
Tissue Pathogens and Cancers: A Review of Commonly Seen Manifestations in Histo- and Cytopathology


{kind=link}
{kind=link}
{kind=link}
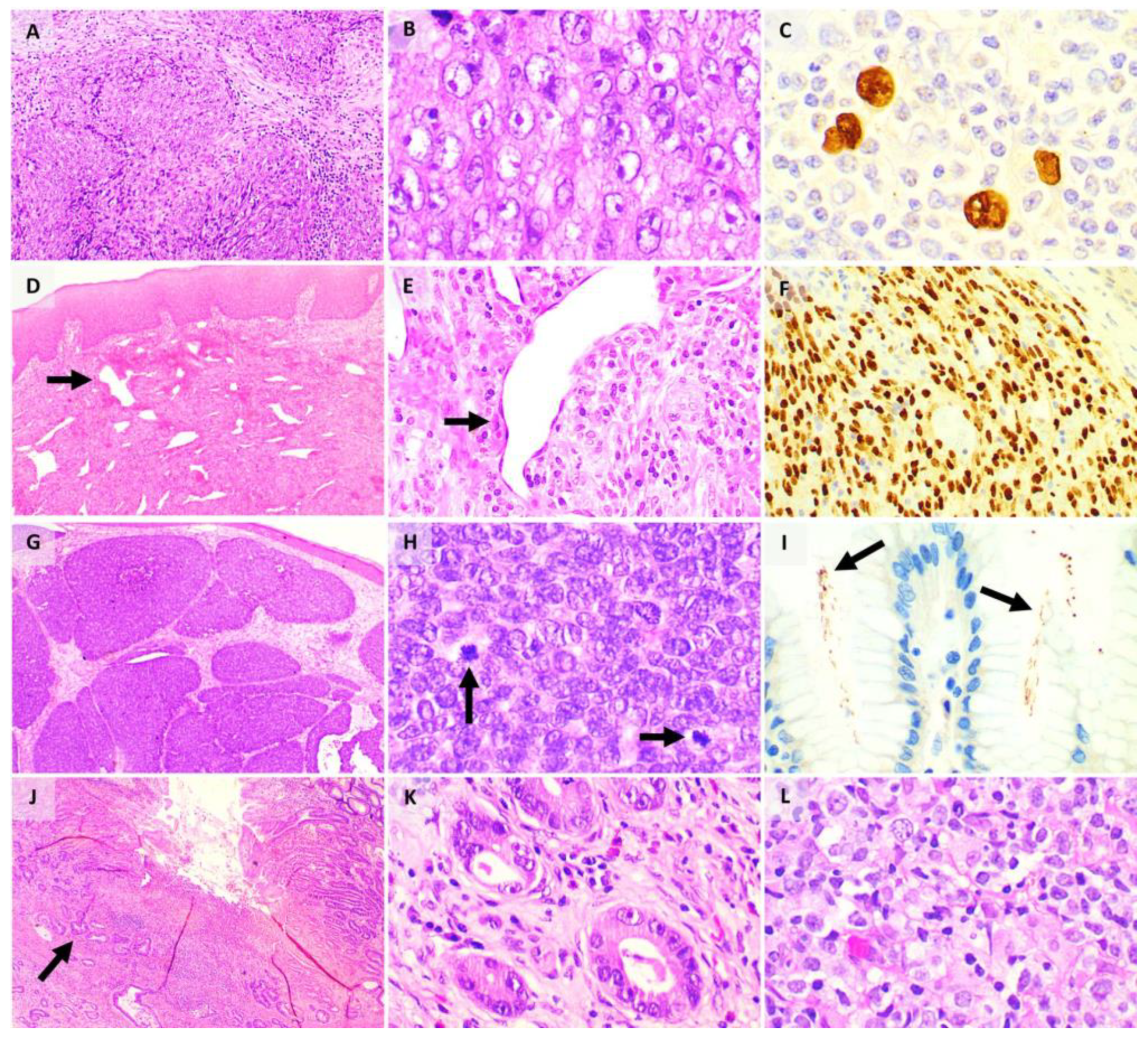{kind=link}
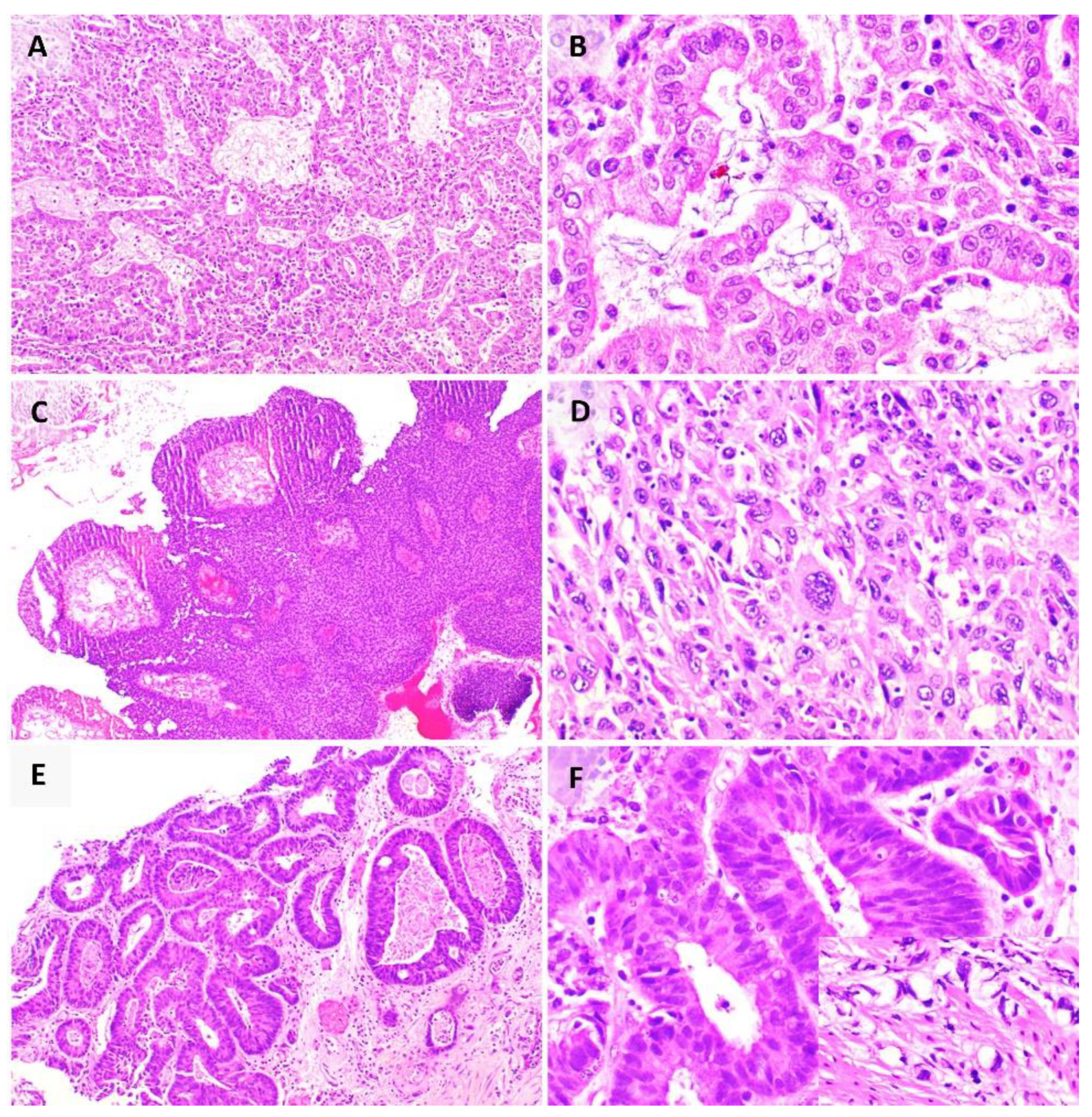{kind=link}
Abstract
:1. Introduction
2. Benign Mimickers of Malignancy
2.1. Herpes Simplex Virus (HSV)
2.2. Cytomegalovirus (CMV)
2.3. BK Polyomavirus (BKPyV)
2.4. Adenovirus, Measles Virus, and Respiratory Syncytial Virus (RSV)
2.5. Granulomatous Inflammation
3. Oncogenic Pathogens
3.1. Hepatitis B and C Viruses (HBV, HCV)
3.2. Human Papillomavirus (HPV)
3.3. Epstein–Barr Virus (EBV)
3.4. Human Herpes Virus 8 (HHV8)
3.5. Human T-Cell Leukemia Virus Type 1 (HTLV-1)
3.6. Merkel Cell Polyomavirus (MCPyV)
3.7. Helicobacter Pylori (HP)
3.8. Opisthorcis Viverrine (Ov) and Clonorchis Sinensis (Cs)
3.9. Schistosoma Haematobium (Sh)
3.10. COVID-19
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Danaei, G.; Vander Hoorn, S.; Lopez, A.D.; Murray, C.J.; Ezzati, M. Comparative Risk Assessment collaborating, g. Causes of cancer in the world: Comparative risk assessment of nine behavioural and environmental risk factors. Lancet 2005, 366, 1784–1793. [Google Scholar] [CrossRef] [Green Version]
- De Flora, S.; La Maestra, S. Epidemiology of cancers of infectious origin and prevention strategies. J. Prev. Med. Hyg. 2015, 56, E15–E20. [Google Scholar]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagano, J.S.; Blaser, M.; Buendia, M.A.; Damania, B.; Khalili, K.; Raab-Traub, N.; Roizman, B. Infectious agents and cancer: Criteria for a causal relation. Semin. Cancer Biol. 2004, 14, 453–471. [Google Scholar] [CrossRef] [PubMed]
- Kuper, H.; Adami, H.O.; Trichopoulos, D. Infections as a major preventable cause of human cancer. J. Intern. Med. 2000, 248, 171–183. [Google Scholar] [CrossRef]
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.X.T.; Han, M.; Ki, M.; Kim, Y.A.; Oh, J.K. The Economic Burden of Cancers Attributable to Infection in the Republic of Korea: A Prevalence-Based Study. Int. J. Environ. Res. Public Health 2020, 17, 7592. [Google Scholar] [CrossRef]
- Cancer IAfRo. IARC Monographs on the Identification of Carcinogenic Hazards to Humans. Available online: https://monographs.iarc.who.int/list-of-classifications/ (accessed on 9 August 2021).
- Cancer IAfRo. Cancers Attributbable to Infections: Data Sources and Methods. Available online: https://gco.iarc.fr/causes/infections/data-sources-methods (accessed on 22 August 2021).
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [Green Version]
- Velavan, T.P.; Meyer, C.G. The COVID-19 epidemic. Trop. Med. Int. Health 2020, 25, 278–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tutuncuoglu, B.; Cakir, M.; Batra, J.; Bouhaddou, M.; Eckhardt, M.; Gordon, D.E.; Krogan, N.J. The Landscape of Human Cancer Proteins Targeted by SARS-CoV-2. Cancer Discov. 2020, 10, 916–921. [Google Scholar] [CrossRef]
- Zong, Z.; Wei, Y.; Ren, J.; Zhang, L.; Zhou, F. The intersection of COVID-19 and cancer: Signaling pathways and treatment implications. Mol. Cancer 2021, 20, 76. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dai, L.; Barrett, L.; Post, S.R.; Qin, Z. SARS-CoV-2 proteins and anti-COVID-19 drugs induce lytic reactivation of an oncogenic virus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Baldassarri, R.J.; Kumar, D.; Baldassarri, S.; Cai, G. Diagnosis of Infectious Diseases in the Lower Respiratory Tract: A Cytopathologist’s Perspective. Arch. Pathol. Lab. Med. 2019, 143, 683–694. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Giron, R.; Pantanowitz, L. Lower respiratory tract viral infections: Diagnostic role of exfoliative cytology. Diagn. Cytopathol. 2017, 45, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Kumar Vinay, A.A.; Aster, J. Robbins Cotrain Pathologic Basis of Disease, 10th ed.; Elsevier: Philadelphia, PA, USA, 2020. [Google Scholar]
- Whitley, R.J.; Roizman, B. Herpes simplex virus infections. Lancet 2001, 357, 1513–1518. [Google Scholar] [CrossRef]
- Naib, Z.M.; Nahmias, A.J.; Josey, W.E. Cytology and histopathology of cervical herpes simplex infection. Cancer 1966, 19, 1026–1031. [Google Scholar] [CrossRef]
- Lapkus, O.; Elsheikh, T.M.; Ujevich, B.A.; Liu, Y.L.; Silverman, J.F. Pitfalls in the diagnosis of herpes simplex infection in respiratory cytology. Acta Cytol. 2006, 50, 617–620. [Google Scholar] [CrossRef]
- Blank, H.; Burgoon, C.F.; Baldridge, G.D.; Mc, C.P.; Urbach, F. Cytologic smears in diagnosis of herpes simplex, herpes zoster, and varicella. J. Am. Med. Assoc. 1951, 146, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.M.; Sharma, B.K.; Singh, V.; Luthra, U.K. Immunocytological demonstration of HSV-II antigen on exfoliated cells from precancerous and cancerous lesions of the uterine cervix. Diagn Cytopathol. 1988, 4, 48–49. [Google Scholar] [CrossRef] [PubMed]
