
The Toxicity of Wiped Dust and Airborne Microbes in Individual Classrooms Increase the Risk of Teachers’ Work-Related Symptoms: A Cross-Sectional Study


Abstract
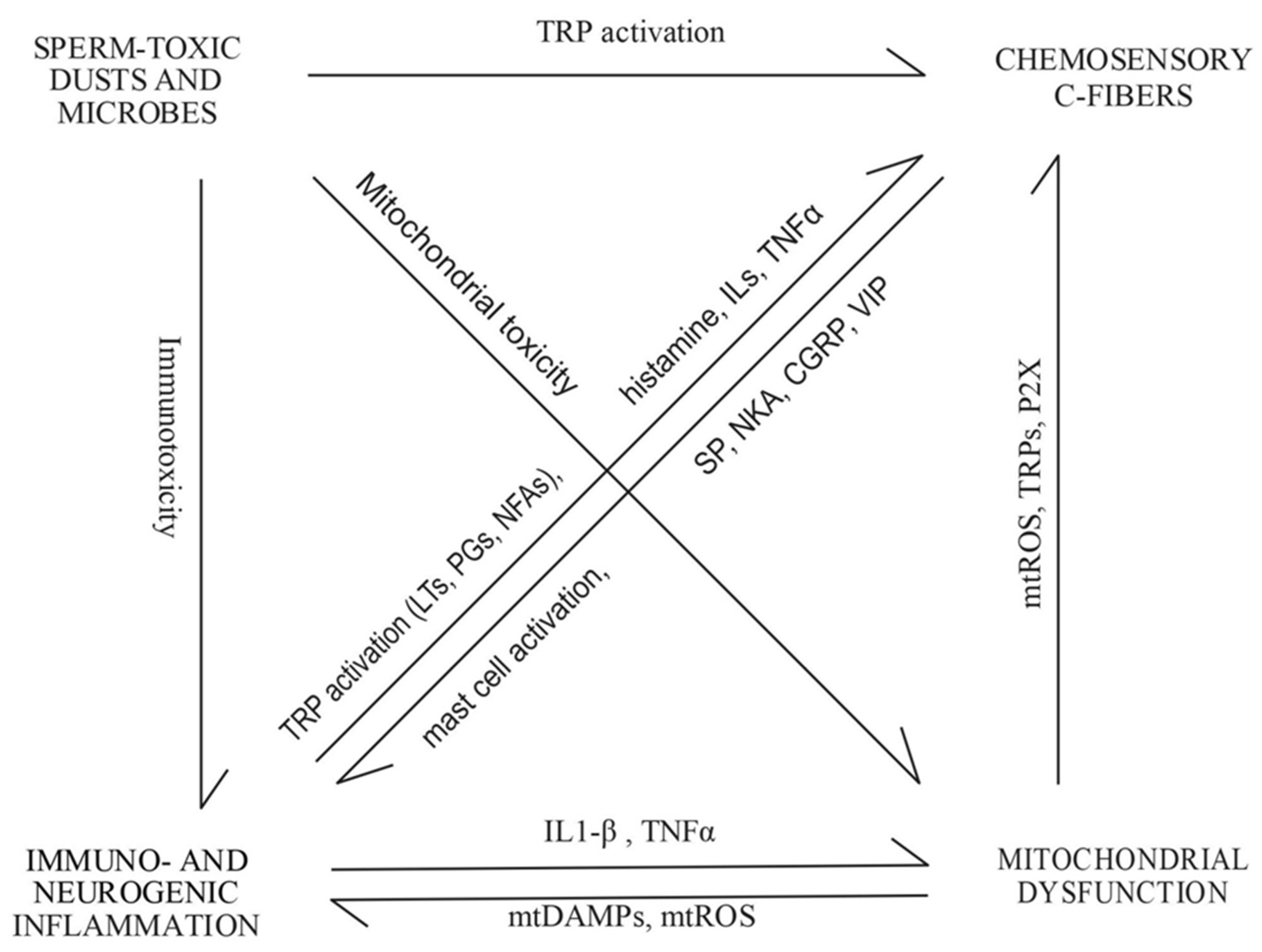
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Schools
4.2. Teachers
4.3. Indoor Samples
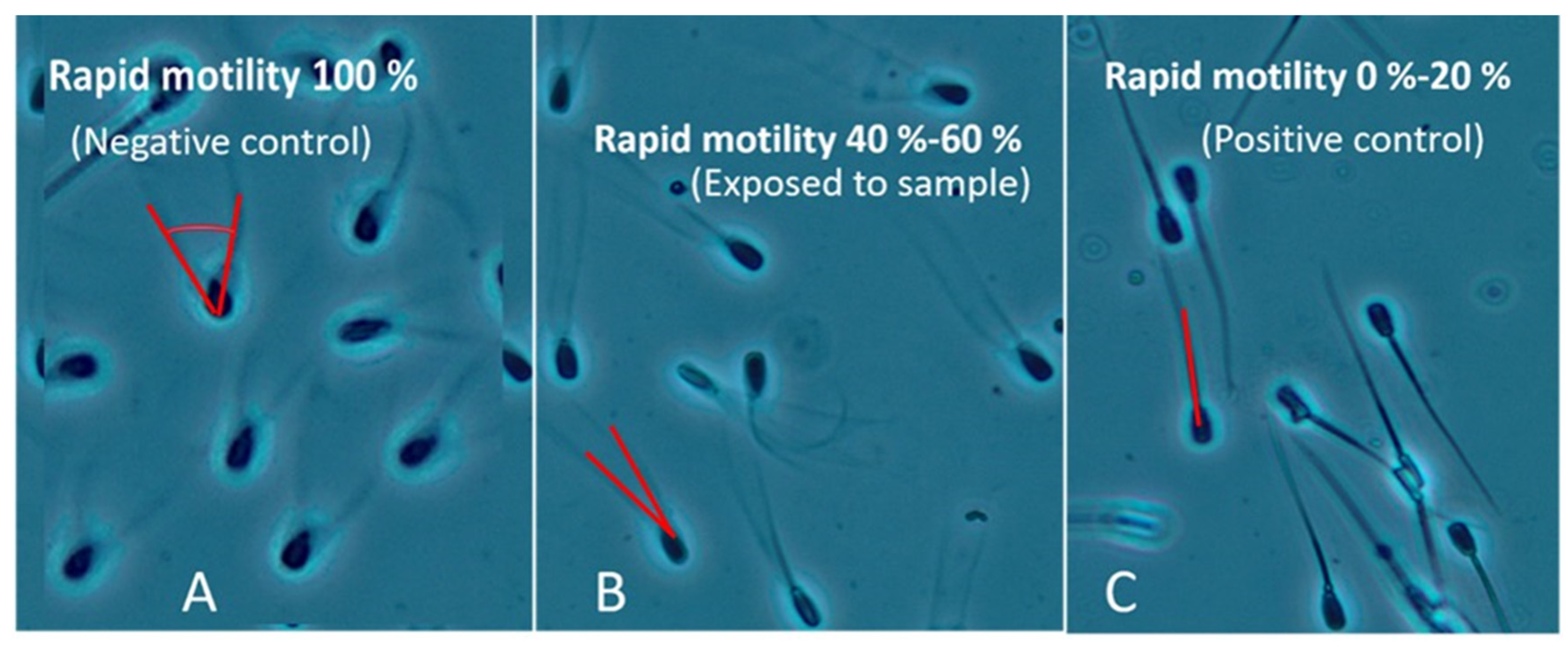
4.4. Toxicity Assay
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BRS | building-related symptoms |
| BSMI | boar sperm motility inhibition assay |
| CGRP | calcitonin gene-related peptide |
| CI | confidence interval |
| EC50 | half maximal effective concentration |
| IL | interleukin |
| IL-1β | interleukin 1beta |
| LPS | lipopolysaccharide |
| LT | leukotriene |
| MEA | malt extract agar |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| mtDAMP | mitochondrial damage-associated molecular pattern |
| mtROS | mitochondrial-derived reactive oxygen species |
| NFA | nitrated fatty acid |
| NKA | neurokinin A |
| NLRP3 | NLR family pyrin domain containing 3 |
| OR | odds ratio |
| PG | prostaglandin |
| P2X | purinergic 2X receptor |
| SP | substance P |
| SD | standard deviation |
| TNFα | tumor necrosis factor alpha |
| TRP | transient receptor potential |
| VIP | vasoactive intestinal peptide |
References
- World Health Organisation. WHO Guidelines for Indoor Air Quality: Dampness and Mould; World Health Organization: Geneva, Swithzerland, 2009; pp. 93–95.
