Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections

 ,
,  and
and
Abstract
:1. Introduction
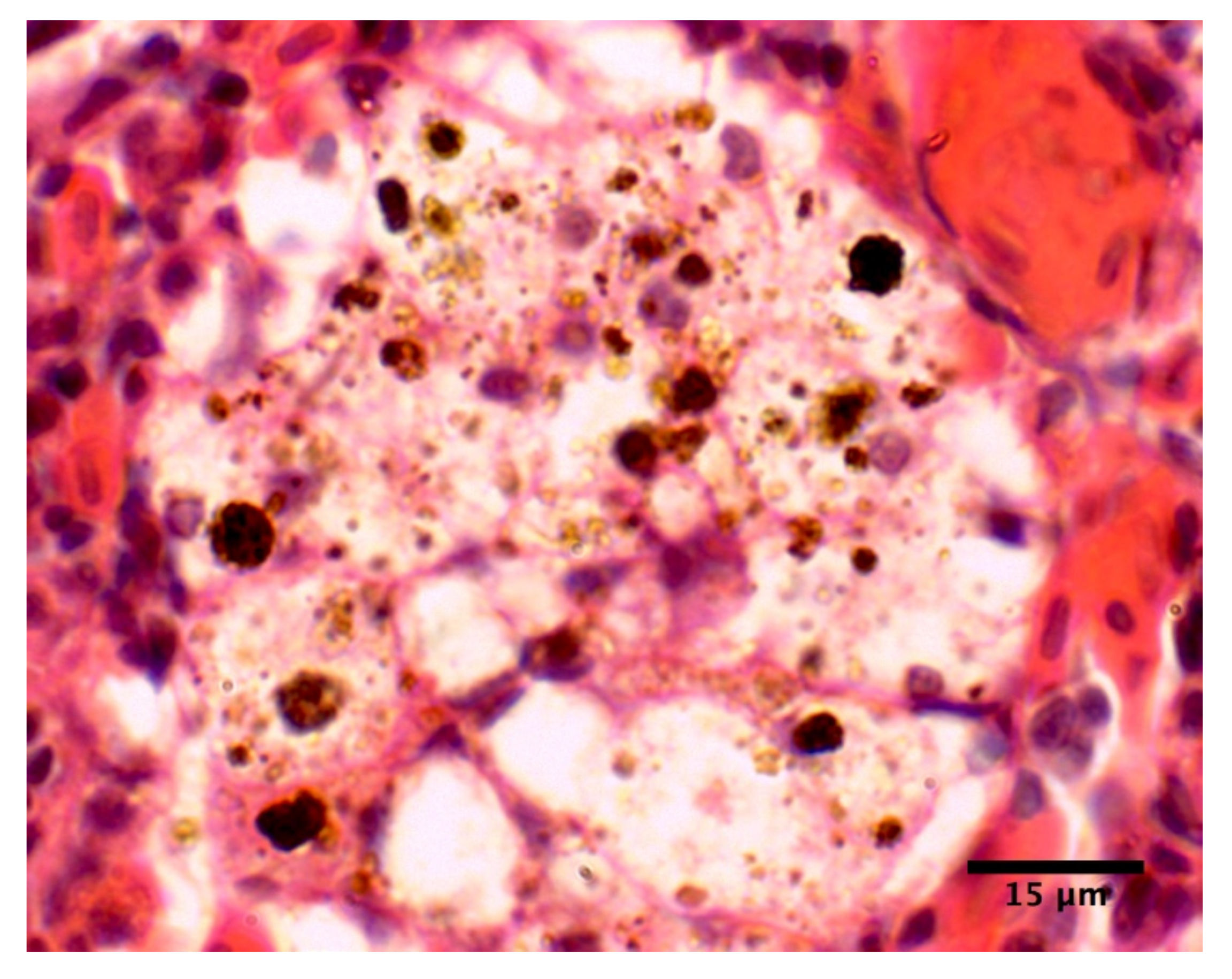
2. Results
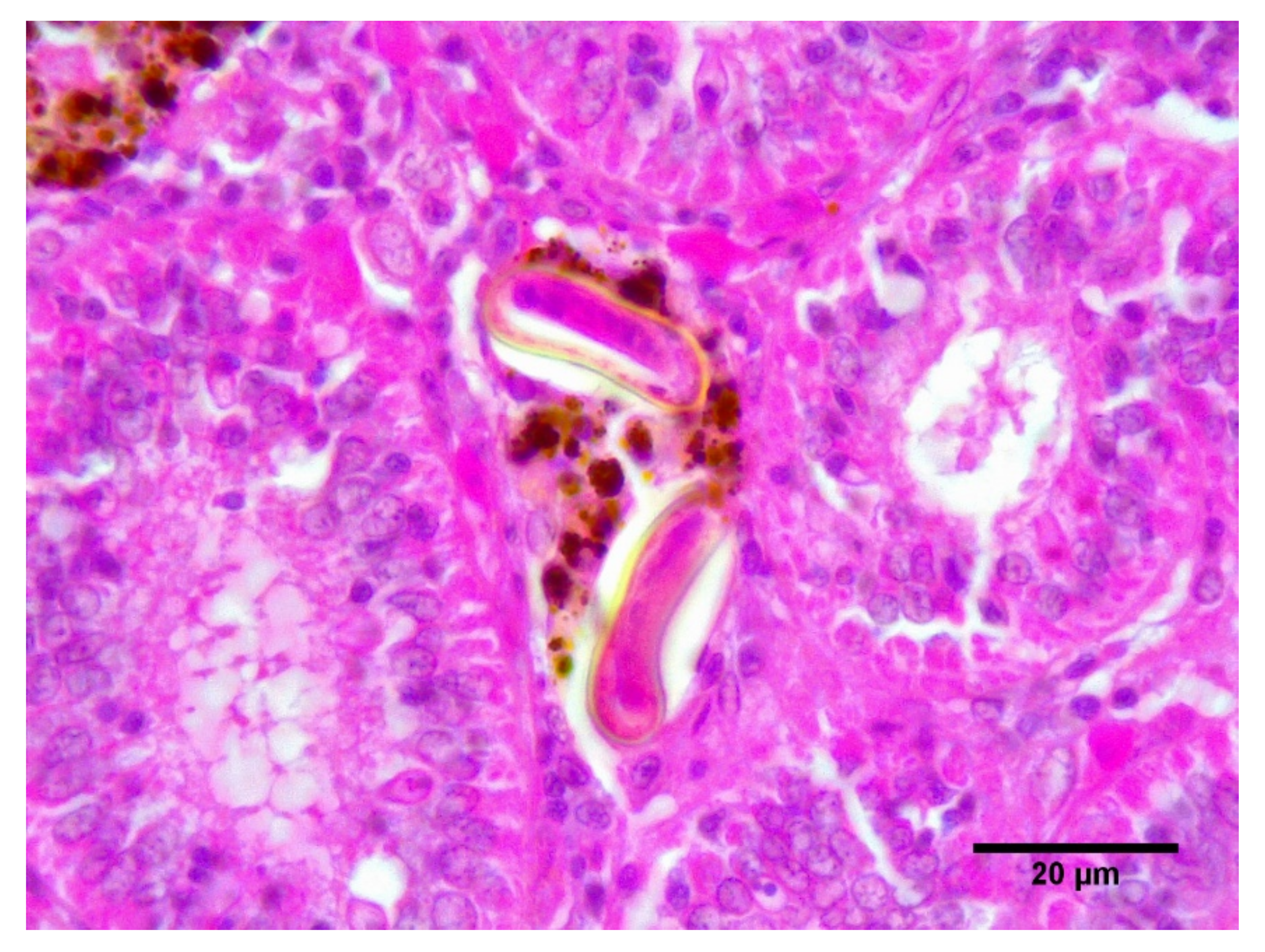
2.1. Blood Fluke Infection
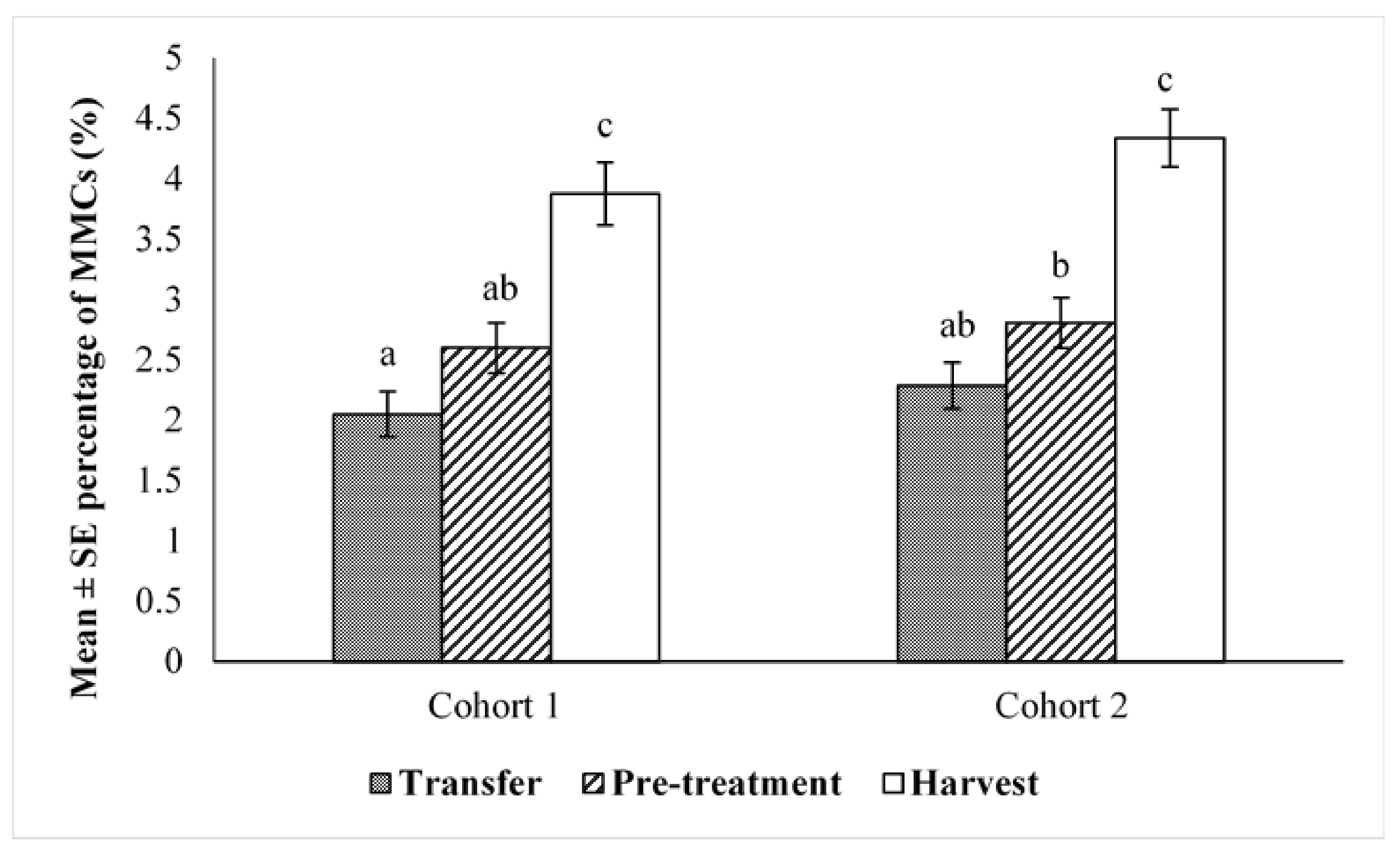
2.2. Effect of Husbandry on MMCs
2.3. Relationship between MMCs and Infection with Cardicola spp.
2.4. Comparison of Cohorts at Transfer
3. Discussion
4. Materials and Methods
4.1. Fish
4.2. Sampling Design and Sample Collection
4.3. Severity of Infection Analysis
4.4. Image Capture and Analysis
4.5. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Steinel, N.C.; Bolnick, D.I. Melanomacrophage centers as a histological indicator of immune function in fish and other poikilotherms. Front. Immunol. 2017, 8, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigliano, F.A.; Bermúdez, R.; Quiroga, M.I.; Nieto, J.M. Evidence for melano-macrophage centres of teleost as evolutionary precursors of germinal centres of higher vertebrates: An immunohistochemical study. Fish Shellfish Immunol. 2006, 21, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, N.T.; Leef, M.J.; Valdenegro, V.; Hayward, C.J.; Nowak, B.F. Correlation of humoral immune response in Southern Bluefin Tuna, T. maccoyii, with infection stage of the blood fluke, Cardicola forsteri. PLoS ONE 2012, 7, e45742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, M.M.; Landos, M.; D’Antignana, T. Case–control study of epidemic mortality and Cardicola forsteri–associated disease in farmed Southern Bluefin Tuna (Thunnus maccoyii) of South Australia. Vet. Pathol. 