Enlisting the Ixodes scapularis Embryonic ISE6 Cell Line to Investigate the Neuronal Basis of Tick—Pathogen Interactions

 ,
,  , , ,
, , ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Neuropeptidome of ISE6 Cells
2.2. Structure of the Gene-Encoding Ixodes TRP
2.3. Expression of ISE6 TRP in Response to A. phagocytophilum Infection
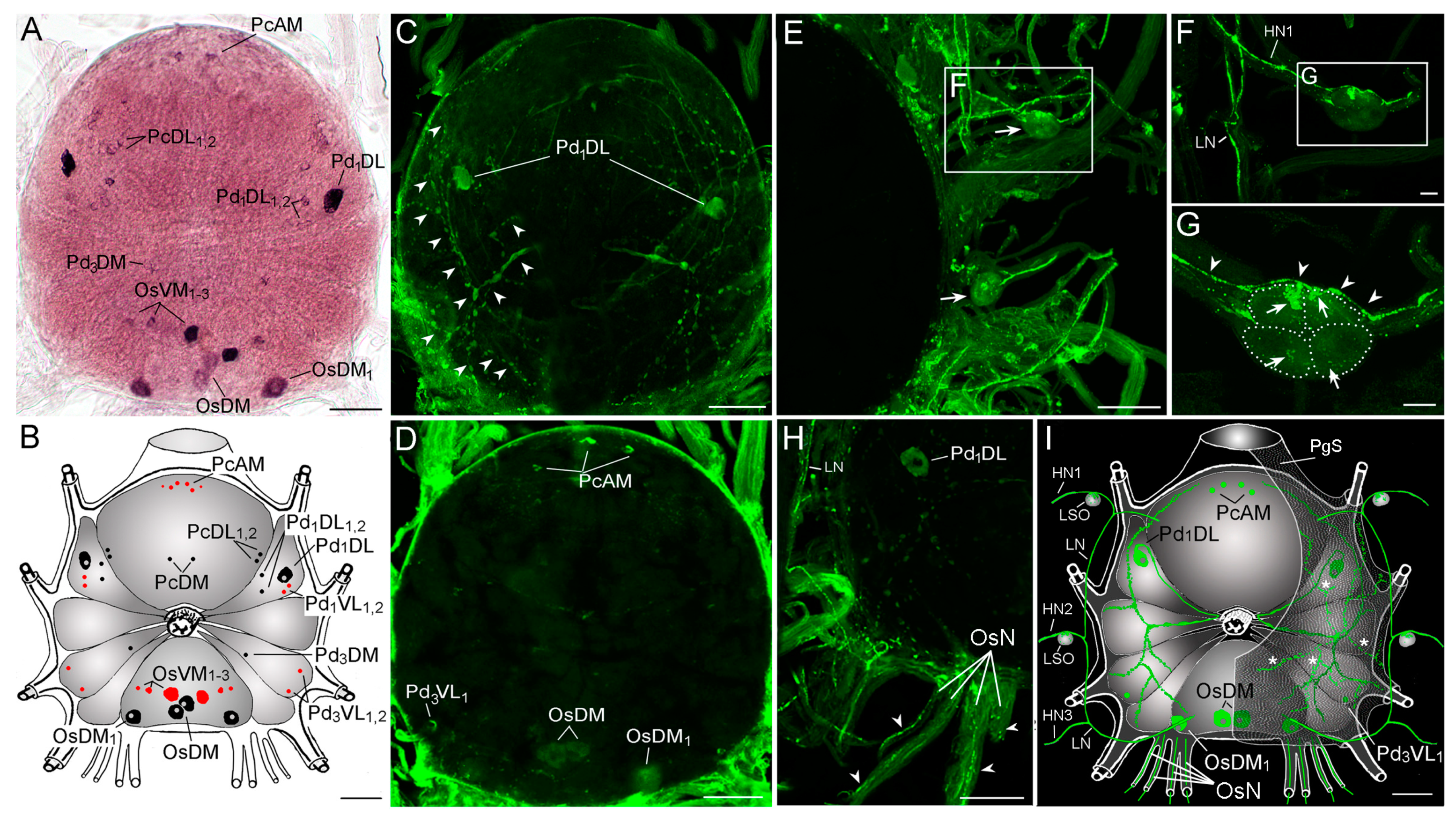
2.4. Expression of TRP in I. scapularis Synganglion
3. Discussion
4. Materials and Methods
4.1. In Silico Identification of Neuropeptides Genes in ISE6 Databases
4.2. Culture of ISE6 Cells
4.3. Neuropeptides Quantitative Real-Time PCR in ISE6 Cells
4.4. Gene Cloning and Sequence Analyses
4.5. Pathogen Infection of ISE6 Cells
4.6. Immunocytochemistry of TRP in Uninfected and A. phagocytophilum-Infected ISE6 Cells
4.7. In Situ Hybridization
4.8. Wholemount Immunohistochemistry of Ixodes Synganglion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamer, S.A.; Tsao, J.I.; Walker, E.D.; Hickling, G.J. Invasion of the lyme disease vector Ixodes scapularis: Implications for Borrelia burgdorferi endemicity. Ecohealth 2010, 7, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and Its Transmitted Pathogens in Urban and Peri-Urban Areas in Europe: New Hazards and Relevance for Public Health. Front. Public Health 2014, 2. [Google Scholar] [CrossRef]
- Kocan, K.M.; De la Fuente, J.; Cabezas-Cruz, A. The genus Anaplasma: New challenges after reclassification. Rev. Off. Int. Epizoot. 2015, 34, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Kocan, K.M.; De la Fuente, J.; Blouin, E.F.; Garcia-Garcia, J.C. Anaplasma marginale (Rickettsiales: Anaplasmataceae): Recent advances in defining host-pathogen adaptations of a tick-borne rickettsia. Parasitology 2004, 129, S285–S300. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldridge, G.D.; Burkhardt, N.Y.; Labruna, M.B.; Pacheco, R.C.; Paddock, C.D.; Williamson, P.C.; Billingsley, P.M.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Wide dispersal and possible multiple origins of low-copy-number plasmids in rickettsia species associated with blood-feeding arthropods. Appl. Environ. Microbiol. 2010, 76, 1718–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munderloh, U.G.; Blouin, E.F.; Kocan, K.M.; Ge, N.L.; Edwards, W.L.; Kurtti, T.J. Establishment of the Tick (Acari: Ixodidae)-Borne Cattle Pathogen Anaplasma marginale (Rickettsiales: Anaplasmataceae) in Tick Cell Culture. J. Med. Entomol. 1996, 33, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Munderloh, U.G.; Silverman, D.J.; MacNamara, K.C.; Ahlstrand, G.G.; Chatterjee, M.; Winslow, G.M. Ixodes ovatus Ehrlichia exhibits unique ultrastructural characteristics in mammalian endothelial and tick-derived cells. Ann. N. Y. Acad. Sci. 2009, 1166, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Munderloh, U.G.; Yabsley, M.J.; Murphy, S.M.; Luttrell, M.P.; Howerth, E.W. Isolation and establishment of the raccoon Ehrlichia-like agent in tick cell culture. Vector Borne Zoonotic Dis. 2007, 7, 418–425. [Google Scholar] [CrossRef]