- Burge, S.; Hedge, A.; Wilson, S.; Bass, J.H.; Robertson, A. Sick building syndrome: A study of 4373 office workers. Ann. Occup. Hyg. 1987, 31, 493–504. [Google Scholar]
- World Health Organisation. Indoor Air Pollutants: Exposure and Health Effects; EURO Reports and Studies 78; World Health Organization: Copenhagen, Denmark, 1983; pp. 23, 25.
- Mendell, M.J.; Mirer, A.G.; Cheung, K.; Tong, M.; Douwes, J. Respiratory and allergic health effects of dampness, mold, and dampness-related agents: A review of the epidemiologic evidence. Environ. Health Perspect. 2011, 119, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Quansah, R.; Jaakkola, M.S.; Hugg, T.T.; Heikkinen, S.A.; Jaakkola, J.J. Residential dampness and molds and the risk of developing asthma: A systematic review and meta-analysis. PLoS ONE 2012, 7, e47526. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, M.S.; Quansah, R.; Hugg, T.T.; Heikkinen, S.A.; Jaakkola, J.J. Association of indoor dampness and molds with rhinitis risk: A systematic review and meta-analysis. J. Allergy Clin. Immunol. 2013, 132, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Engelhart, S.; Loock, A.; Skutlarek, D.; Sagunski, H.; Lommel, A.; Färber, H.; Exner, M. Occurrence of toxigenic Aspergillus versicolor isolates and sterigmatocystin in carpet dust from damp indoor environments. Appl. Environ. Microbiol. 2002, 68, 3886–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirvonen, M.R.; Huttunen, K.; Roponen, M. Bacterial strains from moldy buildings are highly potent inducers of inflammatory and cytotoxic effects. Indoor Air 2005, 15, 65–70. [Google Scholar] [CrossRef]
- Rasimus-Sahari, S.; Teplova, V.V.; Andersson, M.A.; Mikkola, R.; Kankkunen, P.; Matikainen, S.; Gahmberg, C.G.; Andersson, L.C.; Salkinoja-Salonen, M. The peptide toxin amylosin of Bacillus amyloliquefaciens from moisture-damaged buildings is immunotoxic, induces potassium efflux from mammalian cells, and has antimicrobial activity. Appl. Environ. Microbiol. 2015, 81, 2939–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salo, J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salo, J.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. cochliodes and Ch. rectangulare during the Diversity Tracking of Mycotoxin-Producing Chaetomium-Like Isolates Obtained in Buildings in Finland. Toxins 2020, 12, 443. [Google Scholar] [CrossRef]
- Brasel, T.L.; Martin, J.M.; Carriker, C.G.; Wilson, S.C.; Straus, D.C. Detection of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins in the indoor environment. Appl. Environ. Microbiol. 2005, 71, 7376–7388. [Google Scholar] [CrossRef] [Green Version]
- Lemons, A.R.; Croston, T.L.; Goldsmith, W.T.; Barnes, M.A.; Jaderson, M.A.; Park, J.H.; McKinney, W.; Beezhold, D.H.; Green, B.J. Cultivation and aerosolization of Stachybotrys chartarum for modeling pulmonary inhalation exposure. Inhal. Toxicol. 2019, 31, 446–456. [Google Scholar] [CrossRef]
- Brasel, T.L.; Douglas, D.R.; Wilson, S.C.; Straus, D.C. Detection of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins on particulates smaller than conidia. Appl. Environ. Microbiol. 2005, 71, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Rasimus, S.; Hoornstra, D.; Salin, P.; Rahkila, R.; Heikkinen, M.; Mattila, S.; Peltola, J.; Kalso, S.; et al. Boar spermatozoa as a biosensor for detecting toxic substances in indoor dust and aerosols. Toxicol. Vitr. 2010, 24, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Aleksic, B.; Draghi, M.; Ritoux, S.; Bailly, S.; Lacroix, M.; Oswald, I.P.; Bailly, J.D.; Robine, E. Aerosolization of Mycotoxins after Growth of Toxinogenic Fungi on Wallpaper. Appl. Environ. Microbiol. 2017, 83, e01001-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gareis, M.; Gareis, E.M. Guttation droplets of Penicillium nordicum and Penicillium verrucosum contain high concentrations of the mycotoxins ochratoxin A and B. Mycopathologia 2007, 163, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Gareis, M.; Gottschalk, C. Stachybotrys spp. and the guttation phenomenon. Mycotoxin Res. 2014, 30, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.A.; Salo, J.; Kedves, O.; Kredics, L.; Druzhinina, I.; Kurnitski, J.; Salonen, H. Bioreactivity, Guttation and Agents Influencing Surface Tension of Water Emitted by Actively Growing Indoor Mould Isolates. Microorganisms 2020, 8, E1940. [Google Scholar] [CrossRef]
- Bloom, E.; Nyman, E.; Must, A.; Pehrson, C.; Larsson, L. Molds and mycotoxins in indoor environments--a survey in water-damaged buildings. J. Occup. Environ. Hyg. 2009, 6, 671–678. [Google Scholar] [CrossRef]
- Täubel, M.; Sulyok, M.; Vishwanath, V.; Bloom, E.; Turunen, M.; Järvi, K.; Kauhanen, E.; Krska, R.; Hyvärinen, A.; Larsson, L.; et al. Co-occurrence of toxic bacterial and fungal secondary metabolites in moisture-damaged indoor environments. Indoor Air 2011, 21, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Peitzsch, M.; Sulyok, M.; Täubel, M.; Vishwanath, V.; Krop, E.; Borràs-Santos, A.; Hyvärinen, A.; Nevalainen, A.; Krska, R.; Larsson, L. Microbial secondary metabolites in school buildings inspected for moisture damage in Finland, The Netherlands and Spain. J. Environ. Monit. 2012, 14, 2044–2053. [Google Scholar] [CrossRef] [Green Version]
- Došen, I.; Andersen, B.; Phippen, C.B.; Clausen, G.; Nielsen, K.F. Stachybotrys mycotoxins: From culture extracts to dust samples. Anal. Bioanal. Chem. 2016, 408, 5513–5526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Sulyok, M.; Lemons, A.R.; Green, B.J.; Cox-Ganser, J.M. Characterization of fungi in office dust: Comparing results of microbial secondary metabolites, fungal internal transcribed spacer region sequencing, viable culture and other microbial indices. Indoor Air 2018. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Viegas, C.; Almeida, B.; Monteiro, A.; Paciência, I.; Rufo, J.C.; Carolino, E.; Quintal-Gomes, A.; Twarużek, M.; Kosicki, R.; Marchand, G.; et al. Settled dust assessment in clinical environment: Useful for the evaluation of a wider bioburden spectrum. Int. J. Environ. Health Res. 2019, 1–19. [Google Scholar] [CrossRef]
