
Metabolic Profile and Mycoherbicidal Activity of Three Alternaria alternata Isolates for the Control of Convolvulus arvensis, Sonchus oleraceus, and Xanthium strumarium




Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of Alternaria alternata Isolates C1, S1, and X3
2.2. Herbicidal Activity (Detached Leaf Assay)
2.2.1. Effect of C1 Metabolites on Detached Leaves of Convolvulus arvensis
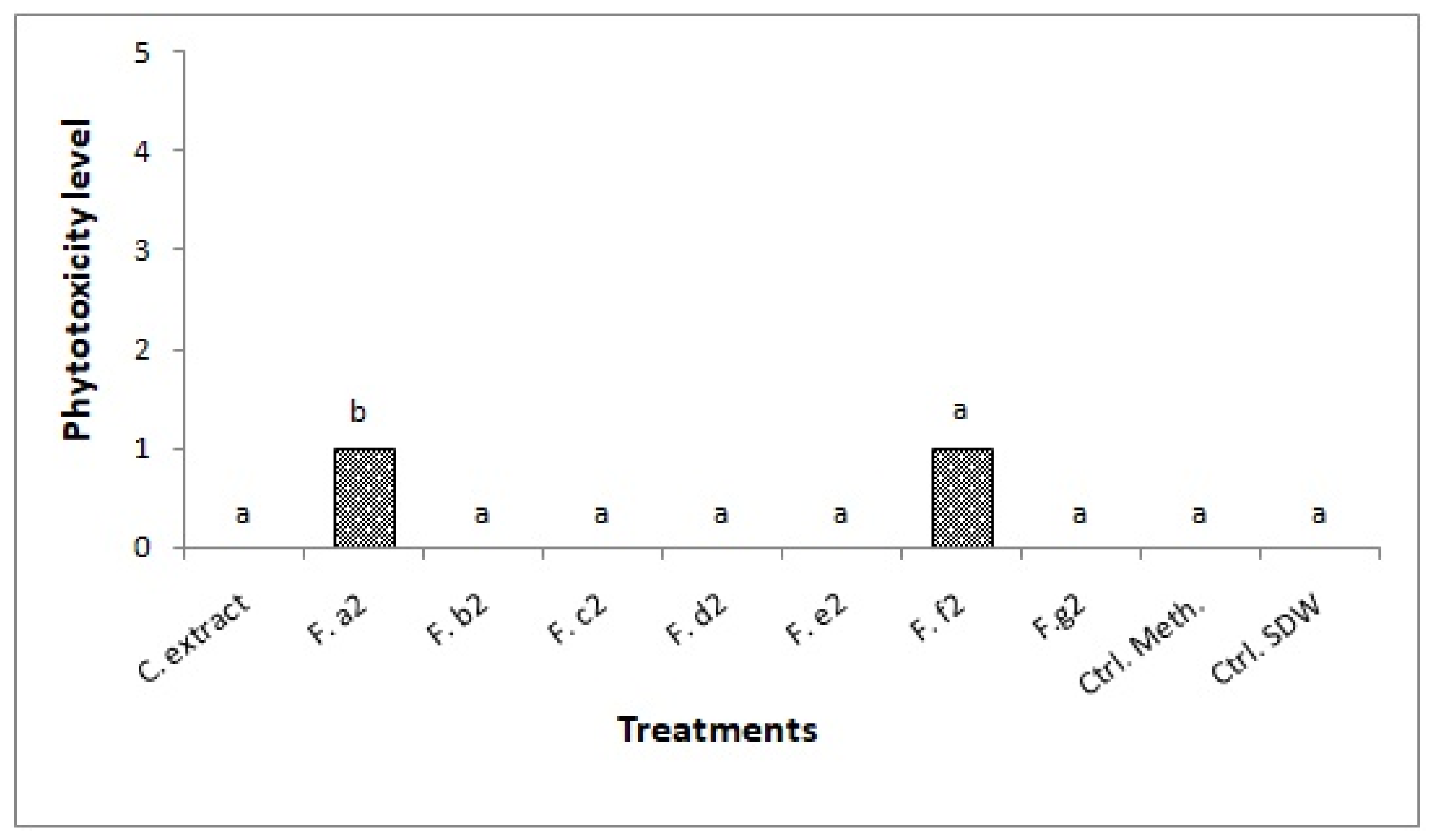
2.2.2. Effect of S1 Metabolites on Detached Leaves of Sonchus oleraceus
2.2.3. Effect of X3 Metabolites on Detached Leaves of Xanthium strumarium
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Metabolites Production and Extraction
4.3. Metabolite Fractionation
4.4. HPLC-MS Analysis
4.5. Detached Leaf Assay
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simmons, E.G. Alternaria: An Identification Manual, 2nd ed.; CBS Biodiversity: Utrecht, The Netherlands, 2007; p. 775. [Google Scholar]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria toxins: Potential virulence factors and genes related to pathogenesis. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Nielsen, K.F.; Pinto, V.F.; Patriarca, A. Characterization of Alternaria strains from Argentinean blueberry, tomato, walnut and wheat. Int. J. Food Microbiol. 2015, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Patriarcaa, A.; Cabrala, L.D.C.; Pavicicha, M.A.; Nielsen, K.F.; Andersen, B. Secondary metabolite profiles of small-spored Alternaria support the new phylogenetic organization of the genus. Int. J. Food Microbiol. 2019, 29, 135–143. [Google Scholar] [CrossRef]
- Meenaa, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation and their toxic effects. Toxicol. Rep. 2019, 6, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, I.; Ortega, S.F.; Gilardi, G.; Bosio, P.; Garibaldi, A.; Gullino, M.L. Molecular phylogeny and characterization of secondary metabolite profile of plant pathogenic Alternaria species isolated from basil. Food Microbiol. 2018, 73, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenfield, D.; Bunkers, G.; Strobel, G.A.; Sugawara, F. Potential new herbicides—Phytotoxins from plant pathogens. Weed Technol. 1988, 2, 519–524. [Google Scholar] [CrossRef]
- Evidente, A.; Cimmino, A.; Andolfi, A.; Berestetskiy, A.; Motta, A. Phomachalasins A-D, 26-oxa [16] and [15] cytochalasans produced by Phomaexigua var. exigua, a potential mycoherbicide for Cirsium arvense biocontrol. Tetrahedron 2011, 67, 1557–1563. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef]
- Charudattan, R.; Dinoor, A. Biological control of weeds using plant pathogens: Accomplishments and limitations. Crop Prot. 2000, 19, 691–695. [Google Scholar] [CrossRef]
- Vurro, M.; Evidente, A.; Andolfi, A.; Zonno, M.C.; Giordano, F.; Motta, A. Brefeldin A and a,b-dehydrocurvularin, two phytotoxins from Alternaria zinniae, a biocontrol agent of Xanthium occidentale. Plant Sci. 1998, 138, 67–79. [Google Scholar] [CrossRef]