- Tate, C.M.; Howerth, E.W.; Mead, D.G.; Dugan, V.G.; Luttrell, M.P.; Sahora, A.I.; Munderloh, U.G.; Davidson, W.R.; Yabsley, M.J. Anaplasma odocoilei sp. nov. (family Anaplasmataceae) from white-tailed deer (Odocoileus virginianus). Ticks Tick Borne Dis. 2013, 4, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Pornwiroon, W.; Pourciau, S.S.; Foil, L.D.; Macaluso, K.R. Rickettsia felis from Cat Fleas: Isolation and Culture in a Tick-Derived Cell Line. Appl. Environ. Microbiol. 2006, 72, 5589–5595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a Spotted Fever Group Rickettsia, from Ticks (Ixodes ricinus) Collected in a European City Park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munderloh, U.G.; Tate, C.M.; Lynch, M.J.; Howerth, E.W.; Kurtti, T.J.; Davidson, W.R. Isolation of an Anaplasma sp. Organism from White-Tailed Deer by Tick Cell Culture. J. Clin. Microbiol. 2003, 41, 4328–4335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obonyo, M.; Munderloh, U.G.; Fingerle, V.; Wilske, B.; Kurtti, T.J. Borrelia burgdorferi in Tick Cell Culture Modulates Expression of Outer Surface Proteins A and C in Response to Temperature. J. Clin. Microbiol. 1999, 37, 2137–2141. [Google Scholar] [CrossRef] [Green Version]
- Garcia, S.; Billecocq, A.; Crance, J.-M.; Munderloh, U.; Garin, D.; Bouloy, M. Nairovirus RNA Sequences Expressed by a Semliki Forest Virus Replicon Induce RNA Interference in Tick Cells. J. Virol. 2005, 79, 8942–8947. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, J.M.; Perera, R.; Roumani, A.M.; Hedrick, V.E.; Inerowicz, H.D.; Hill, C.A.; Kuhn, R.J. Changes in the Proteome of Langat-Infected Ixodes scapularis ISE6 Cells: Metabolic Pathways Associated with Flavivirus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004180. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Liu, Y.; Wang, M.; Chen, C.; Kurtti, T.J. Establishment, maintenance and description of cell lines from the tick Ixodes scapularis. J. Parasitol. 1994, 80, 533–543. [Google Scholar] [CrossRef]
- Oliver, J.D.; Chávez, A.S.O.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. An Ixodes scapularis cell line with a predominantly neuron-like phenotype. Exp. Appl. Acarol. 2015, 66, 427–442. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, P.; Mansfield, K.L.; Manzano-Román, R.; Cook, C.; Ayllón, N.; Villar, M.; Johnson, N.; Fooks, A.R.; De la Fuente, J. Tissue-Specific Signatures in the Transcriptional Response to Anaplasma phagocytophilum Infection of Ixodes scapularis and Ixodes ricinus Tick Cell Lines. Front. Cell Infect. Microbiol. 2016, 6, 20. [Google Scholar] [CrossRef]