- Viegas, S.; Viegas, C.; Martins, C.; Assunção, R. Occupational Exposure to Mycotoxins-Different Sampling Strategies Telling a Common Story Regarding Occupational Studies Performed in Portugal (2012–2020). Toxins 2020, 12, 513. [Google Scholar] [CrossRef]
- Polizzi, V.; Delmulle, B.; Adams, A.; Moretti, A.; Susca, A.; Picco, A.M.; Rosseel, Y.; Kindt, R.; Van Bocxlaer, J.; De Kimpe, N.; et al. JEM Spotlight: Fungi, mycotoxins and microbial volatile organic compounds in mouldy interiors from water-damaged buildings. J. Environ. Monit. 2009, 11, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Allermann, L.; Pejtersen, J.; Gunnarsen, L.; Poulsen, O.M. Building-related symptoms and inflammatory potency of dust from office buildings. Indoor Air 2007, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Tirkkonen, J.; Täubel, M.; Hirvonen, M.R.; Leppänen, H.; Lindsley, W.G.; Chen, B.T.; Hyvärinen, A.; Huttunen, K. Evaluation of sampling methods for toxicological testing of indoor air particulate matter. Inhal. Toxicol. 2016, 28, 500–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roponen, M.; Meklin, T.; Rintala, H.; Hyvärinen, A.; Hirvonen, M.R. Effect of moisture-damage intervention on the immunotoxic potential and microbial content of airborne particles and on occupants’ upper airway inflammatory responses. Indoor Air 2013, 23, 295–302. [Google Scholar] [CrossRef]
- Huttunen, K.; Tirkkonen, J.; Täubel, M.; Krop, E.; Mikkonen, S.; Pekkanen, J.; Heederik, D.; Zock, J.P.; Hyvärinen, A.; Hirvonen, M.R. Inflammatory potential in relation to the microbial content of settled dust samples collected from moisture-damaged and reference schools: Results of HITEA study. Indoor Air 2016, 26, 380–390. [Google Scholar] [CrossRef]
- Allermann, L.; Meyer, H.W.; Poulsen, O.M.; Nielsen, J.B.; Gyntelberg, F. Inflammatory potential of dust from schools and building related symptoms. Occup. Environ. Med. 2003, 60, E5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttunen, K.; Rintala, H.; Hirvonen, M.R.; Vepsalainen, A.; Hyvarinen, A.; Meklin, T.; Toivola, M.; Nevalainen, A. Indoor air particles and bioaerosols before and after renovation of moisture-damaged buildings: The effect on biological activity and microbial flora. Environ. Res. 2008, 107, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Tirkkonen, J.; Täubel, M.; Leppänen, H.; Peltonen, M.; Lindsley, W.; Chen, B.T.; Hyvärinen, A.; Hirvonen, M.R.; Huttunen, K. Toxicity of airborne dust as an indicator of moisture problems in school buildings. Inhal. Toxicol. 2017, 29, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, K.; Wlodarczyk, A.J.; Tirkkonen, J.; Mikkonen, S.; Täubel, M.; Krop, E.; Jacobs, J.; Pekkanen, J.; Heederik, D.; Zock, J.P.; et al. Oxidative capacity and hemolytic activity of settled dust from moisture-damaged schools. Indoor Air 2019, 29, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.A.; Nikulin, M.; Koljalg, U.; Andersson, M.C.; Rainey, F.; Reijula, K.; Hintikka, E.L.; Salkinoja-Salonen, M. Bacteria, molds, and toxins in water-damaged building materials. Appl. Environ. Microbiol. 1997, 63, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Kroppenstedt, R.M.; Rainey, F.A.; Peltola, J.; Helin, J.; Sivonen, K.; Salkinoja-Salonen, M.S. The mitochondrial toxin produced by Streptomyces griseus strains isolated from an indoor environment is valinomycin. Appl. Environ. Microbiol. 1998, 64, 4767–4773. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.A.; Mikkola, R.; Raulio, M.; Kredics, L.; Maijala, P.; Salkinoja-Salonen, M.S. Acrebol, a novel toxic peptaibol produced by an Acremonium exuviarum indoor isolate. J. Appl. Microbiol. 2009, 106, 909–923. [Google Scholar] [CrossRef]
- Kankkunen, P.; Rintahaka, J.; Aalto, A.; Leino, M.; Majuri, M.L.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate inflammatory response in human macrophages. J. Immunol. 2009, 182, 6418–6425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-Residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+ -permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Hautaniemi, M.; Salkinoja-Salonen, M.S. Toxic indole alkaloids avrainvillamide and stephacidin B produced by a biocide tolerant indoor mold Aspergillus westerdijkiae. Toxicon 2015, 99, 58–67. [Google Scholar] [CrossRef]
- Peltola, J.; Andersson, M.A.; Haahtela, T.; Mussalo-Rauhamaa, H.; Rainey, F.A.; Kroppenstedt, R.M.; Samson, R.A.; Salkinoja-Salonen, M.S. Toxic-metabolite-producing bacteria and fungus in an indoor environment. Appl. Environ. Microbiol. 2001, 67, 3269–3274. [Google Scholar] [CrossRef] [Green Version]
- Peltola, J.; Niessen, L.; Nielsen, K.F.; Jarvis, B.B.; Andersen, B.; Salkinoja-Salonen, M.; Möller, E.M. Toxigenic diversity of two different RAPD groups of Stachybotrys chartarum isolates analyzed by potential for trichothecene production and for boar sperm cell motility inhibition. Can. J. Microbiol. 2002, 48, 1017–1029. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Heinonen, M.; Salkinoja-Salonen, M.S. The toxic mode of action of cyclic lipodepsipeptide fusaricidins, produced by Paenibacillus polymyxa, toward mammalian cells. J. Appl. Microbiol. 2017, 123, 436–449. [Google Scholar] [CrossRef]
- Salo, J.; Marik, T.; Bencsik, O.; Mikkola, R.; Kredics, L.; Szekeres, A.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Screening Mold Colonies by Using Two Toxicity Assays Revealed Indoor Strains of Aspergillus calidoustus Producing Ophiobolins G and K. Toxins 2019, 11, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajao, C.; Andersson, M.A.; Teplova, V.V.; Nagy, S.; Gahmberg, C.G.; Andersson, L.C.; Hautaniemi, M.; Kakasi, B.; Roivainen, M.; Salkinoja-Salonen, M. Mitochondrial toxicity of triclosan on mammalian cells. Toxicol. Rep. 2015, 2, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toxicities of Detergents Used in Cleaning Chemicals and Hygiene Products in a Test Battery of Ex Vivo and In Vitro Assays. Updated 2019. Available online: https://www.researchgate.net/publication/332180207_FSdetergentpaper (accessed on 12 April 2021).