2010, 48, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Power, C.; Nowak, B.F.; Cribb, T.H.; Bott, N.J. Bloody flukes: A review of aporocotylids as parasites of cultured marine fishes. Int. J. Parasitol. 2020, 50, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.; Nowak, B.F. Effect of ranching time on melanomacrophage centres in anterior kidney and spleen of Southern Bluefin Tuna, Thunnus maccoyii. Fish Shellfish Immunol. 2016, 59, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Widdicombe, M.; Power, C.; van Gelderen, R.; Nowak, B.F.; Bott, N.J. Relationship between Southern Bluefin Tuna, Thunnus maccoyii, melanomacrophage centres and Cardicola spp. (Trematoda: Aporocotylidae) infection. Fish Shellfish Immunol. 2020, 106, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Power, C.; Webber, C.; Rough, K.; Staunton, R.; Nowak, B.F.; Bott, N.J. The effect of different treatment strategies on Cardicola spp. (Trematoda: Aporocotylidae) infection in ranched Southern Bluefin Tuna (Thunnus maccoyii) from Port Lincoln, South Australia. Aquaculture 2019, 513, 734401. [Google Scholar] [CrossRef]
- Herbert, B.W.; Shaharom, F.M.; Anderson, I.G. Histopathology of cultured sea bass (Lates calcarifer) (Centropomidae) infected with Cruoricola lates (Trematoda: Sanguinicolidae) from Pulau Ketam, Malaysia. Int. J. Parasitol. 1995, 25, 3–13. [Google Scholar] [CrossRef]
- McElroy, E.J.; Nowak, B.; Hill-Spanik, K.M.; Granath, W.O.; Connors, V.A.; Driver, J.; Tucker, C.J.; Kyle, D.E.; de Buron, I. Dynamics of infection and pathology induced by the aporocotylid, Cardicola laruei, in Spotted Seatrout, Cynoscion nebulosus (Sciaenidae). Int. J. Parasitol. 