- Solomon, I.H.; Hornick, J.L.; Laga, A.C. Immunohistochemistry Is Rarely Justified for the Diagnosis of Viral Infections. Am. J. Clin. Pathol. 2017, 147, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.H.; Jiang, H.; Lee, M.; Liu, F.; Zhou, Z.H. Three-dimensional visualization of tegument/capsid interactions in the intact human cytomegalovirus. Virology 1999, 260, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, M.K.; Khanna, R. Human cytomegalovirus: Clinical aspects, immune regulation, and emerging treatments. Lancet Infect. Dis 2004, 4, 725–738. [Google Scholar] [CrossRef]
- Taylor, G.H. Cytomegalovirus. Am. Fam. Physician 2003, 67, 519–524. [Google Scholar] [PubMed]
- Dioverti, M.V.; Razonable, R.R. Cytomegalovirus. Microbiol. Spectr. 2016, 4, 151–171. [Google Scholar] [CrossRef]
- Griffiths, P.; Baraniak, I.; Reeves, M. The pathogenesis of human cytomegalovirus. J. Pathol. 2015, 235, 288–297. [Google Scholar] [CrossRef]
- Elgert, P.A.; Yee-Chang, M.; Simsir, A. Cytomegalovirus (CMV) in cervical cancer screening tests: A series of 8 cases and review of the literature. Diagn. Cytopathol. 2018, 46, 593–599. [Google Scholar] [CrossRef]
- Weldon-Linne, C.M.; Rhone, D.P.; Bourassa, R. Bronchoscopy specimens in adults with AIDS. Comparative yields of cytology, histology and culture for diagnosis of infectious agents. Chest 1990, 98, 24–28. [Google Scholar] [CrossRef]
- Woods, G.L.; Thompson, A.B.; Rennard, S.L.; Linder, J. Detection of cytomegalovirus in bronchoalveolar lavage specimens. Spin amplification and staining with a monoclonal antibody to the early nuclear antigen for diagnosis of cytomegalovirus pneumonia. Chest 1990, 98, 568–575. [Google Scholar] [CrossRef]
- Weiss, R.L.; Snow, G.W.; Schumann, G.B.; Hammond, M.E. Diagnosis of cytomegalovirus pneumonitis on bronchoalveolar lavage fluid: Comparison of cytology, immunofluorescence, and in situ hybridization with viral isolation. Diagn Cytopathol. 1991, 7, 243–247. [Google Scholar] [CrossRef]
- Behrens, H.W.; Quick, C.A. Bronchoscopic diagnosis of cytomegalovirus infection. J. Infect. Dis. 1974, 130, 174–176. [Google Scholar] [CrossRef]
- Delvenne, P.; Arrese, J.E.; Thiry, A.; Borlee-Hermans, G.; Pierard, G.E.; Boniver, J. Detection of cytomegalovirus, Pneumocystis carinii, and aspergillus species in bronchoalveolar lavage fluid. A comparison of techniques. Am. J. Clin. Pathol. 1993, 100, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.Y.; An, J.; Ha, S.Y.; Chung, D.H.; Lee, S.; Kim, H. Morphologic Analysis of Cytomegalovirus Infected Cells in Bronchial Washing Cytology: Comparison of Liquid-Based Preparation and Conventional Smear. J. Pathol. Transl. Med. 2016, 50, 147–154. [Google Scholar] [CrossRef]
- Solans, E.P.; Yong, S.; Husain, A.N.; Eichorst, M.; Gattuso, P. Bronchioloalveolar lavage in the diagnosis of CMV pneumonitis in lung transplant recipients: An immunocytochemical study. Diagn. Cytopathol. 1997, 16, 350–352. [Google Scholar] [CrossRef]
- Siguier, M.; Sellier, P.; Bergmann, J.F. BK-virus infections: A literature review. Med. Mal. Infect. 2012, 42, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Alexiev, B.A.; Randhawa, P.; Vazquez Martul, E.; Zeng, G.; Luo, C.; Ramos, E.; Drachenberg, C.B.; Papadimitriou, J.C. BK virus-associated urinary bladder carcinoma in transplant recipients: Report of 2 cases, review of the literature, and proposed pathogenetic model. Hum. Pathol. 2013, 44, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, H.H. BK virus: Opportunity makes a pathogen. Clin. Infect. Dis. 2005, 41, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Reploeg, M.D.; Storch, G.A.; Clifford, D.B. Bk virus: A clinical review. Clin. Infect. Dis. 2001, 33, 191–202. [Google Scholar] [CrossRef]
- Goudsmit, J.; Wertheim-van Dillen, P.; van Strien, A.; van der Noordaa, J. The role of BK virus in acute respiratory tract disease and the presence of BKV DNA in tonsils. J. Med. Virol. 1982, 10, 91–99. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Randhawa, P.S.; Practice, A.S.T.I.D.C.o. BK polyomavirus in solid organ transplantation-Guidelines from the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transplant. 2019, 33, e13528. [Google Scholar] [CrossRef]
- Geetha, V.; Rao, L.; Monappa, V.; Susmitha, M.; Prabhu, R. Decoy cells in urine cytology: A useful clue to post-transplant polyoma virus infection. J. Cytol. 2012, 29, 133–134. [Google Scholar] [CrossRef] [PubMed]