- Hyvönen, S.; Syrjala, H. Asthma Case Cluster during Renovation of a Water-Damaged and Toxic Building. Microorganisms 2019, 7, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salin, J.T.; Salkinoja-Salonen, M.; Salin, P.J.; Nelo, K.; Holma, T.; Ohtonen, P.; Syrjälä, H. Building-related symptoms are linked to the in vitro toxicity of indoor dust and airborne microbial propagules in schools: A cross-sectional study. Environ. Res. 2017, 154, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Salin, J.T.; Ohtonen, P.; Syrjälä, H. Teachers’ work-related non-literature-known building-related symptoms are also connected to indoor toxicity: A cross-sectional study. Indoor Air 2021, 31, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, O.; Papp, T.; Berta, M.; Zana, A.; Forgó, P.; Dombi, G.; Andersson, M.A.; Salkinoja-Salonen, M.; Vágvölgyi, C.; Szekeres, A. Ophiobolin A from Bipolaris oryzae perturbs motility and membrane integrities of porcine sperm and induces cell death on mammalian somatic cell lines. Toxins 2014, 6, 2857–2871. [Google Scholar] [CrossRef] [Green Version]
- Castagnoli, E.; Salo, J.; Toivonen, M.S.; Marik, T.; Mikkola, R.; Kredics, L.; Vicente-Carrillo, A.; Nagy, S.; Andersson, M.T.; Andersson, M.A.; et al. An Evaluation of Boar Spermatozoa as a Biosensor for the Detection of Sublethal and Lethal Toxicity. Toxins 2018, 10, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; McMullin, D.R. Fungal secondary metabolites as harmful indoor air contaminants: 10 years on. Appl. Microbiol. Biotechnol. 2014, 98, 9953–9966. [Google Scholar] [CrossRef] [PubMed]
- Straus, D.C. Molds, mycotoxins, and sick building syndrome. Toxicol. Ind. Health 2009, 25, 617–635. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Zhang, Y.; Jia, S.; Shang, H.; Fang, W.; Chen, D.; Fang, M. Human Indoor Exposome of Chemicals in Dust and Risk Prioritization Using EPA’s ToxCast Database. Environ. Sci. Technol. 2019, 53, 7045–7054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hsieh, J.H.; Zhu, H. Profiling animal toxicants by automatically mining public bioassay data: A big data approach for computational toxicology. PLoS ONE 2014, 9, e99863. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Teplova, V.V.; Saris, N.E.; Rainey, F.A.; Salkinoja-Salonen, M.S. Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Arch. Microbiol. 2004, 181, 314–323. [Google Scholar]
- Pieckova, E.; Wilkins, K. Airway toxicity of house dust and its fungal composition. Ann. Agric. Environ. Med. 2004, 11, 67–73. [Google Scholar]
- Holme, J.A.; Øya, E.; Afanou, A.K.J.; Øvrevik, J.; Eduard, W. Characterization and pro-inflammatory potential of indoor mold particles. Indoor Air 2020, 30, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Täubel, M.; Karvonen, A.M.; Reponen, T.; Hyvärinen, A.; Vesper, S.; Pekkanen, J. Application of the Environmental Relative Moldiness Index in Finland. Appl. Environ. Microbiol. 2015, 82, 578–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, B.; Frisvad, J.C.; Dunn, R.R.; Thrane, U. A Pilot Study on Baseline Fungi and Moisture Indicator Fungi in Danish Homes. J. Fungi 2021, 7, 71. [Google Scholar] [CrossRef]
- Vornanen-Winqvist, C.; Järvi, K.; Andersson, M.A.; Duchaine, C.; Létourneau, V.; Kedves, O.; Kredics, L.; Mikkola, R.; Kurnitski, J.; Salonen, H. Exposure to indoor air contaminants in school buildings with and without reported indoor air quality problems. Environ. Int. 2020, 141, 105781. [Google Scholar] [CrossRef]
- Andersson, M.A.; Mikkola, R.; Helin, J.; Andersson, M.C.; Salkinoja-Salonen, M. A novel sensitive bioassay for detection of Bacillus cereus emetic toxin and related depsipeptide ionophores. Appl. Environ. Microbiol. 1998, 64, 1338–1343. [Google Scholar] [CrossRef] [Green Version]
- Hoornstra, D.; Andersson, M.A.; Mikkola, R.; Salkinoja-Salonen, M.S. A new method for in vitro detection of microbially produced mitochondrial toxins. Toxicol. Vitr. 2003, 17, 745–751. [Google Scholar] [CrossRef]
- Vicente-Carrillo, A.; Edebert, I.; Garside, H.; Cotgreave, I.; Rigler, R.; Loitto, V.; Magnusson, K.E.; Rodríguez-Martínez, H. Boar spermatozoa successfully predict mitochondrial modes of toxicity: Implications for drug toxicity testing and the 3R principles. Toxicol. Vitr. 2015, 29, 582–591. [Google Scholar] [CrossRef]
- Teplova, V.V.; Belosludtsev, K.N.; Kruglov, A.G. Mechanism of triclosan toxicity: Mitochondrial dysfunction including complex II inhibition, superoxide release and uncoupling of oxidative phosphorylation. Toxicol. Lett 2017, 275, 108–117. [Google Scholar] [CrossRef]
- Dreier, D.A.; Mello, D.F.; Meyer, J.N.; Martyniuk, C.J. Linking Mitochondrial Dysfunction to Organismal and Population Health in the Context of Environmental Pollutants: Progress and Considerations for Mitochondrial Adverse Outcome Pathways. Environ. Toxicol. Chem. 2019, 38, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daiber, A.; Kuntic, M.; Hahad, O.; Delogu, L.G.; Rohrbach, S.; Di Lisa, F.; Schulz, R.; Münzel, T. Effects of air pollution particles (ultrafine and fine particulate matter) on mitochondrial function and oxidative stress-Implications for cardiovascular and neurodegenerative diseases. Arch. Biochem. Biophys. 2020, 696, 108662. [Google Scholar] [CrossRef]
- Li, N.; Sioutas, C.; Cho, A.; Schmitz, D.; Misra, C.; Sempf, J.; Wang, M.; Oberley, T.; Froines, J.; Nel, A. Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage. Environ. Health Perspect. 2003, 111, 455–460. [Google Scholar] [CrossRef]