- Sanodiya, B.S.; Thakur, G.S.; Baghel, R.K.; Pandey, A.K.; Prasad, G.B.K.S.; Bisen, P.S. Isolation and characterization of tenuazonic acid produced by Alternaria alternata, a potential bioherbicidal agent for control of Lantana camara. J. Plant Prot. Res. 2010, 50, 133–139. [Google Scholar] [CrossRef]
- Davis, N.D.; Diener, U.L.; Morgan-Jones, G. Tenuazonic Acid Production by Alternaria alternata and Alternaria tenuissima Isolated from Cotton, Appl. Environ. Microbiol. 1977, 34, 155–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanodiya, B.S.; Thakur, G.S.; Debnath, M.; Pandey, A.K.; Prasadd, G.B.K.S.; Bisen, P.S. Characterization of herbicidal potential of tenuazonic acid isolated from Alternaria alternata for the management of Parthenium hysterophorus. Rom. J. Plant Prot. 2009, 2, 49–61. [Google Scholar]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cottrill, B.; Cravedi, J.; Farmer, P. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407–2504. [Google Scholar]
- Yun, C.S.; Motoyama, T.; Osada, H. Biosynthesis of the mycotoxin tenuazonic acid by a fungal NRPS–PKS hybrid enzyme. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, F.A.; Rightsel, W.A.; Sloan, B.J.; Ehrlich, J.; French, J.C.; Bartz, Q.R.; Dixon, G.J. Antiviral activity of tenuazonic acid. Nature 1963, 200, 1338–1339. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, M.H.; Nicolas, L.; Boutar, M.; Gaudemer, F.; Ranomenjanahary, S.; Gaudemer, A. Relationships between the structure and the phytotoxicity of the fungal toxin tenuazonic acid. Phytochemistry 1988, 27, 77–84. [Google Scholar] [CrossRef]
- Chen, S.; Qiang, S. Recent advances in tenuazonic acid as a potential herbicide. Pestic. Biochem. Physiol. 2017, 143, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Wang, H.; Meng, B.; Wei, R.; Wang, L.; An, C.; Chen, S.; Yang, C.; Qiang, S. An evaluation of tenuazonic acid, a potential biobased herbicide in cotton. Pest Manag. Sci. 2019, 75, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Abdessemed, N.; Kerroum, A.; Bahet, Y.A.; Talbi, N.; Zermane, N. First report of Alternaria leaf spot caused by Alternaria alternata (Fries.) Kiessler on Sonchusoleraceus L. and Convolvulus arvensis L. in Algeria. J. Phytopathol. 2019, 167, 321–325. [Google Scholar] [CrossRef]
- Abdessemed, N.; Bahet, Y.A.; Zermane, N. Mycoherbicide potential of Alternaria alternata (Fries.) Kiessler and its formulations on the host weed Xanthium strumarium L. Biocontrol Sci. Technol. 2020, 30, 1300–1315. [Google Scholar] [CrossRef]
- Abdessemed, N. Champignons À Potentiel Mycoherbicide. Ph.D. Thesis, Ecole Nationale Supérieure Agronomique, El-Harrach, Algeria, 7 January 2021. [Google Scholar]
- Sottor, I.; Wiese, J.; Lipfert, M.; Preußke, N.; Sönnichsen, F.D.; Imho, J.F. Different Secondary Metabolite Profiles of Phylogenetically almost Identical Streptomyces griseus Strains Originating from Geographically Remote Locations. Microorganisms 2019, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Uka, V.; Moore, G.G.; Arroyo-Manzanares, N.; Dashnor, N.; De Saeger, S.; Di Mavungu, J.D. Secondary Metabolite Dereplication and Phylogenetic Analysis Identify Various Emerging Mycotoxins and Reveal the High Intra-Species Diversity in Aspergillus flavus. Front. Microbiol. 2019, 10, 667. [Google Scholar] [CrossRef] [Green Version]