- Artigas-Jerónimo, S.; Alberdi, P.; Rayo, M.V.; Cabezas-Cruz, A.; Prados, P.J.E.; Mateos-Hernández, L.; De la Fuente, J. Anaplasma phagocytophilum modifies tick cell microRNA expression and upregulates isc-mir-79 to facilitate infection by targeting the Roundabout protein 2 pathway. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Espinosa, P.; Alberdi, P.; De la Fuente, J. Tick–Pathogen Interactions: The Metabolic Perspective. Trends Parasitol. 2019, 35, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.K.; Wang, X.; Brown, L.J.; Chávez, A.S.O.; Reif, K.E.; Smith, A.A.; Scott, A.J.; McClure, E.E.; Boradia, V.M.; Hammond, H.L.; et al. Infection-derived lipids elicit an immune deficiency circuit in arthropods. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente, J.; Villar, M.; Cabezas-Cruz, A.; Estrada-Peña, A.; Ayllón, N.; Alberdi, P. Tick–Host–Pathogen Interactions: Conflict and Cooperation. PLoS Pathog. 2016, 12, e1005488. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Kikuchi, T.; Brewer, H.M.; Norbeck, A.D.; Rikihisa, Y. Global Proteomic Analysis of Two Tick-Borne Emerging Zoonotic Agents: Anaplasma Phagocytophilum and Ehrlichia Chaffeensis. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Ayllón, N.; Villar, M.; Galindo, R.C.; Kocan, K.M.; Šíma, R.; López, J.A.; Vázquez, J.; Alberdi, P.; Cabezas-Cruz, A.; Kopáček, P.; et al. Systems Biology of Tissue-Specific Response to Anaplasma phagocytophilum Reveals Differentiated Apoptosis in the Tick Vector Ixodes scapularis. PLoS Genet. 2015, 11, e1005120. [Google Scholar] [CrossRef] [Green Version]
- Cabezas-Cruz, A.; Alberdi, P.; Valdés, J.J.; Villar, M.; De la Fuente, J. Anaplasma phagocytophilum Infection Subverts Carbohydrate Metabolic Pathways in the Tick Vector, Ixodes scapularis. Front. Cell Infect. Microbiol. 2017, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Cabezas-Cruz, A.; Espinosa, P.J.; Obregón, D.A.; Alberdi, P.; De la Fuente, J. Ixodes scapularis Tick Cells Control Anaplasma phagocytophilum Infection by Increasing the Synthesis of Phosphoenolpyruvate from Tyrosine. Front. Cell Infect. Microbiol. 2017, 7, 375. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, S.H.G.; Garcia-Garcia, J.C.; Dumler, J.S. Bioinformatic and mass spectrometry identification of Anaplasma phagocytophilum proteins translocated into host cell nuclei. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Sinclair, S.H.; Rennoll-Bankert, K.E.; Dumler, J.S. Effector bottleneck: Microbial reprogramming of parasitized host cell transcription by epigenetic remodeling of chromatin structure. Front. Genet. 2014, 5. [Google Scholar] [CrossRef]
- Villar, M.; Ayllón, N.; Alberdi, P.; Moreno, A.; Moreno, M.; Tobes, R.; Mateos-Hernández, L.; Weisheit, S.; Bell-Sakyi, L.; De la Fuente, J. Integrated Metabolomics, Transcriptomics and Proteomics Identifies Metabolic Pathways Affected by Anaplasma phagocytophilum Infection in Tick Cells. Mol. Cell Proteom. 2015, 14, 3154–3172. [Google Scholar] [CrossRef] [Green Version]
- Cabezas-Cruz, A.; Alberdi, P.; Valdes, J.J.; Villar, M.; De la Fuente, J. Remodeling of tick cytoskeleton in response to infection with Anaplasma phagocytophilum. Front. Biosci. 2017, 22, 1830–1844. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G. Pathogens Manipulating Tick Behavior—Through a Glass, Darkly. Pathogens 2020, 9, 664. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E. Tick ecology: Processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors. Parasitology 2004, 129, S37–S65. [Google Scholar] [CrossRef] [PubMed]