- Meyer, J.N.; Leung, M.C.; Rooney, J.P.; Sendoel, A.; Hengartner, M.O.; Kisby, G.E.; Bess, A.S. Mitochondria as a target of environmental toxicants. Toxicol. Sci. 2013, 134, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zolkipli-Cunningham, Z.; Falk, M.J. Clinical effects of chemical exposures on mitochondrial function. Toxicology 2017, 391, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottis, A.; Herzig, S.; Auwerx, J. Mitocellular communication: Shaping health and disease. Science 2019, 366, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Moro, L. Mitochondria at the Crossroads of Physiology and Pathology. J. Clin. Med. 2020, 9, 1971. [Google Scholar] [CrossRef]
- West, A.P. Mitochondrial dysfunction as a trigger of innate immune responses and inflammation. Toxicology 2017, 391, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, S.M.; Kim, K.; Esteves, P.; Trian, T.; Barnes, P.J. Mitochondrial dysfunction in lung ageing and disease. Eur. Respir. Rev. 2020, 29, 200165. [Google Scholar] [CrossRef]
- Wiegman, C.H.; Michaeloudes, C.; Haji, G.; Narang, P.; Clarke, C.J.; Russell, K.E.; Bao, W.; Pavlidis, S.; Barnes, P.J.; Kanerva, J.; et al. COPDMAP Oxidative stress-induced mitochondrial dysfunction drives inflammation and airway smooth muscle remodeling in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2015, 136, 769–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Wang, L.; Liu, J.; Xie, F.; Su, B.; Wang, X. Abnormalities of Mitochondrial Dynamics in Neurodegenerative Diseases. Antioxidants 2017, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.W.; Aschner, M. Mitochondrial Redox Dysfunction and Environmental Exposures. Antioxid. Redox. Signal. 2015, 23, 578–595. [Google Scholar] [CrossRef]
- Fetterman, J.L.; Sammy, M.J.; Ballinger, S.W. Mitochondrial toxicity of tobacco smoke and air pollution. Toxicology 2017, 391, 18–33. [Google Scholar] [CrossRef]
- Manevski, M.; Muthumalage, T.; Devadoss, D.; Sundar, I.K.; Wang, Q.; Singh, K.P.; Unwalla, H.J.; Chand, H.S.; Rahman, I. Cellular stress responses and dysfunctional Mitochondrial-cellular senescence, and therapeutics in chronic respiratory diseases. Redox. Biol. 2020, 33, 101443. [Google Scholar] [CrossRef]
- Sundar, I.K.; Maremanda, K.P.; Rahman, I. Mitochondrial dysfunction is associated with Miro1 reduction in lung epithelial cells by cigarette smoke. Toxicol. Lett. 2019, 317, 92–101. [Google Scholar] [CrossRef]
- Sachdeva, K.; Do, D.C.; Zhang, Y.; Hu, X.; Chen, J.; Gao, P. Environmental Exposures and Asthma Development: Autophagy, Mitophagy, and Cellular Senescence. Front. Immunol. 2019, 10, 2787. [Google Scholar] [CrossRef] [PubMed]
- Korkalainen, M.; Täubel, M.; Naarala, J.; Kirjavainen, P.; Koistinen, A.; Hyvärinen, A.; Komulainen, H.; Viluksela, M. Synergistic proinflammatory interactions of microbial toxins and structural components characteristic to moisture-damaged buildings. Indoor Air 2017, 27, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Horssen, J.; van Schaik, P.; Witte, M. Inflammation and mitochondrial dysfunction: A vicious circle in neurodegenerative disorders? Neurosci. Lett. 2019, 710, 132931. [Google Scholar] [CrossRef]
- Mazzone, S.B.; Undem, B.J. Vagal Afferent Innervation of the Airways in Health and Disease. Physiol. Rev. 2016, 96, 975–1024. [Google Scholar] [CrossRef] [Green Version]
- Viana, F. Chemosensory properties of the trigeminal system. ACS Chem. Neurosci. 2011, 2, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Rasimus-Sahari, S.; Mikkola, R.; Andersson, M.A.; Jestoi, M.; Salkinoja-Salonen, M. Streptomyces strains producing mitochondriotoxic antimycin A found in cereal grains. Int. J. Food. Microbiol. 2016, 218, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.R.; Hadley, S.H.; Barannikov, I.; Ajmo, J.M.; Bahia, P.K.; Taylor-Clark, T.E. Antimycin A-induced mitochondrial dysfunction activates vagal sensory neurons via ROS-dependent activation of TRPA1 and ROS-independent activation of TRPV1. Brain Res. 2019, 1715, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Bahia, P.K.; Hadley, S.H.; Barannikov, I.; Sowells, I.; Kim, S.H.; Taylor-Clark, T.E. Antimycin A increases bronchopulmonary C-fiber excitability via protein kinase C alpha. Respir. Physiol. Neurobiol. 2020, 278, 103446. [Google Scholar] [CrossRef]
- Ruan, T.; Lin, Y.J.; Hsu, T.H.; Lu, S.H.; Jow, G.M.; Kou, Y.R. Sensitization by pulmonary reactive oxygen species of rat vagal lung C-fibers: The roles of the TRPV1, TRPA1, and P2X receptors. PLoS ONE 2014, 9, e91763. [Google Scholar] [CrossRef]
- Lehmann, R.; Schöbel, N.; Hatt, H.; van Thriel, C. The involvement of TRP channels in sensory irritation: A mechanistic approach toward a better understanding of the biological effects of local irritants. Arch. Toxicol. 2016, 90, 1399–1413. [Google Scholar] [CrossRef]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef] [PubMed]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019, 101, 412–420.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabata, H.; Artis, D. Neuro-immune crosstalk and allergic inflammation. J. Clin. Investig. 2019, 129, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Thapaliya, M.; Chompunud Na Ayudhya, C.; Amponnawarat, A.; Roy, S.; Ali, H. Mast Cell-Specific MRGPRX2: A Key Modulator of Neuro-Immune Interaction in Allergic Diseases. Curr. Allergy Asthma Rep. 2021, 21, 1–11. [Google Scholar] [CrossRef]