- Sawinski, D.; Goral, S. BK virus infection: An update on diagnosis and treatment. Nephrol. Dial. Transplant. 2015, 30, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Hogan, T.F.; Padgett, B.L.; Walker, D.L.; Borden, E.C.; McBain, J.A. Rapid detection and identification of JC virus and BK virus in human urine by using immunofluorescence microscopy. J. Clin. Microbiol. 1980, 11, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.L.; Manfrinatto, J.A.; Cia, E.M.; Carvalho, R.B.; Quadros, K.R.; Alves-Filho, G.; Mazzali, M. Urine cytology as a screening method for polyoma virus active infection. Transplant. Proc. 2004, 36, 899–901. [Google Scholar] [CrossRef]
- Drachenberg, R.C.; Drachenberg, C.B.; Papadimitriou, J.C.; Ramos, E.; Fink, J.C.; Wali, R.; Weir, M.R.; Cangro, C.B.; Klassen, D.K.; Khaled, A.; et al. Morphological spectrum of polyoma virus disease in renal allografts: Diagnostic accuracy of urine cytology. Am. J. Transplant. 2001, 1, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Bulut, Y.; Ozdemir, E.; Ozercan, H.I.; Etem, E.O.; Aker, F.; Toraman, Z.A.; Seyrek, A.; Firdolas, F. Potential relationship between BK virus and renal cell carcinoma. J. Med. Virol. 2013, 85, 1085–1089. [Google Scholar] [CrossRef]
- Weinreb, D.B.; Desman, G.T.; Amolat-Apiado, M.J.; Burstein, D.E.; Godbold, J.H., Jr.; Johnson, E.M. Polyoma virus infection is a prominent risk factor for bladder carcinoma in immunocompetent individuals. Diagn. Cytopathol. 2006, 34, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Wojno, K.; Imperiale, M.J. BK virus as a cofactor in the etiology of prostate cancer in its early stages. J. Virol. 2008, 82, 2705–2714. [Google Scholar] [CrossRef] [Green Version]
- Das, D.; Shah, R.B.; Imperiale, M.J. Detection and expression of human BK virus sequences in neoplastic prostate tissues. Oncogene 2004, 23, 7031–7046. [Google Scholar] [CrossRef] [Green Version]
- Rollison, D.E.; Sexton, W.J.; Rodriguez, A.R.; Kang, L.C.; Daniel, R.; Shah, K.V. Lack of BK virus DNA sequences in most transitional-cell carcinomas of the bladder. Int. J. Cancer 2007, 120, 1248–1251. [Google Scholar] [CrossRef]
- Kumari, K.; Pradeep, I.; Kakkar, A.; Dinda, A.K.; Seth, A.; Nayak, B.; Singh, G. BK polyomavirus and urothelial carcinoma: Experience at a tertiary care centre in India with review of literature. Ann. Diagn Pathol. 2019, 40, 77–80. [Google Scholar] [CrossRef]
- Lenaerts, L.; De Clercq, E.; Naesens, L. Clinical features and treatment of adenovirus infections. Rev. Med. Virol. 2008, 18, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Kojaoghlanian, T.; Flomenberg, P.; Horwitz, M.S. The impact of adenovirus infection on the immunocompromised host. Rev. Med. Virol. 2003, 13, 155–171. [Google Scholar] [CrossRef]
- Avellon, A.; Perez, P.; Aguilar, J.C.; Lejarazu, R.; Echevarria, J.E. Rapid and sensitive diagnosis of human adenovirus infections by a generic polymerase chain reaction. J. Virol. Methods 2001, 92, 113–120. [Google Scholar] [CrossRef]
- Matthes-Martin, S.; Boztug, H.; Lion, T. Diagnosis and treatment of adenovirus infection in immunocompromised patients. Expert Rev. Anti Infect. Ther. 2013, 11, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Harbhajanka, A.; Michael, C.W. Adenoviral cytopathic changes in pleural fluid: A unique cytological presentation mimicking malignancy. Diagn. Cytopathol. 2021, 49, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Zaharopoulos, P. Viral cytopathic changes in urine cytology of ileal conduit attributed to adenovirus: Report of a case. Diagn. Cytopathol. 2004, 30, 284–287. [Google Scholar] [CrossRef]
- Moss, W.J.; Griffin, D.E. Measles. Lancet 2012, 379, 153–164. [Google Scholar] [CrossRef]
- Katz, M. Clinical spectrum of measles. Curr. Top. Microbiol. Immunol. 1995, 191, 1–12. [Google Scholar] [CrossRef]
- Robbins, F.C. Measles: Clinical features. Pathogenesis, pathology and complications. Am. J. Dis Child. 1962, 103, 266–273. [Google Scholar] [CrossRef]
- Bester, J.C. Measles and Measles Vaccination: A Review. JAMA Pediatr. 2016, 170, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Bolande, R.P. Significance and nature of inclusion-bearing cells in the urine of patients with measles. N. Engl. J. Med. 1961, 265, 919–923. [Google Scholar] [CrossRef]
- Lipsey, A.I.; Bolande, R.P. The exfoliative source of abnormal cells in urine sediment of patients with measles. Am. J. Dis. Child. 1967, 113, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Simoes, E.A. Respiratory syncytial virus infection. Lancet 1999, 354, 847–852. [Google Scholar] [CrossRef]
- Hall, C.B.; Simoes, E.A.; Anderson, L.J. Clinical and epidemiologic features of respiratory syncytial virus. Curr. Top. Microbiol. Immunol. 2013, 372, 39–57. [Google Scholar] [CrossRef]