- Neelakanta, G.; Sultana, H.; Fish, D.; Anderson, J.F.; Fikrig, E. Anaplasma phagocytophilum induces Ixodes scapularis ticks to express an antifreeze glycoprotein gene that enhances their survival in the cold. J. Clin. Investig. 2010, 120, 3179–3190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busby, A.T.; Ayllón, N.; Kocan, K.M.; Blouin, E.F.; De la Fuente, G.; Galindo, R.C.; Villar, M.; De la Fuente, J. Expression of heat shock proteins and subolesin affects stress responses, Anaplasma phagocytophilum infection and questing behaviour in the tick, Ixodes scapularis. Med. Vet. Entomol. 2012, 26, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Lynn, G.E.; Oliver, J.D.; Nelson, C.M.; Felsheim, R.F.; Kurtti, T.J.; Munderloh, U.G. Tissue Distribution of the Ehrlichia muris-Like Agent in a Tick Vector. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Šimo, L.; Daniel, S.E.; Park, Y.; Žitňan, D. The Nervous and Sensory Systems: Structure, Function, Proteomics and Genomics. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 1, pp. 309–367. [Google Scholar]
- Šimo, L.; Slovák, M.; Park, Y.; Zitnan, D. Identification of a complex peptidergic neuroendocrine network in the hard tick, Rhipicephalus appendiculatus. Cell Tissue Res. 2009, 335, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Hewes, R.S.; Taghert, P.H. Neuropeptides and Neuropeptide Receptors in the Drosophila melanogaster Genome. Genome Res. 2001, 11, 1126–1142. [Google Scholar] [CrossRef] [Green Version]
- Caers, J.; Verlinden, H.; Zels, S.; Vandersmissen, H.P.; Vuerinckx, K.; Schoofs, L. More than two decades of research on insect neuropeptide GPCRs: An overview. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Yeoh, J.G.C.; Pandit, A.A.; Zandawala, M.; Nässel, D.R.; Davies, S.-A.; Dow, J.A.T. DINeR: Database for Insect Neuropeptide Research. Insect Biochem. Mol. Biol. 2017, 86, 9–19. [Google Scholar] [CrossRef]
- Schoofs, L.; De Loof, A.; Van Hiel, M.B. Neuropeptides as Regulators of Behavior in Insects. Annu. Rev. Entomol. 2017, 62, 35–52. [Google Scholar] [CrossRef]
- Gulia-Nuss, M.; Nuss, A.B.; Meyer, J.M.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; De la Fuente, J.; Ribeiro, J.M.; Megy, K.; et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupert, S.; Russell, W.K.; Predel, R.; Russell, D.H.; Strey, O.F.; Teel, P.D.; Nachman, R.J. The neuropeptidomics of Ixodes scapularis synganglion. J. Proteom. 2009, 72, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Žitňan, D.; Park, Y. Two novel neuropeptides in innervation of the salivary glands of the black-legged tick, Ixodes scapularis: Myoinhibitory peptide and SIFamide. J. Comp. Neurol. 