- Shouman, K.; Benarroch, E.E. Peripheral neuroimmune interactions: Selected review and some clinical implications. Clin. Auton. Res. 2021, 31, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Bessac, B.F.; Jordt, S.E. Breathtaking TRP channels: TRPA1 and TRPV1 in airway chemosensation and reflex control. Physiology 2008, 23, 360–370. [Google Scholar] [CrossRef]
- Tillie-Leblond, I.; Montani, D.; Crestani, B.; de Blic, J.; Humbert, M.; Tunon-de-Lara, M.; Magnan, A.; Roche, N.; Ostinelli, J.; Chanez, P. Relation between inflammation and symptoms in asthma. Allergy 2009, 64, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, A.M.; Azzegagh, Z.; Tuvim, M.J.; Dickey, B.F. Airway Mucin Secretion. Ann. Am. Thorac. Soc. 2018, 15, S164–S170. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Clark, T.E. Role of reactive oxygen species and TRP channels in the cough reflex. Cell Calcium 2016, 60, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Clark, T.E.; Undem, B.J. Sensing pulmonary oxidative stress by lung vagal afferents. Respir. Physiol. Neurobiol. 2011, 178, 406–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228.e13. [Google Scholar] [CrossRef] [Green Version]
- Diver, S.; Russell, R.J.; Brightling, C.E. Cough and Eosinophilia. J. Allergy. Clin. Immunol. Pract. 2019, 7, 1740–1747. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F. Approach to chronic cough: The neuropathic basis for cough hypersensitivity syndrome. J. Thorac. Dis. 2014, 6, S699–S707. [Google Scholar]
- Pecova, T.; Kocan, I.; Vysehradsky, R.; Pecova, R. Itch and Cough-Similar Role of Sensory Nerves in Their Pathogenesis. Physiol. Res. 2020, 69, S43–S54. [Google Scholar] [CrossRef]
- Smeitink, J.; Koene, S.; Beyrath, J.; Saris, C.; Turnbull, D.; Janssen, M. Mitochondrial Migraine: Disentangling the angiopathy paradigm in m.3243A>G patients. JIMD Rep. 2019, 46, 52–62. [Google Scholar] [PubMed]
- Conti, P.; D’Ovidio, C.; Conti, C.; Gallenga, C.E.; Lauritano, D.; Caraffa, A.; Kritas, S.K.; Ronconi, G. Progression in migraine: Role of mast cells and pro-inflammatory and anti-inflammatory cytokines. Eur. J. Pharmacol. 2019, 844, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.C.; Lisicki, M.; Fischer, D.; Sándor, P.S.; Schoenen, J. The metabolic face of migraine-from pathophysiology to treatment. Nat. Rev. Neurol. 2019, 15, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Dux, M.; Rosta, J.; Messlinger, K. TRP Channels in the Focus of Trigeminal Nociceptor Sensitization Contributing to Primary Headaches. Int. J. Mol. Sci. 2020, 21, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renner, B.; Mueller, C.A.; Shephard, A. Environmental and non-infectious factors in the aetiology of pharyngitis (sore throat). Inflamm. Res. 2012, 61, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Sundar, K.M.; Stark, A.C.; Hu, N.; Barkmeier-Kraemer, J. Is laryngeal hypersensitivity the basis of unexplained or refractory chronic cough? ERJ Open Res. 2021, 7, 00793–02020. [Google Scholar] [CrossRef]
- Wang, W.; Xian, M.; Xie, Y.; Zheng, J.; Li, J. Aggravation of airway inflammation and hyper-responsiveness following nasal challenge with Dermatophagoides pteronyssinus in perennial allergic rhinitis without symptoms of asthma. Allergy 2016, 71, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Eifan, A.O.; Durham, S.R. Pathogenesis of rhinitis. Clin. Exp. Allergy 2016, 46, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liang, C.; Wang, T.; Zou, Q.; Zhou, M.; Cheng, Y.; Peng, H.; Ji, Z.; Deng, Y.; Liao, J.; et al. NLRP3 inflammasome activation promotes the development of allergic rhinitis via epithelium pyroptosis. Biochem. Biophys. Res. Commun. 2020, 522, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Järvenpää, P.; Arkkila, P.; Aaltonen, L.M. Globus pharyngeus: A review of etiology, diagnostics, and treatment. Eur. Arch. Otorhinolaryngol. 2018, 275, 1945–1953. [Google Scholar] [CrossRef] [Green Version]
- Beuerman, R.W.; Stern, M.E. Neurogenic inflammation: A first line of defense for the ocular surface. Ocul. Surf. 2005, 3, S203–S206. [Google Scholar] [CrossRef]
- Ryder, E.C.; Benson, S. Conjunctivitis; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021; pp. 1–4. [Google Scholar]
- Ranjbaran, Z.; Keefer, L.; Stepanski, E.; Farhadi, A.; Keshavarzian, A. The relevance of sleep abnormalities to chronic inflammatory conditions. Inflamm. Res. 2007, 56, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Besedovsky, L.; Lange, T.; Haack, M. The Sleep-Immune Crosstalk in Health and Disease. Physiol. Rev. 2019, 99, 1325–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, V.; Della Marca, G.; Servidei, S.; Primiano, G. Sleep Disorders in Mitochondrial Diseases. Curr. Neurol. Neurosci. Rep. 2021, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gohy, S.T.; Hupin, C.; Pilette, C.; Ladjemi, M.Z. Chronic inflammatory airway diseases: The central role of the epithelium revisited. Clin. Exp. Allergy 2016, 46, 529–542. [Google Scholar] [CrossRef]
- Pieckova, E.; Jesenska, Z. Molds on house walls and the effect of their chloroform-extractable metabolites on the respiratory cilia movement of one-day-old chicks in vitro. Folia Microbiol. 1998, 43, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Pieckova, E.; Kunova, Z. Indoor fungi and their ciliostatic metabolites. Ann. Agric. Environ. Med. 2002, 9, 59–63. [Google Scholar] [PubMed]