2020, 50, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, R.M.; Thulin, J. Response by Plecropomus leopardus and other serranid fishes to Pearsonellum corventum (Digenea: Sanguinicolidae), including melanomacrophage centres in the heart. Aust. J. Zool. 1989, 37, 129–142. [Google Scholar] [CrossRef]
- Neumann, L.; Bridle, A.; Leef, M.; Nowak, B.F. Annual variability of infection with Cardicola forsteri and Cardicola orientalis in ranched and wild Southern Bluefin Tuna (Thunnus maccoyii). Aquaculture 2018, 487, 1–6. [Google Scholar] [CrossRef]
- Polinski, M.; Hamilton, D.B.; Nowak, B.; Bridle, A. SYBR, TaqMan, or both: Highly sensitive, non-invasive detection of Cardicola blood fluke species in Southern Bluefin Tuna (Thunnus maccoyii). Mol. Biochem. Parasitol. 2013, 191, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Shirakashi, S.; Andrews, M.; Kishimoto, Y.; Ishimaru, K.; Okada, T.; Sawada, Y.; Ogawa, K. Oral treatment of praziquantel as an effective control measure against blood fluke infection in Pacific Bluefin Tuna (Thunnus orientalis). Aquaculture 2012, 15–19. [Google Scholar] [CrossRef]
- Aiken, H.; Hayward, C.; Nowak, B.F. An epizootic and its decline of a blood fluke, Cardicola forsteri, in farmed Southern Bluefin Tuna, Thunnus maccoyii. Aquaculture 2006, 254, 40–45. [Google Scholar] [CrossRef]
- Guiard, V.; Rasch, D. The robustness of two sample tests for means: A reply on von Eye’s comment. Psychol. Sci. 2004, 46, 549–554. [Google Scholar]
- Ruxton, G.D. The unequal variance t-test is an underused alternative to Student’s t-test and the Mann–Whitney U test. Behav. Ecol. 2006, 17, 688–690. [Google Scholar] [CrossRef]