- Hall, C.B. Respiratory syncytial virus and parainfluenza virus. N. Engl. J. Med. 2001, 344, 1917–1928. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.K.; Pritt, B.S.; Alexander, M.P. Histopathologic review of granulomatous inflammation. J. Clin. Tuberc. Other Mycobact. Dis. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Chopra, A.; Avadhani, V.; Tiwari, A.; Riemer, E.C.; Sica, G.; Judson, M.A. Granulomatous lung disease: Clinical aspects. Expert. Rev. Respir. Med. 2020, 14, 1045–1063. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.T.; Williams, W.J. Granulomatous inflammation--a review. J. Clin. Pathol. 1983, 36, 723–733. [Google Scholar] [CrossRef]
- Mehrotra, R.; Dhingra, V. Cytological diagnosis of sarcoidosis revisited: A state of the art review. Diagn. Cytopathol. 2011, 39, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, N.N.; Leighton, J.A.; Heigh, R.I.; Shapiro, M.S. Clinical significance of granuloma in Crohn’s disease. Inflamm. Bowel. Dis. 2002, 8, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Brincker, H. Sarcoid reactions in malignant tumours. Cancer Treat. Rev. 1986, 13, 147–156. [Google Scholar] [CrossRef]
- Tjan-Heijnen, V.C.; Vlasveld, L.T.; Pernet, F.P.; Pauwels, P.; De Mulder, P.H. Coincidence of seminoma and sarcoidosis: A myth or fact? Ann. Oncol. 1998, 9, 321–325. [Google Scholar] [CrossRef]
- Khurana, K.K.; Stanley, M.W.; Powers, C.N.; Pitman, M.B. Aspiration cytology of malignant neoplasms associated with granulomas and granuloma-like features: Diagnostic dilemmas. Cancer 1998, 84, 84–91. [Google Scholar] [CrossRef]
- Kennedy, M.P.; Jimenez, C.A.; Mhatre, A.D.; Morice, R.C.; Eapen, G.A. Clinical implications of granulomatous inflammation detected by endobronchial ultrasound transbronchial needle aspiration in patients with suspected cancer recurrence in the mediastinum. J. Cardiothorac. Surg. 2008, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Betz, S.A.; See, W.A.; Cohen, M.B. Granulomatous inflammation in bladder wash specimens after intravesical bacillus Calmette-Guerin therapy for transitional cell carcinoma of the bladder. Am. J. Clin. Pathol. 1993, 99, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Ocal, K.; Dag, A.; Turkmenoglu, O.; Kara, T.; Seyit, H.; Konca, K. Granulomatous mastitis: Clinical, pathological features, and management. Breast J. 2010, 16, 176–182. [Google Scholar] [CrossRef]
- Chandanwale, S.; Naragude, P.; Shetty, A.; Sawadkar, M.; Raj, A.; Bhide, A.; Singh, M. Cytomorphological Spectrum of Granulomatous Mastitis: A Study of 33 Cases. Eur. J. Breast Health 2020, 16, 146–151. [Google Scholar] [CrossRef]
- Erhan, Y.; Veral, A.; Kara, E.; Ozdemir, N.; Kapkac, M.; Ozdedeli, E.; Yilmaz, R.; Koyuncu, A.; Erhan, Y.; Ozbal, O. A clinicopthologic study of a rare clinical entity mimicking breast carcinoma: Idiopathic granulomatous mastitis. Breast 2000, 9, 52–56. [Google Scholar] [CrossRef]
- Heer, R.; Shrimankar, J.; Griffith, C.D. Granulomatous mastitis can mimic breast cancer on clinical, radiological or cytological examination: A cautionary tale. Breast 2003, 12, 283–286. [Google Scholar] [CrossRef]
- Mitra, S.; Dey, P. Fine-needle aspiration and core biopsy in the diagnosis of breast lesions: A comparison and review of the literature. Cytojournal 2016, 13, 18. [Google Scholar] [CrossRef]
- Fletcher, G.J.; Eapen, C.E.; Abraham, P. Hepatitis B genotyping: The utility for the clinicians. Indian J. Gastroenterol. 2020, 39, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Fiel, M.I. Pathology of chronic hepatitis B and chronic hepatitis C. Clin. Liver Dis. 2010, 14, 555–575. [Google Scholar] [CrossRef] [PubMed]
- WHO Classification of Tumours Editorial Board. WHO Classification of Tumours: Digestive System Tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2019. [Google Scholar]
- Bruix, J.; Sherman, M.; American Association for the Study of Liver. Management of hepatocellular carcinoma: An update. Hepatology 2011, 53, 1020–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.D.; Sun, H.C. Emerging agents and regimens for hepatocellular carcinoma. J. Hematol. Oncol. 2019, 12, 110. [Google Scholar] [CrossRef]
- Longo, V.; Brunetti, O.; Gnoni, A.; Licchetta, A.; Delcuratolo, S.; Memeo, R.; Solimando, A.G.; Argentiero, A. Emerging role of Immune Checkpoint Inhibitors in Hepatocellular Carcinoma. Medicina 2019, 55, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Fang, Q.X.; Chen, D.B.; Chen, H.S. Neoantigen vaccine: An emerging immunotherapy for hepatocellular carcinoma. World J. Gastrointest. Oncol. 2021, 13, 673–683. [Google Scholar] [CrossRef]