- Mack, S.M.; Madl, A.K.; Pinkerton, K.E. Respiratory Health Effects of Exposure to Ambient Particulate Matter and Bioaerosols. Compr. Physiol. 2019, 10, 1–20. [Google Scholar]
- Lochhead, J.J.; Thorne, R.G. Intranasal delivery of biologics to the central nervous system. Adv. Drug Deliv. Rev. 2012, 64, 614–628. [Google Scholar] [CrossRef] [PubMed]
- Rojo, A.I.; Montero, C.; Salazar, M.; Close, R.M.; Fernández-Ruiz, J.; Sánchez-González, M.A.; de Sagarra, M.R.; Jackson-Lewis, V.; Cavada, C.; Cuadrado, A. Persistent penetration of MPTP through the nasal route induces Parkinson’s disease in mice. Eur. J. Neurosci. 2006, 24, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.R.; Raftis, J.B.; Langrish, J.P.; McLean, S.G.; Samutrtai, P.; Connell, S.P.; Wilson, S.; Vesey, A.T.; Fokkens, P.H.B.; Boere, A.J.F.; et al. Inhaled Nanoparticles Accumulate at Sites of Vascular Disease. ACS Nano 2017, 11, 4542–4552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baral, P.; Udit, S.; Chiu, I.M. Pain and immunity: Implications for host defence. Nat. Rev. Immunol. 2019, 19, 433–447. [Google Scholar] [CrossRef]
- Sotnichenko, A.; Pantsov, E.; Shinkarev, D.; Okhanov, V. Hydrophobized Reversed-Phase Adsorbent for Protection of Dairy Cattle against Lipophilic Toxins from Diet. Efficiensy in Vitro and in Vivo. Toxins 2019, 11, 256. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Total, N (%) | Work-Related, N (%) | Literature-Known BRS, Yes or No | |
|---|---|---|---|
| GENERAL SYMPTOMS | |||
| Fatigue | 88 (38.1) | 65 (28.1) | yes |
| Headache | 92 (39.8) | 46 (20.0) | yes |
| Fever | 13 (5.6) | 8 (3.5) | yes |
| Chills | 31 (13.4) | 19 (8.2) | yes |
| Generalized feeling of sickness | 60 (26.0) | 46 (20.0) | no |
| Decreased physical condition | 54 (23.4) | 27 (11.7) | no |
| Indefinite feeling of thermoregulation failure | 27 (11.7) | 19 (8.2) | no |
| RESPIRATORY SYMPTOMS | |||
| Nose stuffiness | 106 (45.9) | 64 (27.7) | yes |
| Nose dryness | 84 (36.4) | 52 (22.5) | yes |
| Nose stinging | 22 (9.5) | 7 (3.0) | yes |
| Bloody nasal discharge | 30 (13.0) | 16 (6.9) | yes |
| Runny nose | 74 (32.0) | 37 (16.0) | yes |
| Sneezing | 66 (28.6) | 39 (16.9) | yes |
| Mouth dryness | 48 (20.8) | 30 (13.0) | yes |
| Hoarseness | 88 (38.1) | 62 (26.8) | yes |
| Sore throat | 42 (18.2) | 27 (11.7) | yes |
| Wheezing | 15 (6.5) | 4 (1.7) | yes |
| Shortness of breath | 21 (9.1) | 12 (5.2) | yes |
| Asthma attacks | 12 (5.2) | 5 (2.2) | yes |
| Dry cough | 53 (22.9) | 40 (17.3) | yes |
| Pressure in the cheek | 42 (18.2) | 22 (9.5) | no |
| Globus sensation | 31 (13.4) | 19 (8.2) | no |
| Throat mucus | 70 (30.3) | 50 (21.6) | no |
| Throat itching | 49 (21.2) | 35 (15.2) | no |
| DERMAL SYMPTOMS | |||
| Skin dryness | 58 (25.1) | 20 (8.7) | yes |
| Exanthema | 13 (5.6) | 3 (1.3) | yes |
| Pruritus | 32 (13.9) | 12 (5.2) | yes |
| EYE SYMPTOMS | |||
| Eye irritation | 61 (26.4) | 39 (16.9) | yes |
| Wet eyes | 30 (13.0) | 15 (6.5) | yes |
| Dry eyes | 67 (29.0) | 45 (19.5) | yes |
| Swollen eyelids | 19 (8.2) | 9 (3.9) | yes |
| Red eyes | 32 (13.9) | 18 (7.8) | no |
| Eye discharge | 24 (10.4) | 14 (6.1) | no |
| HEARING SYMPTOMS | |||
| Difficulty distinguishing speech in noise | 51 (22.1) | 27 (11.7) | no |
| Hypersensitivity to sound | 28 (12.1) | 14 (6.1) | no |
| SLEEPING SYMPTOMS | |||
| Insomnia | 41 (17.7) | 25 (10.8) | yes |
| Difficulty falling asleep | 42 (18.2) | 21 (9.1) | yes |
| Increased need for sleep | 34 (14.7) | 18 (7.8) | no |
| MENTAL SYMPTOMS | |||
| Difficulty concentrating | 12 (5.2) | 4 (1.7) | yes |
| Mental irritability | 29 (12.6) | 22 (9.5) | no |
| Decreased stress resistance | 26 (11.3) | 18 (7.8) | no |
| BUILDING-RELATED DISEASES | |||
| Asthma | 22 (9.5) | a | yes |
| Allergic rhinitis | 32 (13.9) | a | yes |
| EC50 12 µg mL−1 | EC50 ≤ 6 µg mL−1 | |||
|---|---|---|---|---|
| OR (95% CI) | p Value | OR (95% CI) | p Value | |
| GENERAL SYMPTOMS | ||||
| Fatigue | 1.08 (0.46,2.53) | 0.86 | 3.00 (0.96,9.36) | 0.058 |
| Headache | 1.04 (0.41,2.65) | >0.90 | 1.79 (0.49,6.57) | 0.38 |
| Fever | 3.75 (0.63,22.2) | 0.15 | 2.59 (0.23,29.4) | 0.44 |
| Chills | 1.50 (0.41,5.56) | 0.54 | 1.22 (0.13,11.2) | 0.86 |
| Generalized feeling of sickness | 1.11 (0.42,2.94) | 0.84 | 3.28 (1.00,10.8) | 0.050 |
| Decreased physical condition | 1.08 (0.31,3.72) | 0.90 | 3.85 (0.99,15.0) | 0.052 |
| Indefinite feeling of thermoregulation failure | 0.66 (0.13,3.30) | 0.61 | 2.96 (0.65,13.4) | 0.16 |
| RESPIRATORY SYMPTOMS | ||||
| Nose stuffiness | 1.78 (0.76,4.15) | 0.18 | 4.08 (1.24,13.4) | 0.021 |
| Nose dryness | 1.65 (0.69,3.91) | 0.26 | 0.97 (0.24,4.01) | >0.90 |
| Nose stinging | 0.67 (0.07,6.37) | 0.73 | 1.67 (0.17,16.8) | 0.66 |
| Bloody nasal discharge | 3.33 (0.73,15.2) | 0.12 | 2.22 (0.21,23.0) | 0.51 |
| Runny nose | 4.24 (1.49,12.1) | 0.0070 | 6.93 (1.76,27.2) | 0.0056 |
| Sneezing | 1.17 (0.43,3.18) | 0.76 | 2.07 (0.55,7.75) | 0.28 |
| Mouth dryness | 1.99 (0.63,6.33) | 0.24 | 2.29 (0.49,10.5) | 0.29 |
| Hoarseness | 3.38 (1.47,7.75) | 0.0041 | 6.42 (1.95,21.1) | 0.0022 |
| Sore throat | 0.92 (0.17,4.95) | >0.90 | 3.80 (0.72,19.9) | 0.12 |
| Wheezing | 4.43 (0.21,95.2) | 0.34 | 7.56 (0.27,209) | 0.23 |
| Shortness of breath | 3.69 (0.76,17.9) | 0.10 | 12.2 (1.95,76.8) | 0.0076 |