- Underwood, A. Techniques of analysis of variance in experimental marine biology and ecology. Oceanogr. Mar. Biol. Ann. Rev. 1981, 19, 513–605. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Infection Measure | Transfer C1 C2 | Pre-Treatment C1 C2 | Harvest C1 C2 |
|---|---|---|---|
| Adult blood fluke heart, prevalence (%), (intensity of infection) | 0 0 | 6.67 73 (1) (4.5) | 73 80 (3) (1) |
| Heart C. forsteri qPCR, prevalence (%), (mean copy number ×103, SE ×103) | 53 67 (114, 8) ab (9, 6) a | 27 40 (9,6) a (12, 7) a | 73 53 (915, 481) b (34, 27) a |
| Gills C. forsteri qPCR, prevalence (%), (mean copy number ×103, SE ×103) | 67 40 (47, 28) a (61, 36) a | 53 27 (126, 85) ab (29, 20) a | 100 93 (1103, 334) b (541, 495) ab |
| Gills C. orientalis qPCR prevalence (%), (mean copy number ×103, SE ×103) | 20 13 (2, 2) (7, 7) | 27 13 (412, 418) (11, 12) | 0 7 (6, 6) |
| Severity of Infection (Organ) | P | R |
|---|---|---|
| C. forsteri qPCR (gills) * | 0.047 | 0.211 |
| C. orientalis qPCR (gills) | 0.210 | −0.134 |
| Cardicola spp. qPCR (gills) | 0.335 | 0.103 |
| Cardicola spp. Adult—flush and count (heart) | 0.075 | 0.189 |
| C. forsteri qPCR (heart) | 0.883 | −0.016 |
| Parasite | T | P |
|---|---|---|
| C. forsteri heart (qPCR) | 1.57 | 0.13 |
| C. forsteri gills (qPCR) | 0.32 | 0.75 |
| C. orientalis gills (qPCR) | 0.15 | 0.50 |
| Hexostoma thynni (count) | 0.43 | 0.67 |
| Euryphorus brachypterus (count) | 1.78 | 0.09 |
| Pseudocycnus appendiculatus (count) | 0.31 | 0.76 |
| Sampling Time Point | Cohort 1 | Cohort 2 |
|---|---|---|
| Transfer | 20 January 2016 | 18 February 2016 |