- Asiaf, A.; Ahmad, S.T.; Mohammad, S.O.; Zargar, M.A. Review of the current knowledge on the epidemiology, pathogenesis, and prevention of human papillomavirus infection. Eur. J. Cancer Prev. 2014, 23, 206–224. [Google Scholar] [CrossRef] [PubMed]
- Lekoane, K.M.B.; Kuupiel, D.; Mashamba-Thompson, T.P.; Ginindza, T.G. The interplay of HIV and human papillomavirus-related cancers in sub-Saharan Africa: Scoping review. Syst. Rev. 2020, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, L.M. Pathobiology of Human Disease: A Dynamic Encyclopedia of Disease Mechanisms; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- WHO Classification of Tumours Editorial Board. WHO Classification of Tumours: Female Genital Tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Thompson, L.D.R. HPV-Related Multiphenotypic Sinonasal Carcinoma. Ear. Nose Throat J. 2020, 99, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Kesic, V. Management of cervical cancer. Eur. J. Surg. Oncol. 2006, 32, 832–837. [Google Scholar] [CrossRef]
- Fakhr, E.; Modic, Z.; Cid-Arregui, A. Recent developments in immunotherapy of cancers caused by human papillomaviruses. Immunology 2021, 163, 33–45. [Google Scholar] [CrossRef]
- Skeate, J.G.; Woodham, A.W.; Einstein, M.H.; Da Silva, D.M.; Kast, W.M. Current therapeutic vaccination and immunotherapy strategies for HPV-related diseases. Hum. Vaccin Immunother. 2016, 12, 1418–1429. [Google Scholar] [CrossRef]
- Luvero, D.; Lopez, S.; Bogani, G.; Raspagliesi, F.; Angioli, R. From the Infection to the Immunotherapy in Cervical Cancer: Can We Stop the Natural Course of the Disease? Vaccines 2020, 8, 597. [Google Scholar] [CrossRef]
- Shannon-Lowe, C.; Rickinson, A. The Global Landscape of EBV-Associated Tumors. Front. Oncol. 2019, 9, 713. [Google Scholar] [CrossRef] [Green Version]
- Louissaint, A., Jr.; Ferry, J.A.; Soupir, C.P.; Hasserjian, R.P.; Harris, N.L.; Zukerberg, L.R. Infectious mononucleosis mimicking lymphoma: Distinguishing morphological and immunophenotypic features. Mod. Pathol. 2012, 25, 1149–1159. [Google Scholar] [CrossRef] [Green Version]
- van Zyl, D.G.; Mautner, J.; Delecluse, H.J. Progress in EBV Vaccines. Front. Oncol. 2019, 9, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke-Serinsoz, E.; Fend, F.; Quintanilla-Martinez, L. Human immunodeficiency virus (HIV) and Epstein-Barr virus (EBV) related lymphomas, pathology view point. Semin. Diagn. Pathol. 2017, 34, 352–363. [Google Scholar] [CrossRef]
- Hong, M.; Tang, K.; Qian, J.; Deng, H.; Zeng, M.; Zheng, S.; Ding, K.; Du, Y.; Sun, R. Immunotherapy for EBV-Associated Nasopharyngeal Carcinoma. Crit. Rev. Oncog. 2018, 23, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Sarid, R.; Olsen, S.J.; Moore, P.S. Kaposi′s sarcoma-associated herpesvirus: Epidemiology, virology, and molecular biology. Adv. Virus Res. 1999, 52, 139–232. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Arber, D.A.; Hasserjian, R.P.; Le Beau, M.M.; et al. WHO Classification of Tumours of Haematopoeitic and Lymphoid Tissues, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2017. [Google Scholar]
- Sarid, R.; Klepfish, A.; Schattner, A. Virology, pathogenetic mechanisms, and associated diseases of Kaposi sarcoma-associated herpesvirus (human herpesvirus 8). Mayo Clin. Proc. 2002, 77, 941–949. [Google Scholar] [CrossRef] [Green Version]
- Vangipuram, R.; Tyring, S.K. Epidemiology of Kaposi sarcoma: Review and description of the nonepidemic variant. Int. J. Dermatol. 2019, 58, 538–542. [Google Scholar] [CrossRef]
- Schulz, T.F.; Cesarman, E. Kaposi Sarcoma-associated Herpesvirus: Mechanisms of oncogenesis. Curr. Opin. Virol. 2015, 14, 116–128. [Google Scholar] [CrossRef]
- Schulz, T.F. Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8): Epidemiology and pathogenesis. J. Antimicrob. Chemother. 2000, 45, 15–27. [Google Scholar] [CrossRef]
- Bhaduri-McIntosh, S. Human herpesvirus-8: Clinical features of an emerging viral pathogen. Pediatr. Infect. Dis. J. 2005, 24, 81–82. [Google Scholar] [CrossRef]
- Geraminejad, P.; Memar, O.; Aronson, I.; Rady, P.L.; Hengge, U.; Tyring, S.K. Kaposi’s sarcoma and other manifestations of human herpesvirus 8. J. Am. Acad. Dermatol. 2002, 47, 641–655. [Google Scholar] [CrossRef]
- Ablashi, D.V.; Chatlynne, L.G.; Whitman, J.E., Jr.; Cesarman, E. Spectrum of Kaposi’s sarcoma-associated herpesvirus, or human herpesvirus 8, diseases. Clin. Microbiol. Rev. 2002, 15, 439–464. [Google Scholar] [CrossRef] [Green Version]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Dittmer, D.P.; Damania, B. Kaposi sarcoma-associated herpesvirus: Immunobiology, oncogenesis, and therapy. J. Clin. Investig. 2016, 126, 3165–3175. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, L.D. Human herpesvirus-8: Kaposi sarcoma, multicentric Castleman disease, and primary effusion lymphoma. Hematol. Am. Soc. Hematol. Educ. Program. 2013, 2013, 103–108. [Google Scholar] [CrossRef]