| Asthma attacks | 7.07 (0.55,91.3) | 0.13 | 14.8 (1.00,219) | 0.050 |
| Dry cough | 2.14 (0.81,5.70) | 0.13 | 4.65 (1.29,16.8) | 0.019 |
| Pressure in the cheek | 0.36 (0.04,3.17) | 0.36 | 2.76 (0.46,16.6) | 0.27 |
| Globus sensation | 1.34 (0.23,7.72) | 0.74 | 9.02 (1.74,46.7) | 0.0088 |
| Throat mucus | 2.46 (0.99,6.14) | 0.053 | 7.64 (2.21,26.4) | 0.0013 |
| Throat itching | 2.19 (0.71,6.75) | 0.17 | 4.35 (1.08,17.6) | 0.039 |
| DERMAL SYMPTOMS | ||||
| Skin dryness | 0.66 (0.17,2.50) | 0.54 | 0.65 (0.07,5.55) | 0.69 |
| Exanthema | 3.45 (0.15,78.0) | 0.44 | 14.78 (0.36,606) | 0.16 |
| Pruritus | 0.63 (0.06,6.29) | 0.69 | 2.24 (0.19,26.2) | 0.52 |
| EYE SYMPTOMS | ||||
| Eye irritation | 0.93 (0.30,2.90) | >0.90 | 3.58 (0.89,14.4) | 0.072 |
| Wet eyes | 4.06 (0.65,25.3) | 0.13 | 12.7 (1.44,112) | 0.022 |
| Dry eyes | 0.86 (0.31,2.40) | 0.78 | 1.94 (0.52,7.28) | 0.32 |
| Swollen eyelids | 0.65 (0.07,6.28) | 0.71 | 2.29 (0.21,25.5) | 0.50 |
| Red eyes | 0.26 (0.03,2.46) | 0.24 | 2.34 (0.17,31.4) | 0.52 |
| Eye discharge | 0.80 (0.08,8.25) | 0.85 | 7.69 (0.96,61.4) | 0.054 |
| HEARING SYMPTOMS | ||||
| Difficulty distinguishing speech in noise | 0.14 (0.02,1.16) | 0.069 | 2.10 (0.45,9.69) | 0.34 |
| Hypersensitivity to sound | N.D. | 7.91 (1.70,36.8) | 0.0084 | |
| SLEEPING SYMPTOMS | ||||
| Insomnia | 1.00 (0.32,3.11) | >0.90 | 2.57 (0.59,11.2) | 0.21 |
| Difficulty falling asleep | 1.14 (0.32,4.02) | 0.84 | 7.58 (1.93,29.8) | 0.0038 |
| Increased need for sleep | 0.22 (0.03,1.76) | 0.15 | 7.74 (2.09,28.6) | 0.0022 |
| MENTAL SYMPTOMS | ||||
| Difficulty concentrating | N.D. | 17.8 (0.64,496) | 0.090 | |
| Mental irritability | 0.91 (0.27,3.10) | 0.88 | 2.53 (0.58,11.0) | 0.22 |
| Decreased stress resistance | 1.42 (0.39,5.18) | 0.60 | 3.33 (0.75,14.8) | 0.11 |
| BUILDING-RELATED DISEASES | ||||
| Asthma | 2.13 (0.59,7.76) | 0.25 | 1.44 (0.22,9.51) | 0.70 |
| Allergic rhinitis | 1.07 (0.33,3.44) | 0.90 | 4.64 (1.08,20.0) | 0.039 |
| EC50 ≤ 12 µg mL−1 | ||
|---|---|---|
| OR (95% CI) | p Value | |
| GENERAL SYMPTOMS | ||
| Fatigue | 1.63 (0.85,3.12) | 0.14 |
| Headache | 2.26 (1.06,4.79) | 0.034 |
| Fever | 2.80 (0.42,18.6) | 0.29 |
| Chills | 0.94 (0.31,2.85) | >0.90 |
| Generalized feeling of sickness | 1.81 (0.85,3.85) | 0.12 |
| Decreased physical condition | 1.47 (0.59,3.64) | 0.40 |
| Indefinite feeling of thermoregulation failure | 2.02 (0.68,5.96) | 0.20 |
| RESPIRATORY SYMPTOMS | ||
| Nose stuffiness | 2.19 (1.10,4.38) | 0.026 |
| Nose dryness | 2.17 (1.05,4.52) | 0.038 |
| Nose stinging a | N.D. | |
| Bloody nasal discharge | 2.34 (0.68,8.04) | 0.18 |
| Runny nose | 2.07 (0.92,4.66) | 0.79 |
| Sneezing | 1.50 (0.67,3.39) | 0.33 |
| Mouth dryness | 2.76 (1.13,6.71) | 0.026 |
| Hoarseness | 2.18 (1.12,4.26) | 0.022 |
| Sore throat | 2.81 (1.08,7.31) | 0.034 |
| Wheezing a | N.D. | |
| Shortness of breath | 4.79 (0.89,25.7) | 0.067 |
| Asthma attacks | 1.67 (0.11,24.5) | 0.71 |
| Dry cough | 1.10 (0.50,2.44) | 0.81 |
| Pressure in the cheek | 2.60 (0.89,7.59) | 0.079 |
| Globus sensation | 2.32 (0.76,7.05) | 0.14 |
| Throat mucus | 2.28 (1.09,4.74) | 0.028 |
| Throat itching | 2.22 (0.93,5.31) | 0.073 |
| DERMAL SYMPTOMS | ||
| Skin dryness | 3.02 (0.97,9.44) | 0.057 |
| Exanthema a | N.D. | |
| Pruritus | 0.90 (0.22,3.76) | 0.89 |
| EYE SYMPTOMS | ||
| Eye irritation | 2.07 (0.94,4.56) | 0.073 |
| Wet eyes | 2.97 (0.90,9.76) | 0.073 |
| Dry eyes | 1.73 (0.84,3.57) | 0.14 |
| Swollen eyelids | 2.23 (0.49,10.1) | 0.30 |
| Red eyes | 2.70 (0.91,7.99) | 0.072 |
| Eye discharge | 10.2 (2.03,50.9) | 0.0048 |
| HEARING SYMPTOMS | ||
| Difficulty distinguishing speech in noise | 1.52 (0.64,3.65) | 0.34 |
| Hypersensitivity to sound | 0.72 (0.20,2.56) | 0.61 |
| SLEEPING SYMPTOMS | ||
| Insomnia | 1.85 (0.70,4.91) | 0.22 |
| Difficulty falling asleep | 2.54 (0.87,7.46) | 0.089 |
| Increased need for sleep | 3.54 (1.03,12.2) | 0.045 |
| MENTAL SYMPTOMS | ||
| Difficulty concentrating | 1.68 (0.19,14.6) | 0.64 |
| Mental irritability | 1.10 (0.37,3.26) | 0.87 |
| Decreased stress resistance | 0.98 (0.31,3.06) | >0.90 |
| BUILDING-RELATED DISEASES | ||
| Asthma | 1.28 (0.43,3.77) | 0.66 |
| Allergic rhinitis | 1.59 (0.63,4.03) | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salin, J.; Ohtonen, P.; Andersson, M.A.; Syrjälä, H. The Toxicity of Wiped Dust and Airborne Microbes in Individual Classrooms Increase the Risk of Teachers’ Work-Related Symptoms: A Cross-Sectional Study. Pathogens 2021, 10, 1360. https://doi.org/10.3390/pathogens10111360
Salin J, Ohtonen P, Andersson MA, Syrjälä H. The Toxicity of Wiped Dust and Airborne Microbes in Individual Classrooms Increase the Risk of Teachers’ Work-Related Symptoms: A Cross-Sectional Study. Pathogens. 2021; 10(11):1360. https://doi.org/10.3390/pathogens10111360
Chicago/Turabian StyleSalin, Janne, Pasi Ohtonen, Maria A. Andersson, and Hannu Syrjälä. 2021. "The Toxicity of Wiped Dust and Airborne Microbes in Individual Classrooms Increase the Risk of Teachers’ Work-Related Symptoms: A Cross-Sectional Study" Pathogens 10, no. 11: 1360. https://doi.org/10.3390/pathogens10111360
APA StyleSalin, J., Ohtonen, P., Andersson, M. A., & Syrjälä, H. (2021). The Toxicity of Wiped Dust and Airborne Microbes in Individual Classrooms Increase the Risk of Teachers’ Work-Related Symptoms: A Cross-Sectional Study. Pathogens, 10(11), 1360. https://doi.org/10.3390/pathogens10111360