- Llorens, A.; Mateo, R.; Hinojo, M.J.; Valle-Algarra, F.M.; Jiménez, M. Influence of environmental factors on the biosynthesis of type B trichothecenes by isolates of Fusarium spp. from Spanish crops. Int. J. Food Microbiol. 2004, 94, 43–54. [Google Scholar] [CrossRef]
- Schumacher, M.J.; Sandra, E.J. Variability of Alternaria alternata: Biochemical and immunological characteristics of culture filtrates from seven isolates. J. Allergy Clin. Immunol. 1976, 58, 263–277. [Google Scholar] [CrossRef]
- Khaldi, N.; Collemare, J.; Lebrun, M.H.; Wolfe, K.H. Evidence for horizontal transfer of a secondary metabolite gene cluster between fungi. Genome Boil. 2008, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ziemert, N.; Lechner, A.; Wietz, M.; Millán-Aguiñaga, N.; Chavarria, K.L.; Jensen, P.R. Diversity and evolution of secondary metabolism in the marine actinomycete genus Salinispora. Proc. Natl. Acad. Sci. USA 2014, 111, E1130–E1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meronuck, R.A.; Steele, J.A.; Mirocha, C.J.; Christensen, C.M. Tenuazonic Acid, a Toxin Produced by Alternaria alternata. Appl. Microbiol. 1972, 23, 613–617. [Google Scholar] [CrossRef]
- Rosett, T.; Sankhala, R.H.; Stickings, C.E.; Taylor, M.E.U.; Thomas, R. Studies in the biochemistry of micro-organisms. 103. Metabolites of Alternaria tenuis Auct.: Culture filtrate products. Biochem. J. 1957, 67, 390–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Shibata, K.; Nobe, K.; Hasumi, K.; Honda, K. A Novel Embolic Model of Cerebral Infarction and Evaluation of Stachybotrys microspora Triprenyl Phenol-7 (SMTP-7), a Novel Fungal Triprenyl Phenol Metabolite. J. Pharmacol. Sci. 2010, 114, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Sawada, H.; Nishimura, N.; Suzuki, E.; Zhuang, J.; Hasegawa, K.; Takamatsu, H.; Honda, K.; Hasumi, K. SMTP-7, a novel small-molecule thrombolytic for ischemic stroke: A study in rodents and primates. J. Cereb. Blood Flow Metab. 2014, 34, 235–241. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Cutignano, A.; Taglialatela-Scafati, O.; Vurro, M.; Zonno, M.C.; Motta, A. Ascaulitoxin, a phytotoxique bis-amino acid N-glucoside from Ascochytacaulina. Phytochemistry 1998, 48, 1131–1137. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Berestetskiy, A.; Vurro, M.; Evidente, A. Chenopodolans A–C: Phytotoxic furopyrans produced by Phoma chenopodiicola, a fungal pathogen of Chenopodium album. Phytochemistry 2013, 96, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Zehra, A.; Dubey, M.K.; Aamir, M.; Gupta, V.K.; Upadhyay, R.S. Comparative evaluation of biochemical changes in tomato (Lycopersicon esculentum Mill.) infected by Alternaria alternata and its toxic metabolites (TeA, AOH, and AME). Front. Plant Sci. 2016, 7, 1408. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Swapnil, P.; Upadhyay, R.S. Isolation, characterization and toxicological potential of Alternaria-mycotoxins (TeA, AOH and AME) in different Alternaria species from various regions of India. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Abdessemed, N.; Kerroum, A.; Bahet, Y.A.; Zermane, N. First report of Alternaria alternata leaf spot on Xanthium strumarium L. in Algeria. Plant Pathol. J. 2019, 101, 781. [Google Scholar] [CrossRef] [Green Version]
- Qiang, S.; Wang, L.; Wei, R.; Zhou, B.; Chen, S.; Zhu, Y.; Dong, Y.; An, C. Bioassay of the Herbicidal Activity of AAC-Toxin Produced by Alternaría alternata isolated from Ageratina adenophora. Weed Technol. 2010, 24, 197–201. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Alternaria alternata Isolates | ||
|---|---|---|---|
| C1 | S1 | X3 | |
| Cyclo-l-prolylglycine | + | + | + |
| 12-Methoxycitromycin | + | − | − |
| Erythroglaucin | + | − | − |
| Alternarian acid | + | − | − |
| TAN 913 | + | − | − |
| Verrol | + | + | + |
| Tenuazonicacid | + | + | + |
| Brassicicene F | + | + | + |
| Rezishanone C | + | + | + |
| Brassicicene D | + | + | − |
| Chaetoquadrin E | + | − | − |
| Tanzawaicacid F | + | − | − |
| Cytosporin C | − | + | − |
| Striatisporin | − | + | − |
| Cyclo-(Pro-Ala) | − | − | + |
| Triprenyl phenol-7 (SMTP-7) | + | + | + |
| Alternaria alternataIsolates | Active Fractions | Compounds |
|---|---|---|
| A. alternata C1 | d1 | SMTP-7 (+)-Phomopsidin Tenuazonic acid |
| e1 | Decarestrictin N SMTP-7 Tenuazonic acid | |
| h1 | SMTP-7 Tenuazonic acid | |
| i1 | Decarestrictin N SMTP-7 Tenuazonic acid | |
| A. alternata S1 | a2 | SMTP-7 |
| f2 | Decarestrictin N SMTP-7 Tenuazonic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdessemed, N.; Staropoli, A.; Zermane, N.; Vinale, F. Metabolic Profile and Mycoherbicidal Activity of Three Alternaria alternata Isolates for the Control of Convolvulus arvensis, Sonchus oleraceus, and Xanthium strumarium. Pathogens 2021, 10, 1448. https://doi.org/10.3390/pathogens10111448
Abdessemed N, Staropoli A, Zermane N, Vinale F. Metabolic Profile and Mycoherbicidal Activity of Three Alternaria alternata Isolates for the Control of Convolvulus arvensis, Sonchus oleraceus, and Xanthium strumarium. Pathogens. 2021; 10(11):1448. https://doi.org/10.3390/pathogens10111448
Chicago/Turabian StyleAbdessemed, Nesma, Alessia Staropoli, Nadjia Zermane, and Francesco Vinale. 2021. "Metabolic Profile and Mycoherbicidal Activity of Three Alternaria alternata Isolates for the Control of Convolvulus arvensis, Sonchus oleraceus, and Xanthium strumarium" Pathogens 10, no. 11: 1448. https://doi.org/10.3390/pathogens10111448
APA StyleAbdessemed, N., Staropoli, A., Zermane, N., & Vinale, F. (2021). Metabolic Profile and Mycoherbicidal Activity of Three Alternaria alternata Isolates for the Control of Convolvulus arvensis, Sonchus oleraceus, and Xanthium strumarium. Pathogens, 10(11), 1448. https://doi.org/10.3390/pathogens10111448