2009, 517, 551–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancová, M.; Bílý, T.; Nebesářová, J.; Grubhoffer, L.; Bonnet, S.; Park, Y.; Šimo, L. Ultrastructural mapping of salivary gland innervation in the tick Ixodes ricinus. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Koči, J.; Park, Y. Receptors for the neuropeptides, myoinhibitory peptide and SIFamide, in control of the salivary glands of the blacklegged tick Ixodes scapularis. Insect. Biochem. Mol. Biol. 2013, 43, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Nässel, D.R.; Zandawala, M.; Kawada, T.; Satake, H. Tachykinins: Neuropeptides That Are Ancient, Diverse, Widespread and Functionally Pleiotropic. Front. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Jiang, H.; Kim, D.; Dobesh, S.; Evans, J.D.; Nachman, R.J.; Kaczmarek, K.; Zabrocki, J.; Park, Y. Ligand selectivity in tachykinin and natalisin neuropeptidergic systems of the honey bee parasitic mite Varroa destructor. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Lkhagva, A.; Daubnerová, I.; Chae, H.-S.; Šimo, L.; Jung, S.-H.; Yoon, Y.-K.; Lee, N.-R.; Seong, J.Y.; Žitňan, D.; et al. Natalisin, a tachykinin-like signaling system, regulates sexual activity and fecundity in insects. Proc. Natl. Acad. Sci. USA 2013, 110, E3526–E3534. [Google Scholar] [CrossRef] [Green Version]
- Coast, G.M.; Schooley, D.A. Toward a consensus nomenclature for insect neuropeptides and peptide hormones. Peptides 2011, 32, 620–631. [Google Scholar] [CrossRef]
- Miller, J.R.; Koren, S.; Dilley, K.A.; Harkins, D.M.; Stockwell, T.B.; Shabman, R.S.; Sutton, G.G. A draft genome sequence for the Ixodes scapularis cell line, ISE6. F1000Res. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Almazán, C.; Šimo, L.; Fourniol, L.; Rakotobe, S.; Borneres, J.; Cote, M.; Peltier, S.; Mayé, J.; Versillé, N.; Richardson, J.; et al. Multiple Antigenic Peptide-Based Vaccines Targeting Ixodes ricinus Neuropeptides Induce a Specific Antibody Response but Do Not Impact Tick Infestation. Pathogens 2020, 9, 900. [Google Scholar] [CrossRef]
- Brock, C.M.; Temeyer, K.B.; Tidwell, J.; Yang, Y.; Blandon, M.A.; Carreón-Camacho, D.; Longnecker, M.T.; Almazán, C.; Pérez de León, A.A.; Pietrantonio, P.V. The leucokinin-like peptide receptor from the cattle fever tick, Rhipicephalus microplus, is localized in the midgut periphery and receptor silencing with validated double-stranded RNAs causes a reproductive fitness cost. Int. J. Parasitol. 2019, 49, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Opdebeeck, J.P.; Wong, J.Y.; Jackson, L.A.; Dobson, C. Vaccines to protect Hereford cattle against the cattle tick, Boophilus microplus. Immunology 1988, 63, 363–367. [Google Scholar] [PubMed]
