath, E.; Jones, R.A.; Yang, D. Structure of a Two-G-Tetrad Intramolecular G-Quadruplex Formed by a Variant Human Telomeric Sequence in K+ Solution: Insights into the Interconversion of Human Telomeric G-Quadruplex Structures. Nucleic Acids Res. 2010, 38, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.L.; Liu, H.S.; Xu, Y. Hybrid-Type and Two-Tetrad Antiparallel Telomere DNA G-Quadruplex Structures in Living Human Cells. Nucleic Acids Res. 2019, 47, 4940–4947. [Google Scholar] [CrossRef] [PubMed]
- Petraccone, L.; Trent, J.O.; Chaires, J.B. The Tail of the Telomere. J. Am. Chem. Soc. 2008, 130, 16530–16532. [Google Scholar] [CrossRef]
- Iida, K.; Nakamura, T.; Yoshida, W.; Tera, M.; Nakabayashi, K.; Hata, K.; Ikebukuro, K.; Nagasawa, K. Fluorescent-Ligand-Mediated Screening of G-Quadruplex Structures Using a DNA Microarray. Angew. Chem. Int. Ed. Engl. 2013, 52, 12052–12055. [Google Scholar] [CrossRef] [PubMed]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-Throughput Sequencing of DNA G-Quadruplex Structures in the Human Genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, W.; Saikyo, H.; Nakabayashi, K.; Yoshioka, H.; Bay, D.H.; Iida, K.; Kawai, T.; Hata, K.; Ikebukuro, K.; Nagasawa, K.; et al. Identification of G-Quadruplex Clusters by High-Throughput Sequencing of Whole-Genome Amplified Products with a G-Quadruplex Ligand. Sci. Rep. 2018, 8, 3116. [Google Scholar] [CrossRef]
- Zheng, K.W.; Zhang, J.Y.; He, Y.D.; Gong, J.Y.; Wen, C.J.; Chen, J.N.; Hao, Y.H.; Zhao, Y.; Tan, Z. Detection of Genomic G-Quadruplexes in Living Cells Using a Small Artificial Protein. Nucleic Acids Res. 2020, 48, 11706–11720. [Google Scholar] [CrossRef]
- Kwok, C.K.; Marsico, G.; Sahakyan, A.B.; Chambers, V.S.; Balasubramanian, S. rG4-Seq Reveals Widespread Formation of G-Quadruplex Structures in the Human Transcriptome. Nat. Methods 2016, 13, 841–844. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m6dA Site | Tm (°C) |
|---|---|
| None (unmethylated) | 54.9 ± 0.4 |
| A1 | 55.3 ± 0.4 |
| A7 | 51.9 ± 0.2 * |
| A13 | 56.9 ± 0.3 * |
| A19 | 51.6 ± 0.2 * |
| m6dA Site | Tm (°C) | |
|---|---|---|
| Form-2 G4 | Form-1 G4 | |
| None (unmethylated) | 54.5 ± 0.3 | 54.0 ± 0.5 |
| A3 | 53.5 ± 0.4 | 54.2 ± 0.5 |
| A9 | 53.3 ± 0.3 | 54.5 ± 0.1 |
| A15 | 56.4 ± 0.2 * | 56.6 ± 0.3 * |
| A21 | 54.2 ± 0.2 | 55.9 ± 0.3 * |
| A15 and A21 | n.d. | 58.2 ± 0.2 * |
| m6dA Site | Tm (°C) |
|---|---|
| None (unmethylated) | 66.1 ± 0.1 |
| A6 | 66.0 ± 0.5 |
| A12 | 69.5 ± 0.3 * |
| A18 | 66.0 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wada, R.; Yoshida, W. Thermal Stability Changes in Telomeric G-Quadruplex Structures Due to N6-Methyladenine Modification. Epigenomes 2021, 5, 5. https://doi.org/10.3390/epigenomes5010005
Wada R, Yoshida W. Thermal Stability Changes in Telomeric G-Quadruplex Structures Due to N6-Methyladenine Modification. Epigenomes. 2021; 5(1):5. https://doi.org/10.3390/epigenomes5010005
Chicago/Turabian StyleWada, Ryohei, and Wataru Yoshida. 2021. "Thermal Stability Changes in Telomeric G-Quadruplex Structures Due to N6-Methyladenine Modification" Epigenomes 5, no. 1: 5. https://doi.org/10.3390/epigenomes5010005
APA StyleWada, R., & Yoshida, W. (2021). Thermal Stability Changes in Telomeric G-Quadruplex Structures Due to N6-Methyladenine Modification. Epigenomes, 5(1), 5. https://doi.org/10.3390/epigenomes5010005