| Pre-treatment | 16 February 2016 | 21 March 2016 |
| Harvest | 1 July 2017 | 28 June 2016 |
| Variable | Cohort 1 | Cohort 2 |
|---|---|---|
| Transfer date | 21 January 2016 | 18 February 2016 |
| Average weight at transfer (pontoon) (kg) | 16.2 | 18.5 |
| Average weight at transfer (sampled fish) (kg) | 15.1 | 15.7 |
| Average length at transfer (sampled fish) (cm) | 92.3 | 92.4 |
| Hexostoma thynni at transfer (%) | 2.6 | 1.7 |
| Pseudocycnus appendiculatus at transfer (%) | 0.4 | 0.5 |
| Euryphorus brachypterus at transfer (%) | 4.3 | 1.1 |
| Praziquantel treatment date | 17 February 2016 | 22 March 2016 |
| Mortality to harvest (%) * | 3.7 | 4.6 |
| Average weight at harvest (kg) | 32.3 | 32.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, B.F.; Dang, M.; Webber, C.; Neumann, L.; Bridle, A.; Bermudez, R.; Evans, D. Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections. Pathogens 2021, 10, 79. https://doi.org/10.3390/pathogens10010079
Nowak BF, Dang M, Webber C, Neumann L, Bridle A, Bermudez R, Evans D. Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections. Pathogens. 2021; 10(1):79. https://doi.org/10.3390/pathogens10010079
Chicago/Turabian StyleNowak, Barbara F., Mai Dang, Claire Webber, Lukas Neumann, Andrew Bridle, Roberto Bermudez, and Daryl Evans. 2021. "Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections" Pathogens 10, no. 1: 79. https://doi.org/10.3390/pathogens10010079
APA StyleNowak, B. F., Dang, M., Webber, C., Neumann, L., Bridle, A., Bermudez, R., & Evans, D. (2021). Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections. Pathogens, 10(1), 79. https://doi.org/10.3390/pathogens10010079