- Christie, A.E. Neuropeptide discovery in Ixodoidea: An in silico investigation using publicly accessible expressed sequence tags. Gen. Comp. Endocrinol. 2008, 157, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Park, Y. Endocrine regulation of insect diuresis in the early postgenomic era1. Can. J. Zool. 2012. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Šimo, L.; Park, Y. Molecular characterization of neuropeptide elevenin and two elevenin receptors, IsElevR1 and IsElevR2, from the blacklegged tick, Ixodes scapularis. Insect. Biochem. Mol. Biol. 2018, 101, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Koči, J.; Šimo, L.; Park, Y. Validation of Internal Reference Genes for Real-Time Quantitative Polymerase Chain Reaction Studies in the Tick, Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2013, 50, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Silaghi, C.; Nieder, M.; Sauter-Louis, C.; Knubben-Schweizer, G.; Pfister, K.; Pfeffer, M. Epidemiology, genetic variants and clinical course of natural infections with Anaplasma phagocytophilum in a dairy cattle herd. Parasites Vectors 2018, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuen, S.; Van De Pol, I.; Bergström, K.; Schouls, L.M. Identification of Anaplasma phagocytophila (Formerly Ehrlichia phagocytophila) Variants in Blood from Sheep in Norway. J. Clin. Microbiol. 2002, 40, 3192–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanovich, K.M.; Bakken, J.S.; Madigan, J.E.; Rosenfeld, M.A.; Wormser, G.P.; Dumler, J.S. Antigenic Diversity of Granulocytic Ehrlichia Isolates from Humans in Wisconsin and New York and a Horse in California. J. Infect. Dis. 1997, 176, 1029–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateos-Hernandéz, L.; Defaye, B.; Vancová, M.; Hajdusek, O.; Sima, R.; Park, Y.; Attoui, H.; Šimo, L. Cholinergic axons regulate type I acini in salivary glands of Ixodes ricinus and Ixodes scapularis ticks. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Neuropeptide Name | Computed Annotation | Scaffold | mRNA (SRA) |
|---|---|---|---|
| Adipokinetic hormone/corazonin-related peptide, (ACP) | XR_003917229 | PKSA02006111.1 | ✓ |
| Achatin-like (GFGE) | XM_029988349 | PKSA02000030.1 | ✓ |
| Allatostatin CC | ISCI001408 | PKSA02005862.1 | ✓ |
| Allatotropin | ISCI017791 | PKSA02000866.1 | ✓ |
| Arginine-vasopressin-like peptide (Inotocin) | XM_029985633 1 | PKSA02000317.1; PKSA02014421.1 | ✓ |
| Bursicon alpha | ISCI004617 | PKSA02001242.1 | ✓ |
| Bursicon beta | ISCI004618 | PKSA02001242.1 | ✓ |
| Calcitonin-like diuretic hormone 34a | ISCI020490 1 | PKSA02012946.1; PKSA02003019.1 | ✓ |
| Calcitonin-like diuretic hormone 34b | ISCI009341 | PKSA02005071.1 | ✓ |
| CCHamide-1 | ISCI013057 | PKSA02001506.1 | ✓ |
| Corticotropin-releasing factor-related diuretic hormone | ISCI007845 1 | PKSA02003125.1; PKSA02006996.1 | ✓ |
| Corazonin | ISCI014429 | PKSA02006111.1 | ✓ |
| Crustacean cardioactive peptide | ISCI010619 | PKSA02006111.1 | ✓ |
| Crustacean hyperglycaemic hormone/related ion transport peptide (CHH/ITP) isoform a | ISCI023228 1 | PKSA02003886.1; PKSA02006116.1 | ✓ |
| Crustacean hyperglycaemic hormone/related ion transport peptide (CHH/ITP) isoform b | XM_029989718 1 | PKSA02003886.1; PKSA02006116.1 | ✓ |
| Eclosion hormone | ISCI001941 | PKSA02005732.1 | ✓ |