- WHO Classification of Tumours Editorial Board. WHO Classification of Tumours: Soft Tissue and Bone tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Cheuk, W.; Wong, K.O.; Wong, C.S.; Dinkel, J.E.; Ben-Dor, D.; Chan, J.K. Immunostaining for human herpesvirus 8 latent nuclear antigen-1 helps distinguish Kaposi sarcoma from its mimickers. Am. J. Clin. Pathol. 2004, 121, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Robin, Y.M.; Guillou, L.; Michels, J.J.; Coindre, J.M. Human herpesvirus 8 immunostaining: A sensitive and specific method for diagnosing Kaposi sarcoma in paraffin-embedded sections. Am. J. Clin. Pathol. 2004, 121, 330–334. [Google Scholar] [CrossRef]
- Du, M.Q.; Bacon, C.M.; Isaacson, P.G. Kaposi sarcoma-associated herpesvirus/human herpesvirus 8 and lymphoproliferative disorders. J. Clin. Pathol. 2007, 60, 1350–1357. [Google Scholar] [CrossRef] [Green Version]
- Gantt, S.; Casper, C. Human herpesvirus 8-associated neoplasms: The roles of viral replication and antiviral treatment. Curr. Opin. Infect. Dis. 2011, 24, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Broussard, G.; Damania, B. KSHV: Immune Modulation and Immunotherapy. Front. Immunol. 2019, 10, 3084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambichler, T.; Koim, S.; Wrobel, M.; Kafferlein, H.U.; Bruning, T.; Stockfleth, E.; Becker, J.C.; Lang, K. Expression of Programmed Cell Death Proteins in Kaposi Sarcoma and Cutaneous Angiosarcoma. J. Immunother. 2020, 43, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Edlich, R.F.; Arnette, J.A.; Williams, F.M. Global epidemic of human T-cell lymphotropic virus type-I (HTLV-I). J. Emerg. Med. 2000, 18, 109–119. [Google Scholar] [CrossRef]
- Manns, A.; Hisada, M.; La Grenade, L. Human T-lymphotropic virus type I infection. Lancet 1999, 353, 1951–1958. [Google Scholar] [CrossRef]
- Hlela, C.; Shepperd, S.; Khumalo, N.P.; Taylor, G.P. The prevalence of human T-cell lymphotropic virus type 1 in the general population is unknown. AIDS Rev. 2009, 11, 205–214. [Google Scholar]
- Ahmadi Ghezeldasht, S.; Shirdel, A.; Assarehzadegan, M.A.; Hassannia, T.; Rahimi, H.; Miri, R.; Rezaee, S.A. Human T Lymphotropic Virus Type I (HTLV-I) Oncogenesis: Molecular Aspects of Virus and Host Interactions in Pathogenesis of Adult T cell Leukemia/Lymphoma (ATL). Iran. J. Basic Med. Sci. 2013, 16, 179–195. [Google Scholar] [PubMed]
- Bangham, C.R.M.; Matsuoka, M. Human T-cell leukaemia virus type 1: Parasitism and pathogenesis. Philos Trans. R Soc. Lond. B Biol. Sci. 2017, 372, 20160272. [Google Scholar] [CrossRef] [Green Version]
- Tagaya, Y.; Matsuoka, M.; Gallo, R. 40 years of the human T-cell leukemia virus: Past, present, and future. F1000 Res. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.L.; Wei, J.Y.; Wang, L.; Huang, S.L.; Chen, J.L. Human T-cell lymphotropic virus type 1 and its oncogenesis. Acta Pharmacol. Sin. 2017, 38, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Tsukasaki, K.; Tobinai, K. Human T-cell lymphotropic virus type I-associated adult T-cell leukemia-lymphoma: New directions in clinical research. Clin. Cancer Res. 2014, 20, 5217–5225. [Google Scholar] [CrossRef] [Green Version]
- Hermine, O.; Ramos, J.C.; Tobinai, K. A Review of New Findings in Adult T-cell Leukemia-Lymphoma: A Focus on Current and Emerging Treatment Strategies. Adv. Ther. 2018, 35, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Reljic, T.; Klocksieben, F.; Sher, T.; Ayala, E.; Murthy, H.; Bazarbachi, A.; Kumar, A.; Kharfan-Dabaja, M.A. Efficacy of Allogeneic Hematopoietic Cell Transplantation in Human T Cell Lymphotropic Virus Type 1-Associated Adult T Cell Leukemia/Lymphoma: Results of a Systematic Review/Meta-Analysis. Biol. Blood Marrow Transplant. 2019, 25, 1695–1700. [Google Scholar] [CrossRef]
- Spurgeon, M.E.; Lambert, P.F. Merkel cell polyomavirus: A newly discovered human virus with oncogenic potential. Virology 2013, 435, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.C.; Stang, A.; DeCaprio, J.A.; Cerroni, L.; Lebbe, C.; Veness, M.; Nghiem, P. Merkel cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17077. [Google Scholar] [CrossRef]
- Liu, W.; MacDonald, M.; You, J. Merkel cell polyomavirus infection and Merkel cell carcinoma. Curr. Opin. Virol. 2016, 20, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Takeuchi, Y.; Takenouchi, T.; Hirofuji, A.; Tsuchida, T.; Kabumoto, T.; Fujiwara, H.; Ito, M.; Shimizu, A.; Okada, E.; et al. The prevalence of Merkel cell polyomavirus in Japanese patients with Merkel cell carcinoma. J. Dermatol. Sci. 2013, 70, 99–107. [Google Scholar] [CrossRef]
- Duncavage, E.J.; Zehnbauer, B.A.; Pfeifer, J.D. Prevalence of Merkel cell polyomavirus in Merkel cell carcinoma. Mod. Pathol. 2009, 22, 516–521. [Google Scholar] [CrossRef]
- Mangana, J.; Dziunycz, P.; Kerl, K.; Dummer, R.; Cozzio, A. Prevalence of Merkel cell polyomavirus among Swiss Merkel cell carcinoma patients. Dermatology 2010, 221, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Garneski, K.M.; Warcola, A.H.; Feng, Q.; Kiviat, N.B.; Leonard, J.H.; Nghiem, P. Merkel cell polyomavirus is more frequently present in North American than Australian Merkel cell carcinoma tumors. J. Investig. Dermatol. 2009, 129, 246–248. [Google Scholar] [CrossRef] [Green Version]
- Wieland, U.; Kreuter, A. Merkel cell polyomavirus infection and Merkel cell carcinoma in HIV-positive individuals. Curr. Opin. Oncol. 2011, 23, 488–493. [Google Scholar] [CrossRef]
- Santos-Juanes, J.; Fernandez-Vega, I.; Fuentes, N.; Galache, C.; Coto-Segura, P.; Vivanco, B.; Astudillo, A.; Martinez-Camblor, P. Merkel cell carcinoma and Merkel cell polyomavirus: A systematic review and meta-analysis. Br. J. Dermatol. 2015, 173, 42–49. [Google Scholar] [CrossRef]
- Pietropaolo, V.; Prezioso, C.; Moens, U. Merkel Cell Polyomavirus and Merkel Cell Carcinoma. Cancers 2020, 12, 1774. [Google Scholar] [CrossRef]
- Grundhoff, A.; Fischer, N. Merkel cell polyomavirus, a highly prevalent virus with tumorigenic potential. Curr. Opin. Virol. 2015, 14, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Elder, D.E.; Scholyer, R.A.; Willemze, R. WHO Classification of Skin Tumours, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2018. [Google Scholar]
- Leech, S.N.; Kolar, A.J.; Barrett, P.D.; Sinclair, S.A.; Leonard, N. Merkel cell carcinoma can be distinguished from metastatic small cell carcinoma using antibodies to cytokeratin 20 and thyroid transcription factor 1. J. Clin. Pathol. 2001, 54, 727–729. [Google Scholar] [CrossRef]
- Tabachnick-Cherny, S.; Pulliam, T.; Church, C.; Koelle, D.M.; Nghiem, P. Polyomavirus-driven Merkel cell carcinoma: Prospects for therapeutic vaccine development. Mol. Carcinog. 2020, 59, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe, S.E. Helicobacter pylori Infection. N. Engl. J. Med. 2019, 380, 1158–1165. [Google Scholar] [CrossRef]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [Green Version]
- Katoh, H.; Ishikawa, S. Lifestyles, genetics, and future perspectives on gastric cancer in east Asian populations. J. Hum. Genet. 2021, 66, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Lopez, A.; Shanbhag, N.; Badgwell, B.; Baba, H.; Ajani, J. Recent advances in the management of gastric adenocarcinoma patients. F1000 Res. 2018, 7. [Google Scholar] [CrossRef]
- Silva, R.; Gullo, I.; Carneiro, F. The PD-1:PD-L1 immune inhibitory checkpoint in Helicobacter pylori infection and gastric cancer: A comprehensive review and future perspectives. Porto Biomed. J. 2016, 1, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Oster, P.; Vaillant, L.; Riva, E.; McMillan, B.; Begka, C.; Truntzer, C.; Richard, C.; Leblond, M.M.; Messaoudene, M.; Machremi, E.; et al. Helicobacter pylori infection has a detrimental impact on the efficacy of cancer immunotherapies. Gut 2021. [Google Scholar] [CrossRef]
- Hong, S.T.; Fang, Y. Clonorchis sinensis and clonorchiasis, an update. Parasitol. Int. 2012, 61, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kaewpitoon, N.; Kaewpitoon, S.J.; Pengsaa, P.; Sripa, B. Opisthorchis viverrini: The carcinogenic human liver fluke. World J. Gastroenterol. 2008, 14, 666–674. [Google Scholar] [CrossRef]
- Sithithaworn, P.; Haswell-Elkins, M. Epidemiology of Opisthorchis viverrini. Acta Trop. 2003, 88, 187–194. [Google Scholar] [CrossRef]
- Smout, M.J.; Sripa, B.; Laha, T.; Mulvenna, J.; Gasser, R.B.; Young, N.D.; Bethony, J.M.; Brindley, P.J.; Loukas, A. Infection with the carcinogenic human liver fluke, Opisthorchis viverrini. Mol. Biosyst. 2011, 7, 1367–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.L.; Huang, Y.; Yu, X.B. Current status and perspectives of Clonorchis sinensis and clonorchiasis: Epidemiology, pathogenesis, omics, prevention and control. Infect. Dis. Poverty 2016, 5, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. DPDx—Laboatory Identification of Parasites of Public Health Concern: Opisthorchiasis. Available online: https://www.cdc.gov/dpdx/opisthorchiasis/index.html (accessed on 24 August 2021).