| EFLamide | ISCW014582 1 | PKSA02010407.1; PKSA02009519.1 | ✓ |
| Elevenin | AXL48134.1 1,2 | PKSA02008257.1; PKSA02016953.1 PKSA02007065.1; PKSA02009048.1 | ✓ |
| FGLa-related allatostatin (Allatostatin A) | ISCI022939 1 | PKSA02013181.1; PKSA02004535.1 | ✓ |
| Glycoprotein A2 | XM_029991759 1 | PKSA02013868.1; PKSA02004257.1 | ✓ |
| Glycoprotein B5 | ISCI010926 1 | PKSA02004257.1; PKSA02012429.1 | ✓ |
| Insulin-related peptide 1 | ISCI002549 1 | PKSA02002782.1; PKSA02000328.1 | ✓ |
| Insulin-related peptide 2 | ISCW020331 | PKSA02008623.1; PKSA02013879.1 | ✓ |
| PISCF-related allatostatin (Allatostatin C) | ISCI001803 | PKSA02005862.1 | ✓ |
| Myoinhibitory peptide (Allatostatin B) | ISCI017595 1 | PKSA02004180.1; PKSA02003484.1 | ✓ |
| Kinin | ISCI024200 | PKSA02005591.1 | ✓ |
| Natalisin | ISCI021632 | PKSA02002188.1 | ND |
| Neuroparsin | XM_0299932151 | PKSA02004554.1 | ✓ |
| Orcokinin | ISCI010518 64.1 | PKSA02003264.1 | ✓ |
| Proctolin | ISCI005701 1,2 |
PKSA02003526.1; PKSA02018840.1 PKSA02018839.1 | ✓ |
| Prothoracicotropic hormone-like (Trunk) | ISCI001809 | PKSA02005517.1 | ✓ |
| Pyrokinin | ISCI019582 | PKSA02005732.1 | ✓ |
| RYamide | ISCI005825 | PKSA02000030.1 | ✓ |
| SIFamide | ISCI022950 | PKSA02005457.1 | ✓ |
| Short neuropeptide F | ISCI007409 | PKSA02002025.1 | ✓ |
| Sulfakinin | XM_029979447 | PKSA02001743.1 | ✓ |
| Tachykinin related peptide (TRP) | ISCI008383 * | PKSA02005591.1 | ✓ |
| Trissin | ISCI011258 | PKSA02005519.1 | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateos-Hernández, L.; Pipová, N.; Allain, E.; Henry, C.; Rouxel, C.; Lagrée, A.-C.; Haddad, N.; Boulouis, H.-J.; Valdés, J.J.; Alberdi, P.; et al. Enlisting the Ixodes scapularis Embryonic ISE6 Cell Line to Investigate the Neuronal Basis of Tick—Pathogen Interactions. Pathogens 2021, 10, 70. https://doi.org/10.3390/pathogens10010070
Mateos-Hernández L, Pipová N, Allain E, Henry C, Rouxel C, Lagrée A-C, Haddad N, Boulouis H-J, Valdés JJ, Alberdi P, et al. Enlisting the Ixodes scapularis Embryonic ISE6 Cell Line to Investigate the Neuronal Basis of Tick—Pathogen Interactions. Pathogens. 2021; 10(1):70. https://doi.org/10.3390/pathogens10010070
Chicago/Turabian StyleMateos-Hernández, Lourdes, Natália Pipová, Eléonore Allain, Céline Henry, Clotilde Rouxel, Anne-Claire Lagrée, Nadia Haddad, Henri-Jean Boulouis, James J. Valdés, Pilar Alberdi, and et al. 2021. "Enlisting the Ixodes scapularis Embryonic ISE6 Cell Line to Investigate the Neuronal Basis of Tick—Pathogen Interactions" Pathogens 10, no. 1: 70. https://doi.org/10.3390/pathogens10010070
APA StyleMateos-Hernández, L., Pipová, N., Allain, E., Henry, C., Rouxel, C., Lagrée, A. -C., Haddad, N., Boulouis, H. -J., Valdés, J. J., Alberdi, P., de la Fuente, J., Cabezas-Cruz, A., & Šimo, L. (2021). Enlisting the Ixodes scapularis Embryonic ISE6 Cell Line to Investigate the Neuronal Basis of Tick—Pathogen Interactions. Pathogens, 10(1), 70. https://doi.org/10.3390/pathogens10010070