- Kim, T.S.; Pak, J.H.; Kim, J.B.; Bahk, Y.Y. Clonorchis sinensis, an oriental liver fluke, as a human biological agent of cholangiocarcinoma: A brief review. BMB Rep. 2016, 49, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Boonmars, T.; Boonjaraspinyo, S.; Nagano, I.; Pinlaor, S.; Puapairoj, A.; Yongvanit, P.; Takahashi, Y. Candidate genes involving in tumorigenesis of cholangiocarcinoma induced by Opisthorchis viverrini infection. Parasitol. Res. 2011, 109, 657–673. [Google Scholar] [CrossRef]
- Sripa, B.; Brindley, P.J.; Mulvenna, J.; Laha, T.; Smout, M.J.; Mairiang, E.; Bethony, J.M.; Loukas, A. The tumorigenic liver fluke Opisthorchis viverrini--multiple pathways to cancer. Trends Parasitol. 2012, 28, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaeteewoottacharn, K.; Seubwai, W.; Bhudhisawasdi, V.; Okada, S.; Wongkham, S. Potential targeted therapy for liver fluke associated cholangiocarcinoma. J. Hepatobiliary Pancreat Sci. 2014, 21, 362–370. [Google Scholar] [CrossRef]
- Seubwai, W.; Wongkham, C.; Puapairoj, A.; Khuntikeo, N.; Pugkhem, A.; Hahnvajanawong, C.; Chaiyagool, J.; Umezawa, K.; Okada, S.; Wongkham, S. Aberrant expression of NF-kappaB in liver fluke associated cholangiocarcinoma: Implications for targeted therapy. PLoS ONE 2014, 9, e106056. [Google Scholar] [CrossRef]
- Dabney, R.S.; Khalife, M.; Shahid, K.; Phan, A.T. Molecular pathways and targeted therapy in cholangiocarcinoma. Clin. Adv. Hematol. Oncol. 2019, 17, 630–637. [Google Scholar] [PubMed]
- Massironi, S.; Pilla, L.; Elvevi, A.; Longarini, R.; Rossi, R.E.; Bidoli, P.; Invernizzi, P. New and Emerging Systemic Therapeutic Options for Advanced Cholangiocarcinoma. Cells 2020, 9, 688. [Google Scholar] [CrossRef] [Green Version]
- Zaghloul, M.S.; Zaghloul, T.M.; Bishr, M.K.; Baumann, B.C. Urinary schistosomiasis and the associated bladder cancer: Update. J. Egypt Natl. Canc. Inst. 2020, 32, 44. [Google Scholar] [CrossRef]
- Smith, J.H.; Christie, J.D. The pathobiology of Schistosoma haematobium infection in humans. Hum. Pathol. 1986, 17, 333–345. [Google Scholar] [CrossRef]
- Honeycutt, J.; Hammam, O.; Fu, C.L.; Hsieh, M.H. Controversies and challenges in research on urogenital schistosomiasis-associated bladder cancer. Trends Parasitol. 2014, 30, 324–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botelho, M.; Ferreira, A.C.; Oliveira, M.J.; Domingues, A.; Machado, J.C.; da Costa, J.M. Schistosoma haematobium total antigen induces increased proliferation, migration and invasion, and decreases apoptosis of normal epithelial cells. Int. J. Parasitol. 2009, 39, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Veiga, I.; Oliveira, P.A.; Lopes, C.; Teixeira, M.; da Costa, J.M.; Machado, J.C. Carcinogenic ability of Schistosoma haematobium possibly through oncogenic mutation of KRAS gene. Adv. Cancer Res. Treat. 2013, 2013, 876585. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. DPDx—Laboratory Identificaiton of Parasites of Public Health Concern: Schistosomiasis. Available online: https://www.cdc.gov/dpdx/schistosomiasis/index.html (accessed on 24 August 2021).
- Moch, H.; Ulbright, T.M.; Reuter, V.E. WHO Classification of Tumours of the Urinary System and Male Genital Organs, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2016. [Google Scholar]
- Molehin, A.J. Schistosomiasis vaccine development: Update on human clinical trials. J. Biomed. Sci. 2020, 27, 28. [Google Scholar] [CrossRef]
- Sundahl, N.; Rottey, S.; De Maeseneer, D.; Ost, P. Pembrolizumab for the treatment of bladder cancer. Expert Rev. Anticancer Ther. 2018, 18, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Center JHUoMCR. COVID-19 Dashboard. Available online: https://coronavirus.jhu.edu/map.html (accessed on 24 August 2021).
- Marini, J.J.; Gattinoni, L. Management of COVID-19 Respiratory Distress. JAMA 2020, 323, 2329–2330. [Google Scholar] [CrossRef]
- Sadhukhan, P.; Ugurlu, M.T.; Hoque, M.O. Effect of COVID-19 on Lungs: Focusing on Prospective Malignant Phenotypes. Cancers 2020, 12, 3822. [Google Scholar] [CrossRef]
- Sabbatino, F.; Conti, V.; Franci, G.; Sellitto, C.; Manzo, V.; Pagliano, P.; De Bellis, E.; Masullo, A.; Salzano, F.A.; Caputo, A.; et al. PD-L1 Dysregulation in COVID-19 Patients. Front. Immunol. 2021, 12, 695242. [Google Scholar] [CrossRef]
- Thanh Le, T.; Andreadakis, Z.; Kumar, A.; Gomez Roman, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chua, T.H.; Punjabi, L.S.; Khor, L.Y. Tissue Pathogens and Cancers: A Review of Commonly Seen Manifestations in Histo- and Cytopathology. Pathogens 2021, 10, 1410. https://doi.org/10.3390/pathogens10111410
Chua TH, Punjabi LS, Khor LY. Tissue Pathogens and Cancers: A Review of Commonly Seen Manifestations in Histo- and Cytopathology. Pathogens. 2021; 10(11):1410. https://doi.org/10.3390/pathogens10111410
Chicago/Turabian StyleChua, Tzy Harn, Lavisha S Punjabi, and Li Yan Khor. 2021. "Tissue Pathogens and Cancers: A Review of Commonly Seen Manifestations in Histo- and Cytopathology" Pathogens 10, no. 11: 1410. https://doi.org/10.3390/pathogens10111410
APA StyleChua, T. H., Punjabi, L. S., & Khor, L. Y. (2021). Tissue Pathogens and Cancers: A Review of Commonly Seen Manifestations in Histo- and Cytopathology. Pathogens, 10(11), 1410. https://doi.org/10.3390/pathogens10111410